Molecular Cloning of Novel-Type Phosphoenolpyruvate Carboxylase Isoforms in Pitaya (Hylocereus undatus)

Abstract
:1. Introduction
2. Results
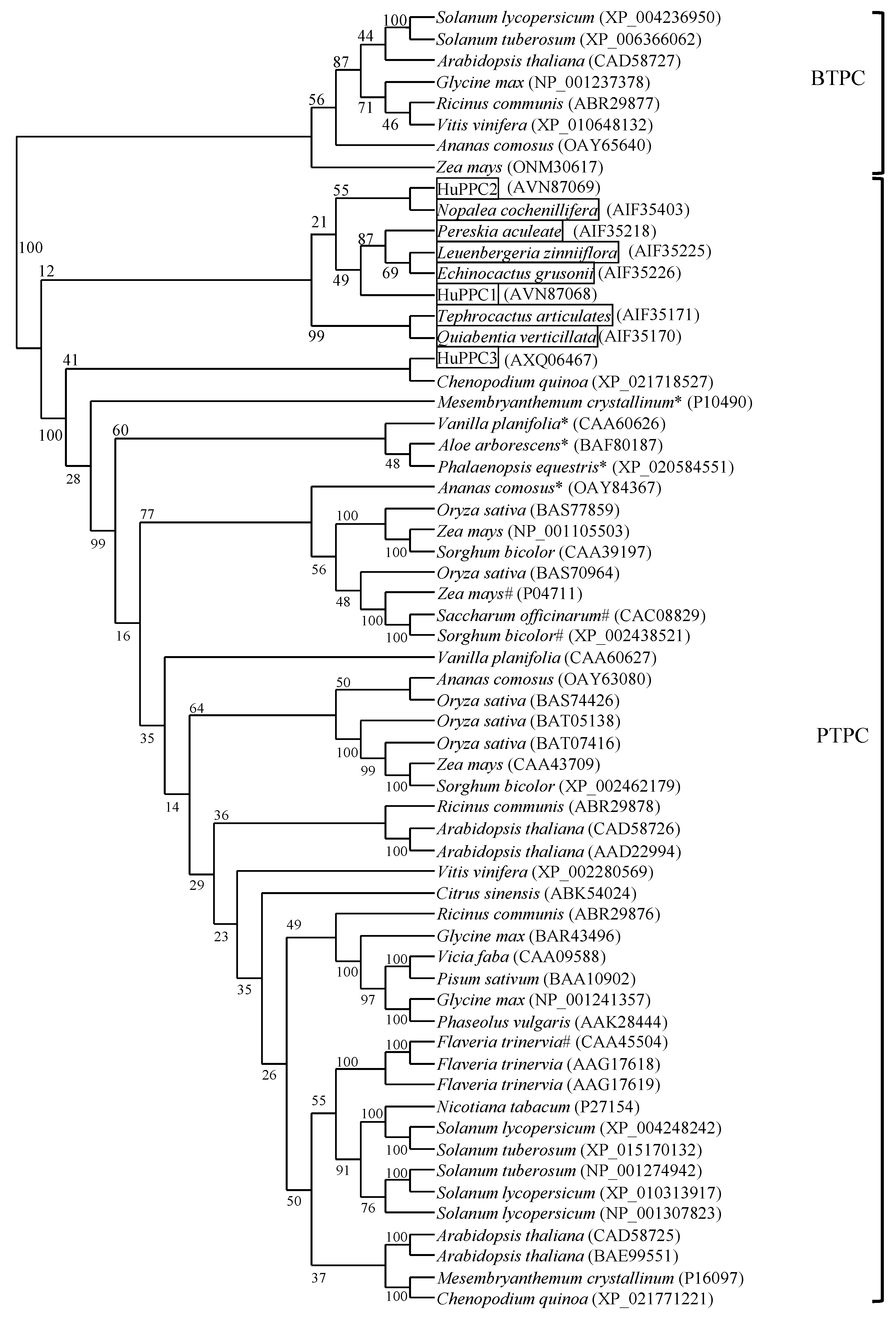
2.1. Amino Acid Sequence Alignment and Phylogenetic Analysis of PEPC Isoforms
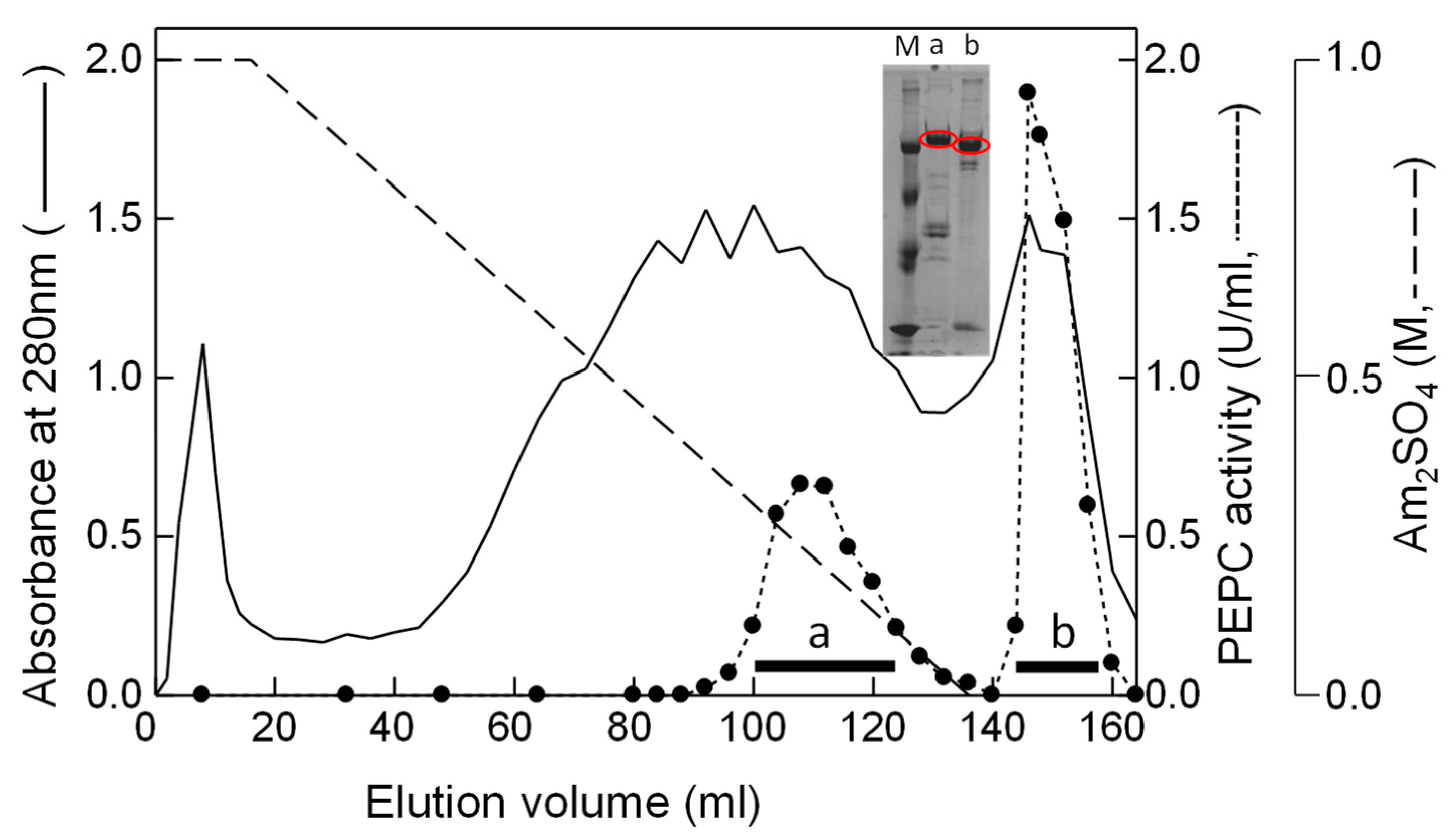
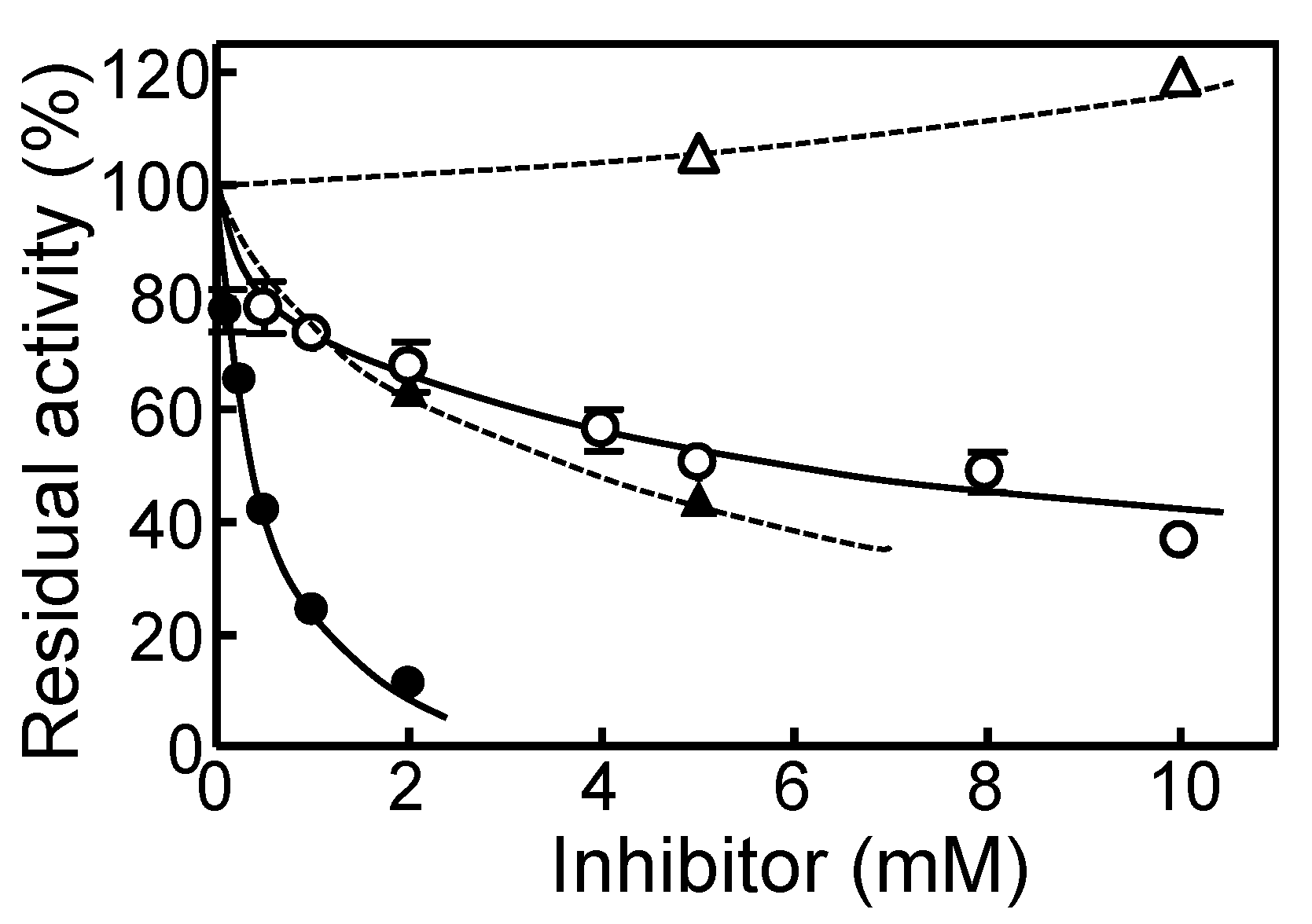
2.2. Partial Purification and Characterization of HuPPC Isoforms
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction of RNA
4.3. cDNA Cloning of PEPC
4.4. Phylogenetic Analysis
4.5. Partial Purification of HuPPC
4.5.1. Extraction and Concentration Using Anion Exchange Resin
4.5.2. Phenyl-Sepharose Column Chromatography
4.5.3. Mono Q Column Chromatography
4.5.4. Hydroxyapatite (HA) Column Chromatography
4.5.5. Mono S Column Chromatography
4.6. Enzyme Assay
4.7. MS Analysis to Identify the Amino Acid Sequence
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- O’Leary, B.; Park, J.; Plaxton, W.C. The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase): Recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs. Biochem. J. 2011, 436, 15–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latzko, E.; Kelly, G.J. The many-faceted function of phosphoenolpyruvate carboxylase in C3 plants. Physiol. Veg. 1983, 21, 805–815. [Google Scholar]
- Yanagisawa, S.; Akiyama, A.; Kisaka, U.; Uchimiya, H.; Miwa, T. Metabolic engineering with Dof1 transcription factor in plants: Improved nitrogen assimilation and growth under low-nitrogen conditions. Proc. Natl. Acad. Sci. USA 2004, 101, 7833–7838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, M.; Mai, H.T.; Fujii, M.; Hata, S.; Izui, K.; Tajima, S. Phosphoenolpyruvate carboxylase plays a crucial role in limiting nitrogen fixation in Lotus japonicas nodules. Plant Cell Physiol. 2006, 47, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Sugimoto, T.; Takano, T.; Sasou, A.; Morita, S.; Yano, K.; Masumura, T. The plant-type phosphoenolpyruvate carboxylase Gmppc2 is developmentally induced in immature soy seeds at the late maturation stage: A potential protein biomarker for seed chemical composition. Biosci. Biotech. Biochem. 2019, 84, 552–562. [Google Scholar] [CrossRef]
- Sriram, G.; Fulton, D.B.; Shanks, J.V. Flux quantification in central carbon metabolism of Catharanthus roseus hairy roots by C-13 labeling and comprehensive bondomer balancing. Phytochemistry 2007, 68, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, X.; Zhang, C.; Dai, C.; Zhou, J.; Ren, C.; Zhang, J. Phosphoenolpyruvate carboxylase regulation in C4-PEPC-expressing transgenic rice during early responsesto drought stress. Physiol. Plant. 2017, 159, 178–200. [Google Scholar] [CrossRef]
- Wikkick, I.R.; Plaxton, W.C.; Lolle, S.J.; Nacfie, S.M. Transcriptional and post-translational upregulation of phosphoenolpyruvate carboxylase in Arabidopsis thaliana (L. Heynh) under cadmium stress. Environ. Exp. Bot. 2019, 164, 29–39. [Google Scholar]
- Izui, K.; Matsumura, H.; Furumoto, T.; Kai, Y. Phosphoenolpyruvate carboxylase: A new era of structural biology. Annu. Rev. Plant Biol. 2004, 55, 69–84. [Google Scholar] [CrossRef]
- Chollet, R.; Vidal, J.; O’Leary, M.H. Phosphoenolpyruvate carboxylase: A ubiquitous, highly regulated enzyme in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 273–298. [Google Scholar] [CrossRef] [Green Version]
- Nimmo, H.G. The regulation of phosphoenolpyruvate carboxylase in CAM plants. Trends Plant Sci. 2000, 5, 75–80. [Google Scholar] [CrossRef]
- Nimmo, H.G. Control of the physophorylation of phosphoenolpyruvate carboxylase in higher plants. Arch. Biochem. Biophys. 2003, 414, 189–196. [Google Scholar] [CrossRef]
- Mallona, I.; Egea-Cortines, M.; Weiss, J. Conserved and divergent rhythms of Crassulacean acid metabolism-related and core clock gene expression in the cactus Opntia ficus-indica. Plant Physiol. 2011, 156, 1978–1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bläsing, O.E.; Ernst, K.; Streubel, M.; Westhoff, P.; Svesson, P. The non-photosynthetic phosphoenolpyruvate carboxylases of the C4 dicot Flaveria trinervia-implications for the evolution of C4 photosynthesis. Planta 2002, 215, 448–456. [Google Scholar] [CrossRef]
- Gehrig, H.H.; Wood, J.A.; Cushman, M.A.; Virga, A.; Cushman, J.C.; Winter, K. Large gene family of phosphoenolpyruvate carboxylase in the crassulacean acid metabolism plant Kalanchoe pinnata (Crassulaceae) characterized by partial cDNA sequence analysis. Funct. Plant Biol. 2005, 32, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, M.; Suzuki, R.; Yamazaki, T.; Miyano, M. Comparison of plant-type phosphoenolpyruvate carboxylases from rice: Identification of two plant-specific regulatory regions of the allosteric enzyme. Plant Cell Physiol. 2015, 56, 468–480. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.F. Strawberry Pear. In Fruit of Warm Climates; Curtis, F.D., Jr., Ed.; Media Incorporated: North Carolina, NC, USA, 1987; pp. 347–348. [Google Scholar]
- Thomson, P. Pitahaya (Hylocreus species) a Promising New Fruit Crop for Southern California; Bonsall Publications: Bonsall, CA, USA, 2002. [Google Scholar]
- Raveh, E.; Gersani, M.; Nobel, P.S. CO2 uptake and fluorescence responses for shade-tolerant cactus Hylocereus undatus under current and doubled CO2 concentrations. Physiol. Planta. 1995, 93, 505–511. [Google Scholar] [CrossRef]
- Nobel, P.S.; Barrera, E.D.L. High temperature and net CO2 uptake, growth, and stem damage for the hemiepiphytic Cactus Hylocereus Undatus. Biotropica 2002, 34, 225–231. [Google Scholar] [CrossRef]
- Nomura, K.; Ide, M.; Ashida, T.; Ynonemoto, Y. Seasonal defference in diurnal patterns of metabolites and enzyme activities in pitaya (Hylocereus undatus) grown in a temperate zone. J. Jpn. Soc. Hort. Sci. 2010, 79, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Uhrig, R.G.; She, Y.; Leach, C.A.; Plaxton, W.C. Regulatory monoubiquitination of phosphoenolpyruvate carboxylase in germinating castor oil seeds. J. Biol. Chem. 2008, 283, 29650–29657. [Google Scholar] [CrossRef] [Green Version]
- Kai, Y.; Matsumura, H.; Izui, K. Phosphoenolpyruvate carboxylase: Three-dimensional structure and molecular mechanisms. Arch. Biochem. Biophys. 2003, 414, 170–179. [Google Scholar] [CrossRef]
- Fan, Q.J.; Yan, F.X.; Qiao, G.; Zhang, B.X.; Wen, X.P. Identification of differentially-expressed genes potentially implicated in drought response in pitaya (Hylocereus undatus) by suppression subtractive hybridization and cDNA microarray analysis. Gene 2014, 533, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, H.; Xie, Y.; Shirakata, S.; Inoue, T.; Yoshinaga, T.; Ueno, Y.; Izui, K.; Kai, Y. Crystal structures of C4 form maize and quaternary complex of E. coli phosphoenolpyruvate carboxylases. Structure 2002, 10, 1721–1730. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Terada, A.; Kotera, M.; Ohshima, K.; Furumoto, T.; Matsumura, H.; Kai, Y.; Izui, K. Maize phosphoenolpyruvate carboxylase. Mutation at the putative binding site for glucose-6-phosphate caused desensitization and abolish responsiveness to regulatory phosphorylation. J. Biol. Chem. 2005, 280, 11798–11806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldous, S.H.; Weise, S.E.; Sharkey, T.D.; Waldera-Lupa, D.M.; Stühler, K.; Mallmann, J.; Groth, G.; Gowik, U.; Westhoff, P.; Arsova, B. Evolution of the phosphoenolpyruvate carboxylase protein kinase family in C3 and C4 Flaveria spp. Plant Physiol. 2014, 165, 1076–1091. [Google Scholar] [CrossRef] [Green Version]
- Ping, C.Y.; Chen, F.C.; Cheng, T.C.; Lin, H.L.; Lin, T.S.; Yang, W.J.; Lee, Y.I. Expression profiles of phosphoenolpyruvate carboxylase and phosphoenolpyruvate carboxylase kinase genes in Phalaenopsis, implications for regulating the performance of crassulacean acid metabolism. Front. Plant Sci. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Paulus, J.K.; Schlieper, D.; Groth, G. Greater efficiency of photosynthetic carbon fixation due to single amino-acid substitution. Nat. Commun. 2013, 4, 1518. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.; Engelmann, S.; Westhoff, P.; Gowik, U. Evolution of C4 phosphoenolpyruvate carboxylase in Flaveria: Determination for high tolerance towards the inhibitor L-malate. Plant Cell Environ. 2008, 31, 793–803. [Google Scholar] [CrossRef]
- Engelmann, S.; Blässing, O.E.; Westhoff, P.; Svensson, P. Serine 774 and amino acids 296 to 437 comprise the major C4 determinants of the C4 phosphoenolpyruvate carboxylase of Flaveria trinervia. FEBS Lett. 2002, 524, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Rosnow, J.J.; Edwards, G.E.; Roalson, E.H. Positive selection of Kranz and non-Kranz C4 phosphoenolpyruvate carboxylase amino acids in Suaedoideae (Chenopodiaceae). J. Exp. Bot. 2014, 65, 3595–3607. [Google Scholar] [CrossRef] [Green Version]
- DiMario, R.J.; Cousins, A.B. A single serine to alanine substitution decreases bicarbonate affinity of phosphoenolpyruvate carboxylase in C4 Flaveria trinervia. J. Exp. Bot. 2019, 70, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geissler, N.; Hussin, S.; El-Far, M.M.M.; Koyro, H.-W. Elevated atmospheric CO2 concentration leads to different salt resistance mechanism in a C3 (Chenopodium quinoa) and a C4 (Atriplex nummularia) halophyte. Envirn. Exp. Bot. 2015, 118, 67–77. [Google Scholar] [CrossRef]
- Chirgwin, J.M.; Przybyla, A.E.; MacDonald, R.J.; Rutter, W.J. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 1979, 18, 5294–5299. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Bensadoun, A.; Weinstein, D. Assay of Proteins in the Presence of Interfering Materials. Anal. Biochem. 1976, 70, 241–250. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Activity (U) | Recovery (%) | Specific Activity (U/mg) | Total Protein (mg) | Purification Fold | |
|---|---|---|---|---|---|
| Extract | 33.3 | 100 | 0.0318 | 1050 | 1 |
| Q-Sepharose | 24.3 | 73.0 | 0.122 | 199 | 3.84 |
| Phenyl-Sepharose-a | 9.10 | 27.4 | 0.315 | 28.9 | 9.91 |
| Phenyl-Sepharose-b | 11.3 | 34.1 | 0.486 | 23.3 | 15.3 |
| Mono Q-a | 7.18 | 21.6 | 2.19 | 3.28 | 68.9 |
| Mono Q-b | 8.17 | 24.6 | 0.836 | 9.78 | 26.3 |
| Hydroxyapatite-a | 4.37 | 13.1 | 3.87 | 1.13 | 122 |
| Hydroxyapatite-b | 7.43 | 22.3 | 1.49 | 5.00 | 46.9 |
| Mono S-a | 4.24 | 12.7 | 7.45 | 0.256 | 234 |
| Mono S-b | 1.91 | 5.73 | 1.07 | 2.38 | 33.6 |
| HuPPC1 | HuPPC2 | HuPPC3 | |
|---|---|---|---|
| HuPPCa | 12% | 18% | 75% |
| HuPPCb | 10% | 67% | 13% |
| HuPPC2 | HuPPC3 | |
|---|---|---|
| pH stability | 6.5–7.0 | 5.5–6.5 |
| Heat stability | Up to 35 °C | Up to 50 °C |
| Optimal pH | 6.5–7.2 | 7.0–7.2 |
| Optimal temperature | 28 °C | 55 °C |
| Km,PEP (μM) | 44.3 ± 7.0 | 48.5 ± 4.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nomura, K.; Sakurai, Y.; Dozono, M. Molecular Cloning of Novel-Type Phosphoenolpyruvate Carboxylase Isoforms in Pitaya (Hylocereus undatus). Plants 2020, 9, 1241. https://doi.org/10.3390/plants9091241
Nomura K, Sakurai Y, Dozono M. Molecular Cloning of Novel-Type Phosphoenolpyruvate Carboxylase Isoforms in Pitaya (Hylocereus undatus). Plants. 2020; 9(9):1241. https://doi.org/10.3390/plants9091241
Chicago/Turabian StyleNomura, Keiichi, Yuho Sakurai, and Mayu Dozono. 2020. "Molecular Cloning of Novel-Type Phosphoenolpyruvate Carboxylase Isoforms in Pitaya (Hylocereus undatus)" Plants 9, no. 9: 1241. https://doi.org/10.3390/plants9091241