
Multiple Myeloma Cell Simulation Using an Agent-Based Framework Coupled with a Continuous Fluid Model

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Methods
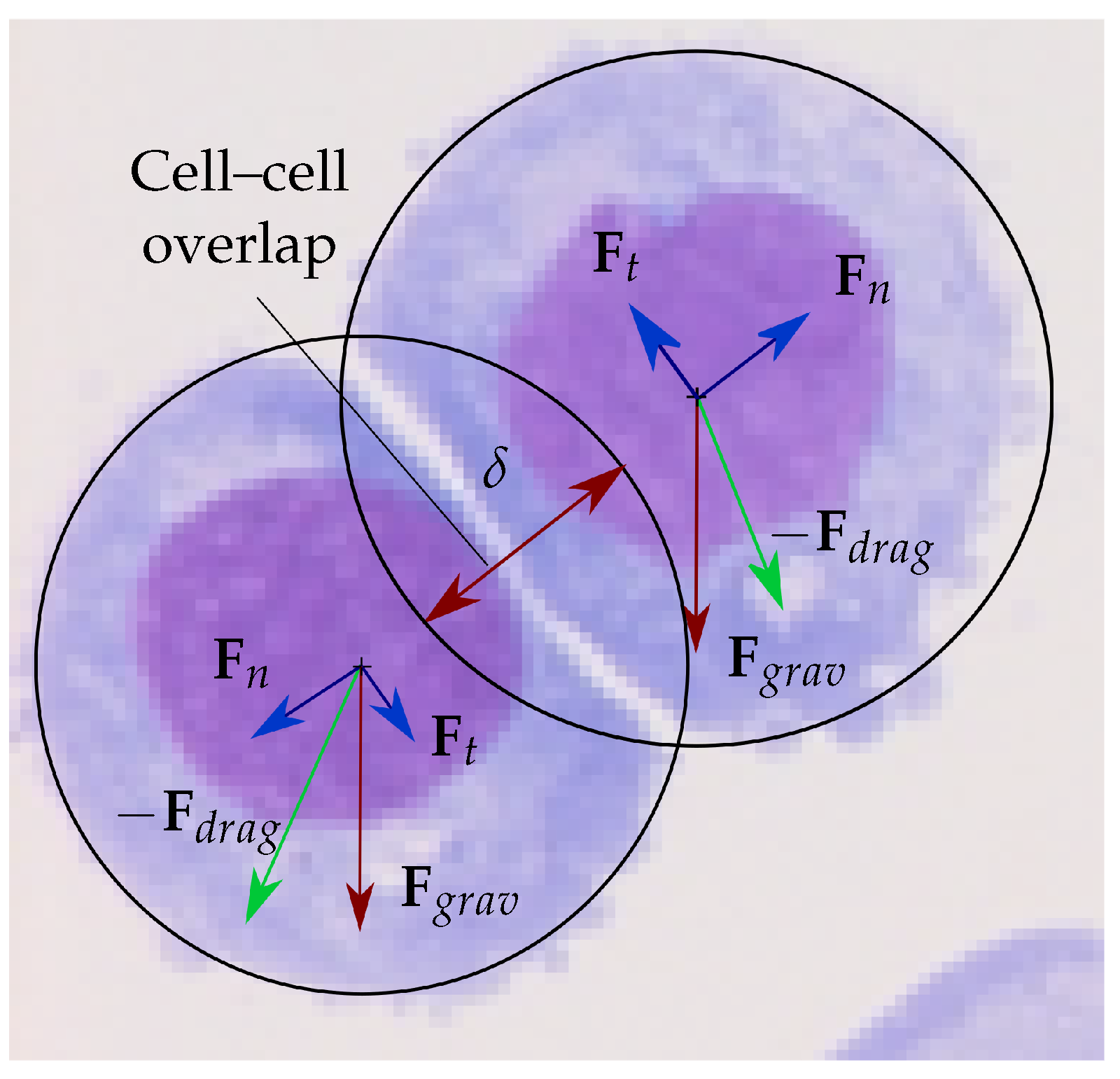
2.1. Multiphase Coupled Fluid-Particle Model (ANS)
2.2. Design of In Vitro Experiments and Data Collection
2.3. Model Assumptions
3. Results
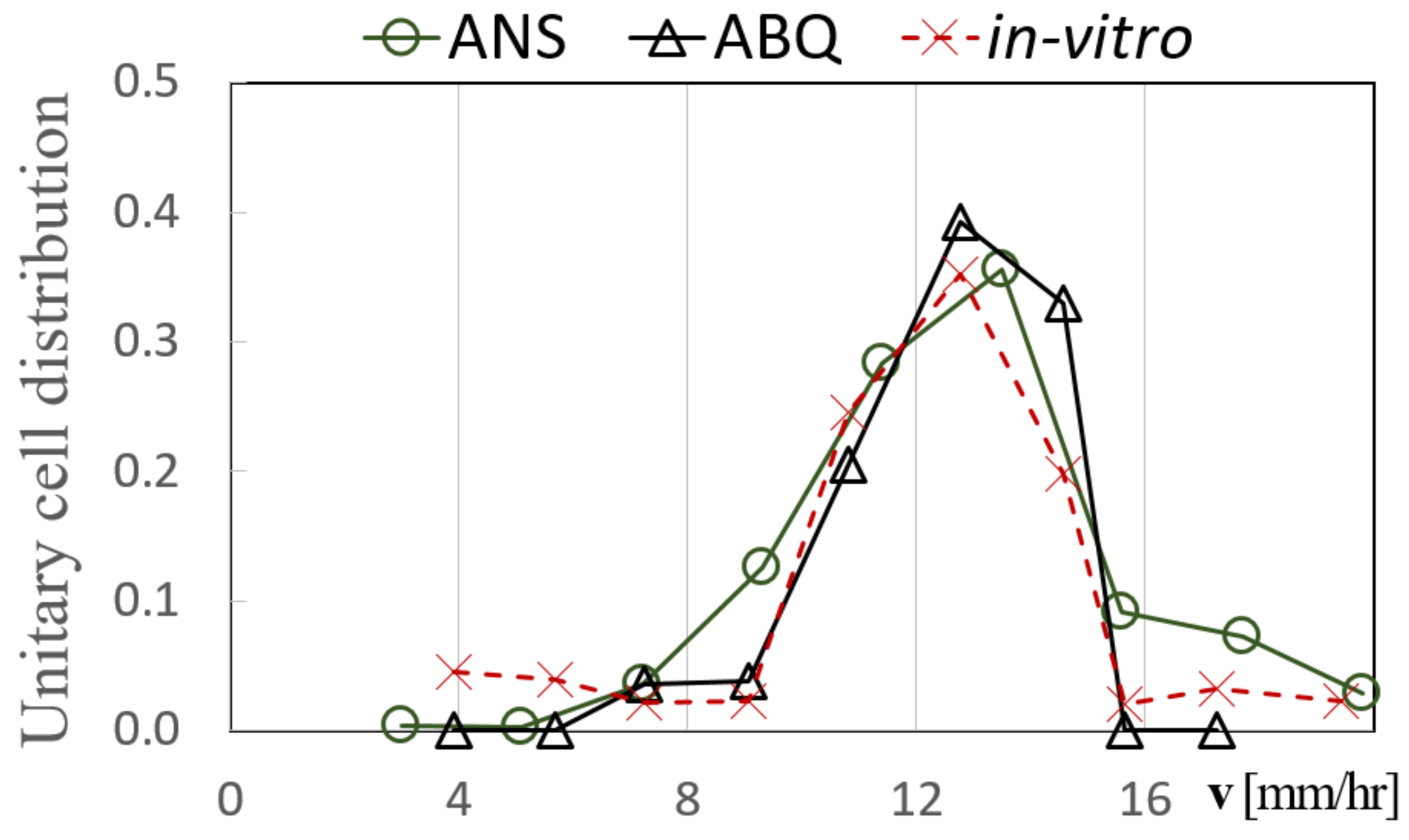
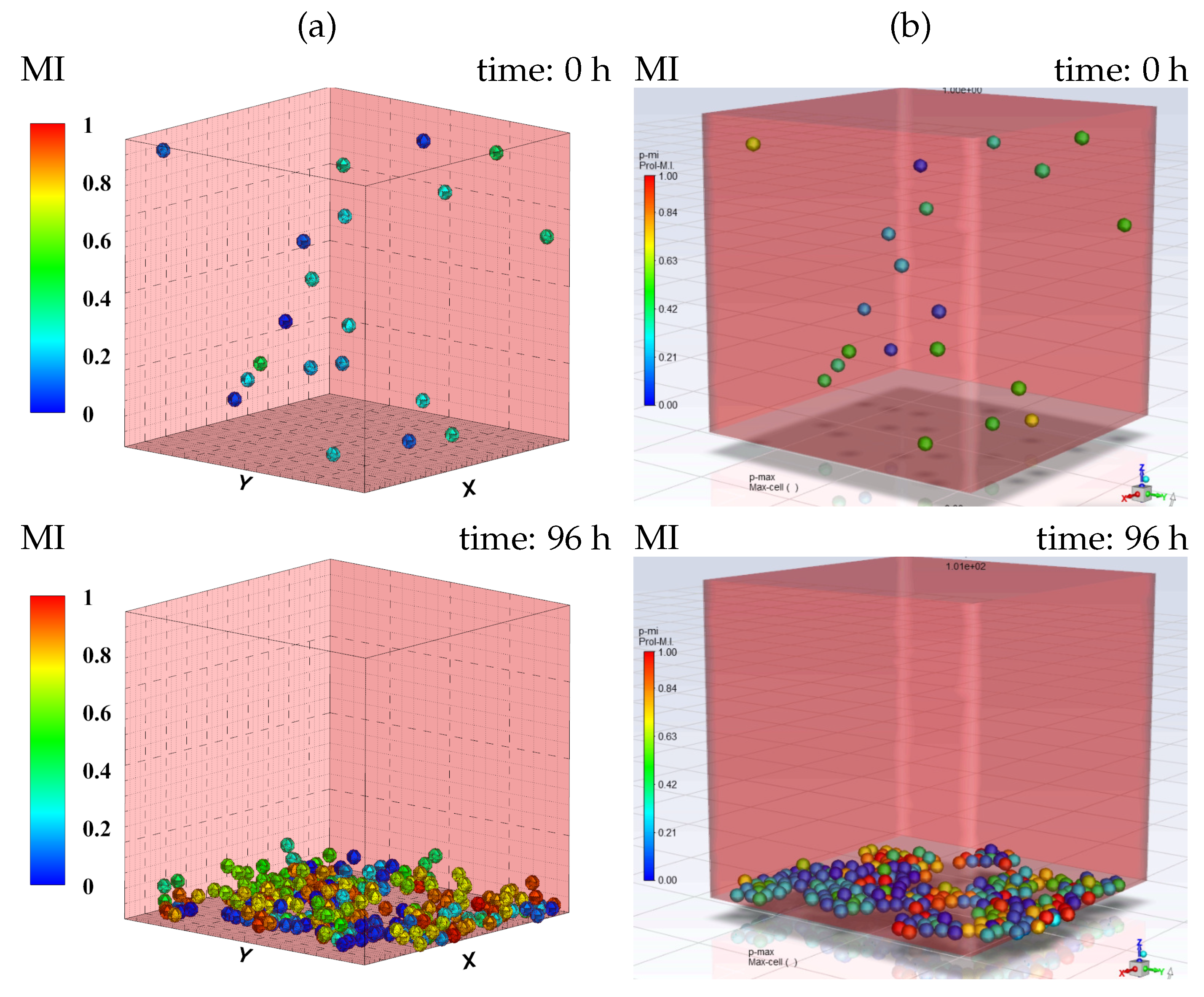
3.1. Cell Sedimentation
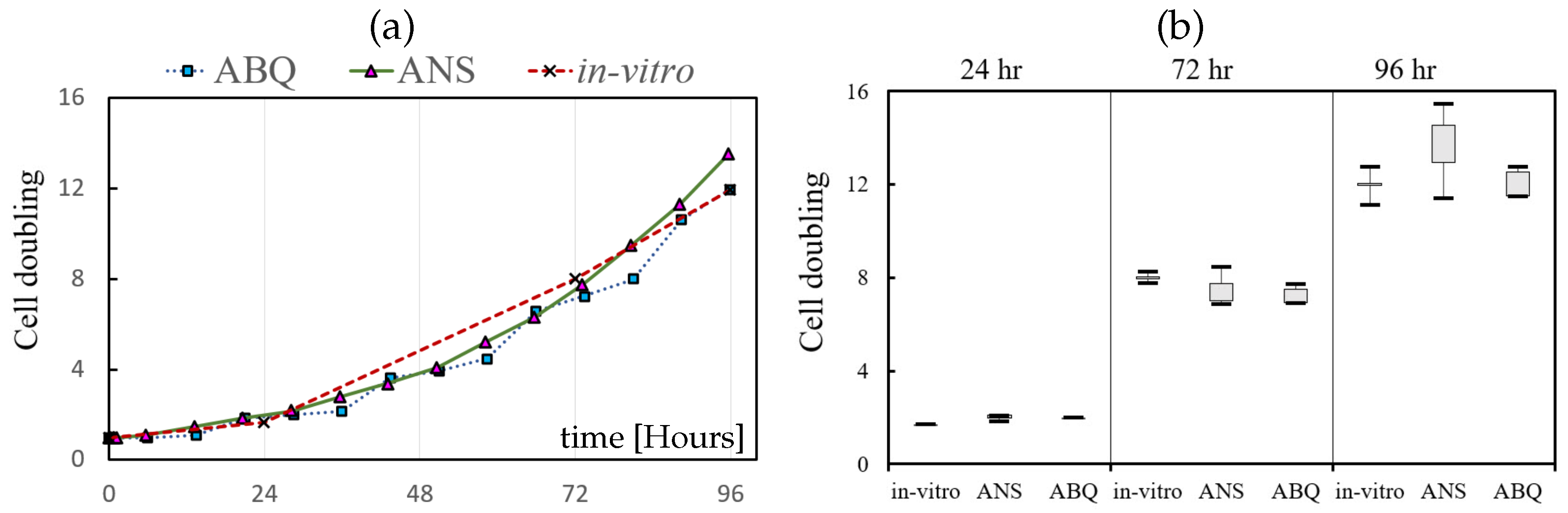
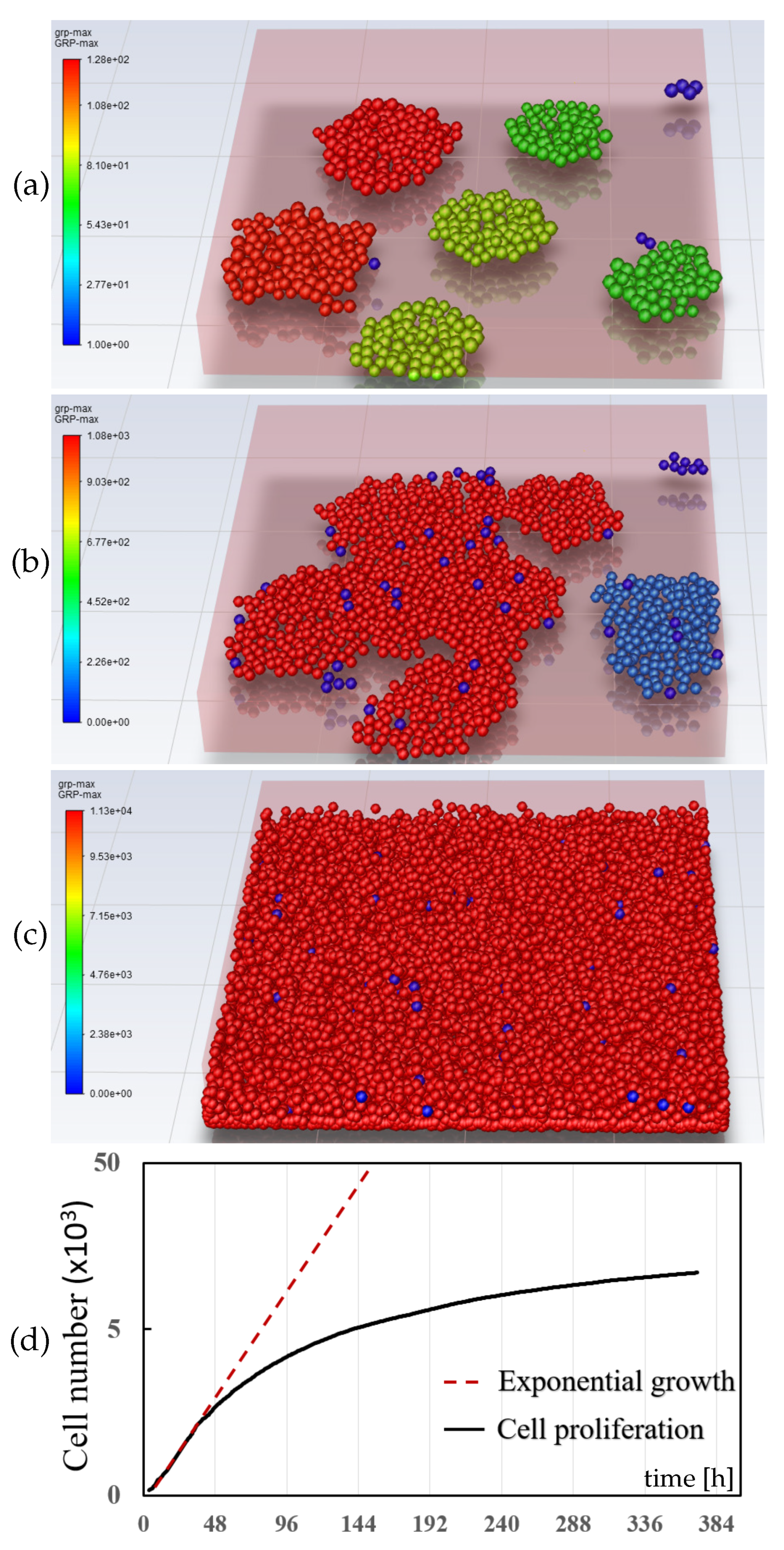
3.2. Cell Proliferation
3.3. Tumor Aggregation Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, S.K.; Rajkumar, V.; Kyle, R.A.; van Duin, M.; Sonneveld, P.; Mateos, M.V.; Gay, F.; Anderson, K.C. Multiple myeloma. Nat. Rev. Dis. Prim. 2017, 3, 17046. [Google Scholar] [CrossRef] [PubMed]
- Perez-Amill, L.; Suñe, G.; Antoñana-Vildosola, A.; Castella, M.; Najjar, A.; Bonet, J.; Fernández-Fuentes, N.; Inogés, S.; López, A.; Bueno, C.; et al. Preclinical development of a humanized chimeric antigen receptor against B cell maturation antigen for multiple myeloma. Haematologica 2020, 106, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Muz, B.; Alhallak, K.; Markovic, M.; Gurley, S.; Wang, Z.; Guenthner, N.; Wasden, K.; Fiala, M.; King, J.; et al. Targeting CD47 as a Novel Immunotherapy for Multiple Myeloma. Cancers 2020, 12, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, Y.W.; Walsh, K.; Yao, L.; Kedei, N.; Blumberg, P.M.; Rubin, J.S.; Shaughnessy, J.; Rudikoff, S. Wnts induce migration and invasion of myeloma plasma cells. Blood 2005, 106, 1786–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Guo, X.; Su, J.; Chen, R.; Berenzon, D.; Guthold, M.; Bonin, K.; Zhao, W.; Zhou, X. CD138-negative myeloma cells regulate mechanical properties of bone marrow stromal cells through SDF-1/CXCR4/AKT signaling pathway. Biochim. Biophys. Acta-Mol. Cell Res. 2015, 1853, 338–347. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Ofek, G.; Choi, D.S.; Wen, J.; Hu, J.; Zhao, H.; Zu, Y.; Athanasiou, K.A.; Chang, C.C. Unique biomechanical interactions between myeloma cells and bone marrow stroma cells. Prog. Biophys. Mol. Biol. 2010, 103, 148–156. [Google Scholar] [CrossRef]
- Clara-Trujillo, S.; Ferrer, G.G.; Ribelles, J.L.G. In Vitro Modeling of Non-Solid Tumors: How Far Can Tissue Engineering Go? Int. J. Mol. Sci. 2020, 21, 5747. [Google Scholar] [CrossRef]
- Podar, K.; Tai, Y.T.; Lin, B.K.; Narsimhan, R.P.; Sattler, M.; Kijima, T.; Salgia, R.; Gupta, D.; Chauhan, D.; Anderson, K.C. Vascular Endothelial Growth Factor-induced Migration of Multiple Myeloma Cells Is Associated with β1 Integrin- and Phosphatidylinositol 3-Kinase-dependent PKCα Activation. J. Biol. Chem. 2002, 277, 7875–7881. [Google Scholar] [CrossRef] [Green Version]
- Zlei, M.; Egert, S.; Wider, D.; Ihorst, G.; Wäsch, R.; Engelhardt, M. Characterization of in vitro growth of multiple myeloma cells. Exp. Hematol. 2007, 35, 1550–1561. [Google Scholar] [CrossRef]
- Jin, J.; Wang, T.; Wang, Y.; Chen, S.; Li, Z.; Li, X.; Zhang, J.; Wang, J. SRC3 expressed in BMSCs promotes growth and migration of multiple myeloma cells by regulating the expression of Cx43. Int. J. Oncol. 2017, 51, 1694–1704. [Google Scholar] [CrossRef] [Green Version]
- Huh, D.; Hamilton, G.A.; Ingber, D.E. From 3D cell culture to organs-on-chips. Trends Cell Biol. 2011, 21, 745–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.N.; Ingber, D.E. Microfluidic organs-on-chips. Nat. Biotechnol. 2014, 32, 760–772. [Google Scholar] [CrossRef]
- Mogilner, A. Mathematics of cell motility: Have we got its number? J. Math. Biol. 2009, 58, 105–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, M.L.; McGarry, P.J.; Sniadecki, N.J. Review on cell mechanics: Experimental and modeling approaches. Appl. Mech. Rev. 2013, 65, 060801. [Google Scholar] [CrossRef] [Green Version]
- Ayensa-Jiménez, J.; Pérez-Aliacar, M.; Randelovic, T.; Oliván, S.; Fernández, L.; Sanz-Herrera, J.A.; Ochoa, I.; Doweidar, M.H.; Doblaré, M. Mathematical formulation and parametric analysis of in vitro cell models in microfluidic devices: Application to different stages of glioblastoma evolution. Sci. Rep. 2020, 10, 21193. [Google Scholar] [CrossRef]
- Cioffi, M.; Küffer, J.; Ströbel, S.; Dubini, G.; Martin, I.; Wendt, D. Computational evaluation of oxygen and shear stress distributions in 3D perfusion culture systems: Macro-scale and micro-structured models. J. Biomech. 2008, 41, 2918–2925. [Google Scholar] [CrossRef]
- Soleimani, S.; Shamsi, M.; Ghazani, M.A.; Modarres, H.P.; Valente, K.P.; Saghafian, M.; Ashani, M.M.; Akbari, M.; Sanati-Nezhad, A. Translational models of tumor angiogenesis: A nexus of in silico and in vitro models. Biotechnol. Adv. 2018, 36, 880–893. [Google Scholar] [CrossRef]
- Carlier, A.; Skvortsov, G.A.; Hafezi, F.; Ferraris, E.; Patterson, J.; Koc, B.; Van Oosterwyck, H. Computational model-informed design and bioprinting of cell-patterned constructs for bone tissue engineering. Biofabrication 2016, 8, 025009. [Google Scholar] [CrossRef]
- Kang, K.T.; Park, J.H.; Kim, H.J.; Lee, H.Y.H.M.; Lee, K.I.; Jung, H.H.; Lee, H.Y.H.M.; Jang, J.W. Study of Tissue Differentiation of Mesenchymal Stem Cells by Mechanical Stimuli and an Algorithm for Bone Fracture Healing. Tissue Eng. Regen. Med. 2011, 8, 359–370. [Google Scholar]
- Fouliard, S.; Benhamida, S.; Lenuzza, N.; Xavier, F. Modeling and simulation of cell populations interaction. Math. Comput. Model. 2009, 49, 2104–2108. [Google Scholar] [CrossRef]
- Bissell, M.J.; Rizki, A.; Mian, I.S. Tissue architecture: The ultimate regulator of breast epithelial function. Curr. Opin. Cell Biol. 2003, 15, 753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- te Boekhorst, V.; Preziosi, L.; Friedl, P. Plasticity of Cell Migration In Vivo and In Silico. Annu. Rev. Cell Dev. Biol. 2016, 32, 491–526. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.C.; Silberberg, Y.R.; Abeyaratne, R.; Kamm, R.D.; Asada, H.H. Computational modeling of three-dimensional ECM-rigidity sensing to guide directed cell migration. Proc. Natl. Acad. Sci. USA 2018, 115, E390–E399. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.J.; Doweidar, M.H. Encapsulated piezoelectric nanoparticle–hydrogel smart material to remotely regulate cell differentiation and proliferation: A finite element model. Comput. Mech. 2019, 63, 471–489. [Google Scholar] [CrossRef]
- Farsad, M.; Vernerey, F.J. An XFEM-based numerical strategy to model mechanical interactions between biological cells and a deformable substrate. Int. J. Numer. Methods Eng. 2012, 92, 238–267. [Google Scholar] [CrossRef]
- Katti, D.R.; Katti, K.S. Cancer cell mechanics with altered cytoskeletal behavior and substrate effects: A 3D finite element modeling study. J. Mech. Behav. Biomed. Mater. 2017, 76, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Urdeitx, P.; Farzaneh, S.; Mousavi, S.J.; Doweidar, M.H. Role of oxygen concentration in the osteoblasts behavior: A finite element model. J. Mech. Med. Biol. 2020, 20, 1950064. [Google Scholar] [CrossRef]
- Malekian, N.; Habibi, J.; Zangooei, M.H.; Aghakhani, H. Integrating evolutionary game theory into an agent-based model of ductal carcinoma in situ: Role of gap junctions in cancer progression. Comput. Methods Programs Biomed. 2016, 136, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Urdeitx, P.; Doweidar, M.H. Mechanical stimulation of cell microenvironment for cardiac muscle tissue regeneration: A 3D in-silico model. Comput. Mech. 2020, 66, 1003–1023. [Google Scholar] [CrossRef]
- Urdeitx, P.; Doweidar, M.H. A Computational Model for Cardiomyocytes Mechano-Electric Stimulation to Enhance Cardiac Tissue Regeneration. Mathematics 2020, 8, 1875. [Google Scholar] [CrossRef]
- ANSYS Inc. Fluent Theory Guide, 15th ed.; ANSYS, Inc.: Canonsburg, PA, USA, 2013; pp. 724–746. [Google Scholar]
- Abaqus (ABQ). Abaqus 6.14; Dassault Systemes: Vélizy-Villacoublay, France, 2014. [Google Scholar]
- Urdeitx, P.; Doweidar, M.H. Enhanced Piezoelectric Fibered Extracellular Matrix to Promote Cardiomyocyte Maturation and Tissue Formation: A 3D Computational Model. Biology 2021, 10, 135. [Google Scholar] [CrossRef] [PubMed]
- Urdeitx, P.; Mousavi, S.J.; Avril, S.; Doweidar, M.H. Computational modeling of multiple myeloma interactions with resident bone marrow cells. Comput. Biol. Med. 2023, 153, 106458. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.Q.; Chen, Q. Mechanoregulation of chondrocyte proliferation, maturation, and hypertrophy: Ion-channel dependent transduction of matrix deformation signals. Exp. Cell Res. 2000, 256, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Tse, J.; Jain, R.K.; Munn, L.L. Micro-environmental mechanical stress controls tumor spheroid size and morphology by suppressing proliferation and inducing apoptosis in cancer cells. PLoS ONE 2009, 4, e4632. [Google Scholar] [CrossRef] [Green Version]
- Isobe, T.; Ikeda, Y.; Ohta, H. Comparison of sizes and shapes of tumor cells in plasma cell leukemia and plasma cell myeloma. Blood 1979, 53, 1028–1030. [Google Scholar] [CrossRef] [Green Version]
- Clara-Trujillo, S.; Tolosa, L.; Cordón, L.; Sempere, A.; Ferrer, G.G.; Luis, J.; Ribelles, G.; Ribelles, J.L.G. Novel microgel culture system as semi-solid three-dimensional in vitro model for the study of multiple myeloma proliferation and drug resistance. Biomater. Adv. 2022, 135, 212749. [Google Scholar] [CrossRef]
- Bam, R.; Ling, W.; Khan, S.; Pennisi, A.; Venkateshaiah, S.U.; Li, X.; van Rhee, F.; Usmani, S.; Barlogie, B.; Shaughnessy, J.; et al. Role of Bruton’s tyrosine kinase in myeloma cell migration and induction of bone disease. Am. J. Hematol. 2013, 88, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Zipursky, A.; Bow, E.; Seshadri, R.; Brown, E. Leukocyte density and volume in normal subjects and in patients with acute lymphoblastic leukemia. Blood 1976, 48, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Aranda-Lara, L.; Torres-García, E.; Oros-Pantoja, R. Biological Tissue Modeling with Agar Gel Phantom for Radiation Dosimetry of 99mTc. Open J. Radiol. 2014, 4, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Thompson, B.R.; Horozov, T.S.; Stoyanov, S.D.; Paunov, V.N. An ultra melt-resistant hydrogel from food grade carbohydrates. RSC Adv. 2017, 7, 45535–45544. [Google Scholar] [CrossRef] [Green Version]
- WATASE, M.; ARAKAWA, K. Rheological Properties of Hydrogels of Agar-agar. Nippon Kagaku Zassi 1971, 92, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Hamburger, A.; Salmon, S.E. Primary Bioassay of Human Myeloma Stem Cells. J. Clin. Investig. 1977, 60, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Cowley, G.S.; Weir, B.A.; Vazquez, F.; Tamayo, P.; Scott, J.A.; Rusin, S.; East-Seletsky, A.; Ali, L.D.; Gerath, W.F.; Pantel, S.E.; et al. Parallel genome-scale loss of function screens in 216 cancer cell lines for the identification of context-specific genetic dependencies. Sci. Data 2014, 1, 140035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, K.E.; Huang, H.; Mythreye, K.; Blobe, G.C. The type III transforming growth factor-β receptor inhibits proliferation, migration, and adhesion in human myeloma cells. Mol. Biol. Cell 2011, 22, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Peacock, C.D.; Wang, Q.; Gesell, G.S.; Corcoran-Schwartz, I.M.; Jones, E.; Kim, J.; Devereux, W.L.; Rhodes, J.T.; Huff, C.A.; Beachy, P.A.; et al. Hedgehog signaling maintains a tumor stem cell compartment in multiple myeloma. Proc. Natl. Acad. Sci. USA 2007, 104, 4048–4053. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.J.; Doblaré, M.; Doweidar, M.H. Computational modelling of multi-cell migration in a multi-signalling substrate. Phys. Biol. 2014, 11, 026002. [Google Scholar] [CrossRef]
- Mousavi, S.J.; Doweidar, M.H. Numerical modeling of cell differentiation and proliferation in force-induced substrates via encapsulated magnetic nanoparticles. Comput. Methods Programs Biomed. 2016, 130, 106–117. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Value | Refs. |
|---|---|---|---|
| Nominal cell radius | m | [37] | |
| Cell mass density | 1077 kg/m3 | [39,40] | |
| ECM mass density | 940 kg/m3 | [41] | |
| ECM viscosity | Pa s | [42,43] | |
| E | Cell stiffness | kPa | [5,6] |
| ECM stiffness | kPa | [42] | |
| Friction coefficient for cell contacts | [6,44] | ||
| Natural maturation time without stimulus | 60 h | [45] | |
| Proportional time factor for cell–wall contacts | 1.60 | – | |
| Proportional time factor for cell–cell contacts | 0.14 | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urdeitx, P.; Clara-Trujillo, S.; Gomez Ribelles, J.L.; Doweidar, M.H. Multiple Myeloma Cell Simulation Using an Agent-Based Framework Coupled with a Continuous Fluid Model. Mathematics 2023, 11, 1824. https://doi.org/10.3390/math11081824
Urdeitx P, Clara-Trujillo S, Gomez Ribelles JL, Doweidar MH. Multiple Myeloma Cell Simulation Using an Agent-Based Framework Coupled with a Continuous Fluid Model. Mathematics. 2023; 11(8):1824. https://doi.org/10.3390/math11081824
Chicago/Turabian StyleUrdeitx, Pau, Sandra Clara-Trujillo, Jose Luis Gomez Ribelles, and Mohamed H. Doweidar. 2023. "Multiple Myeloma Cell Simulation Using an Agent-Based Framework Coupled with a Continuous Fluid Model" Mathematics 11, no. 8: 1824. https://doi.org/10.3390/math11081824