Bioactive Compounds from Polygala tenuifolia and Their Inhibitory Effects on Lipopolysaccharide-Stimulated Pro-inflammatory Cytokine Production in Bone Marrow-Derived Dendritic Cells

 , and
, and
Abstract
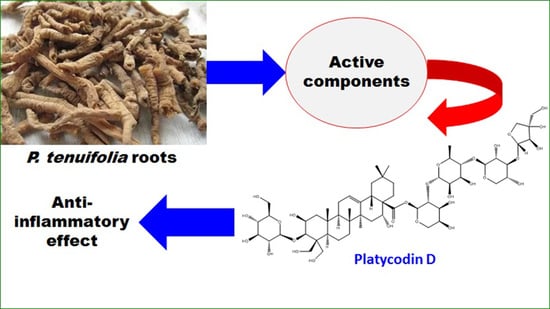
:
1. Introduction
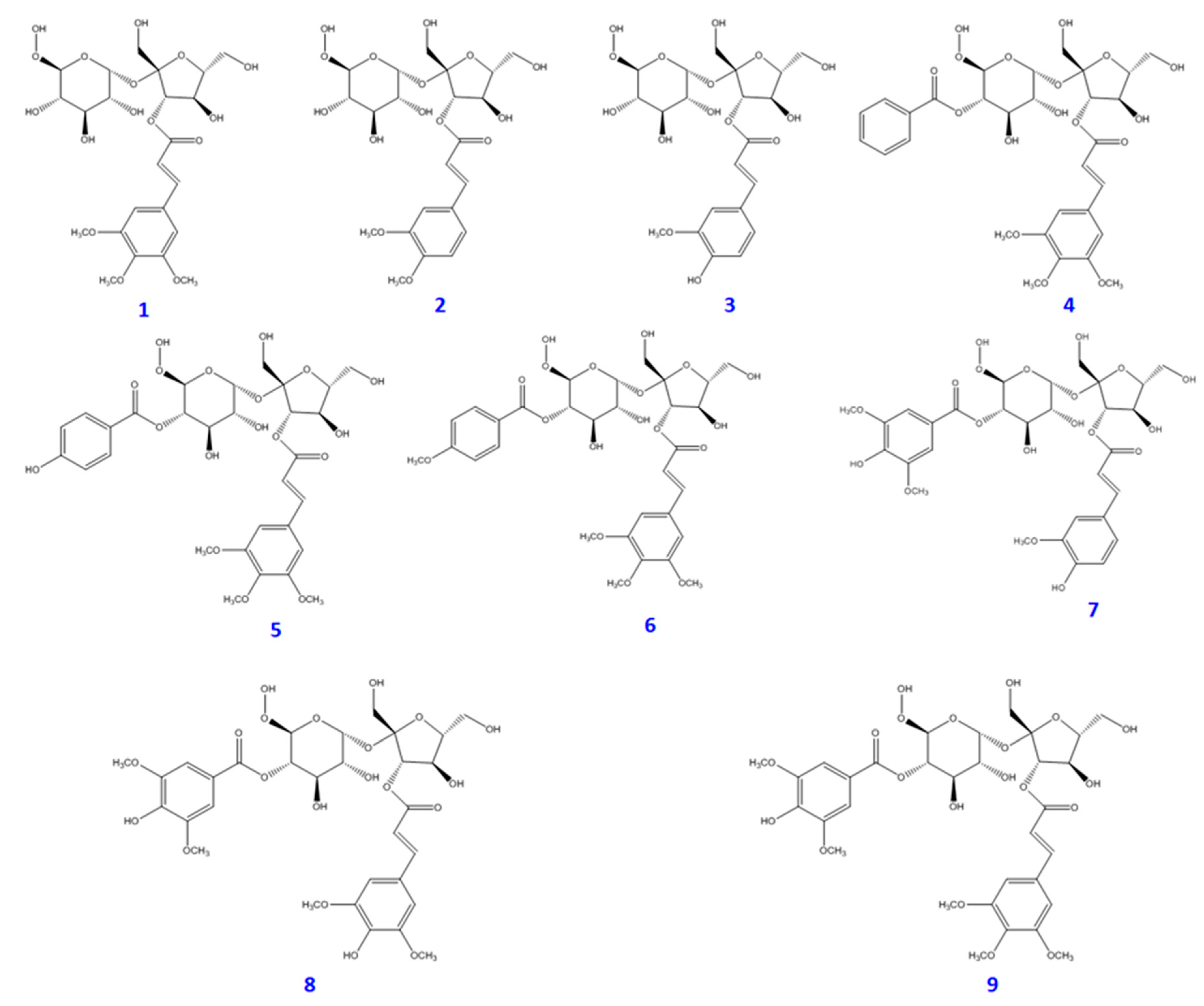
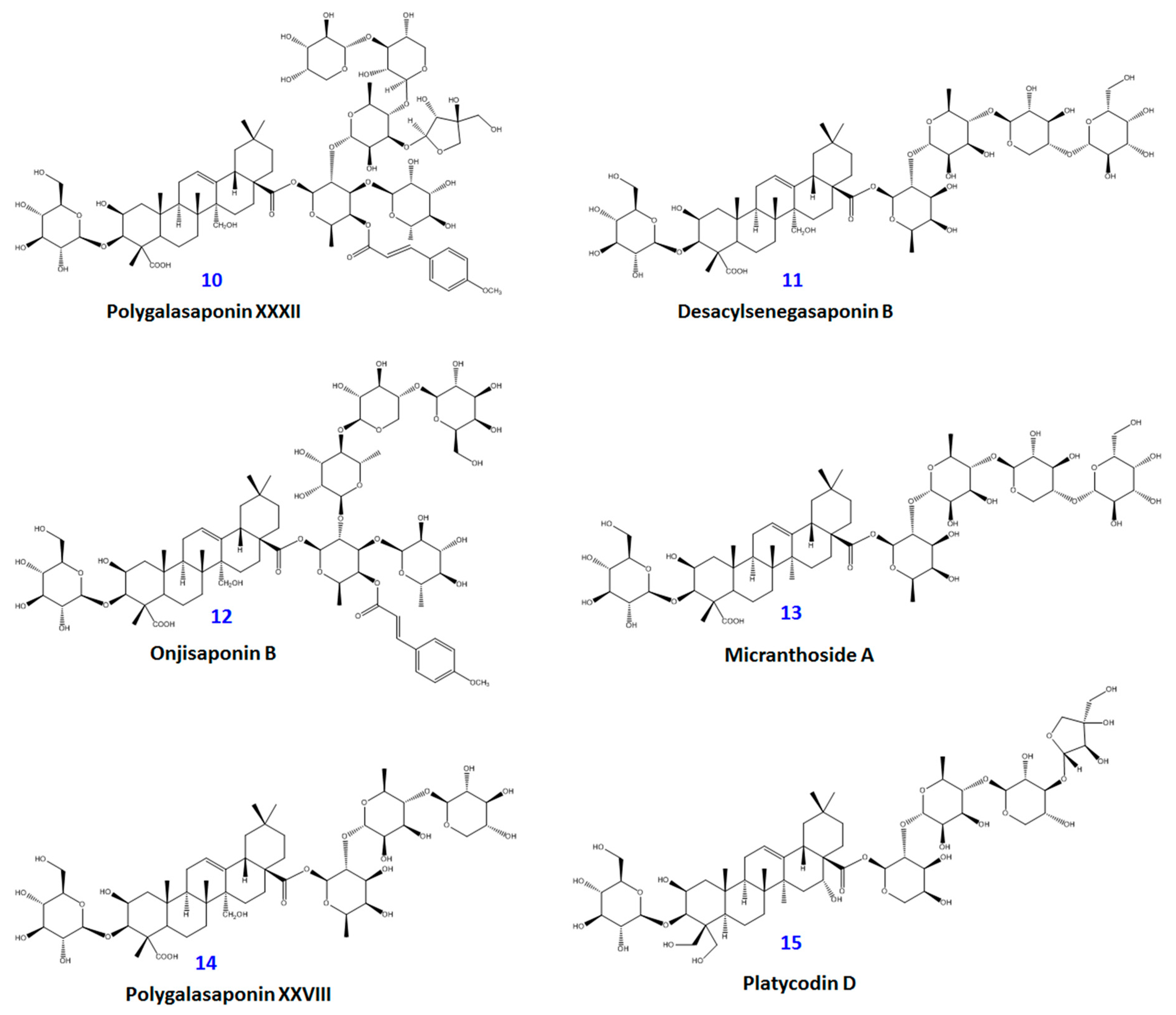
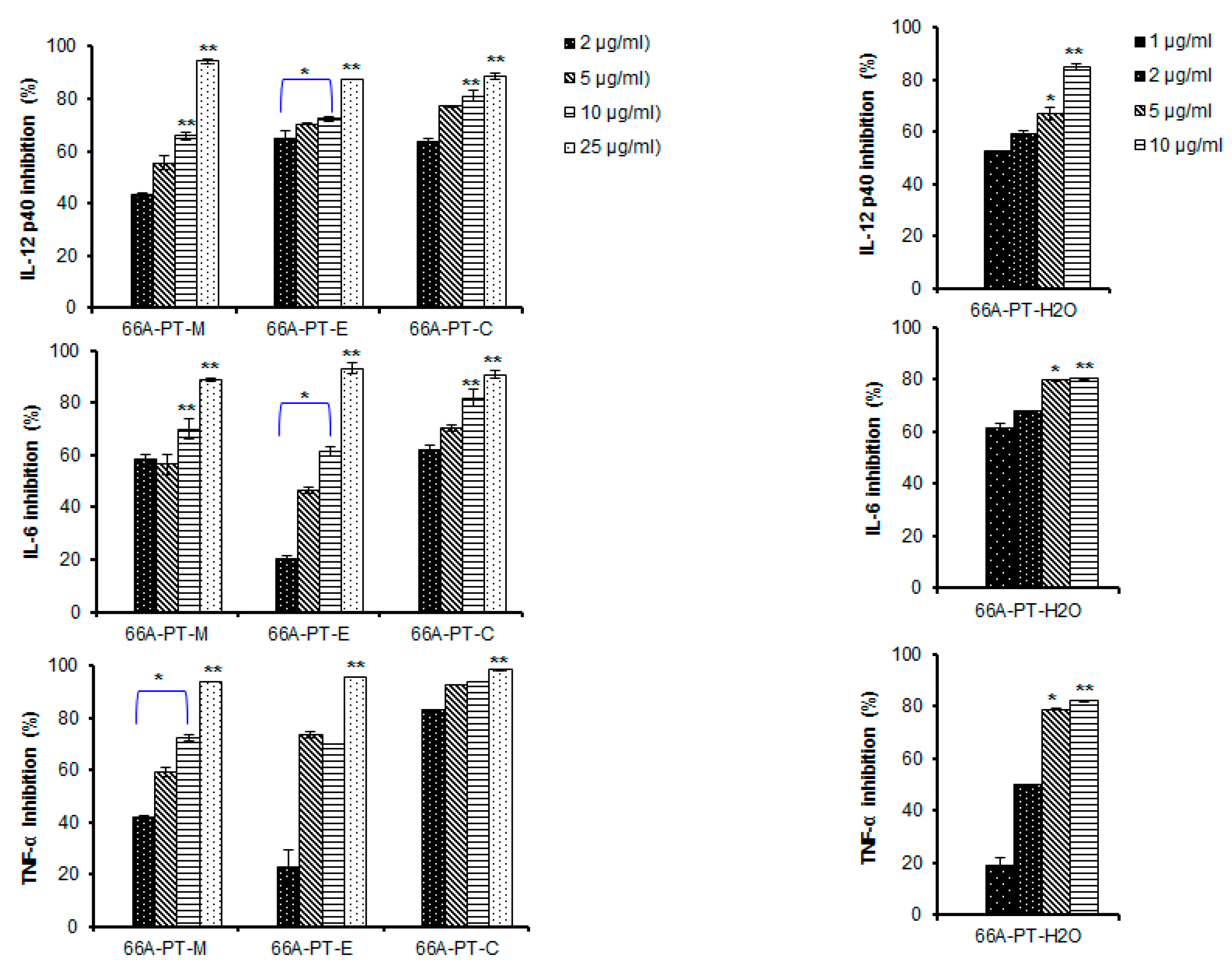
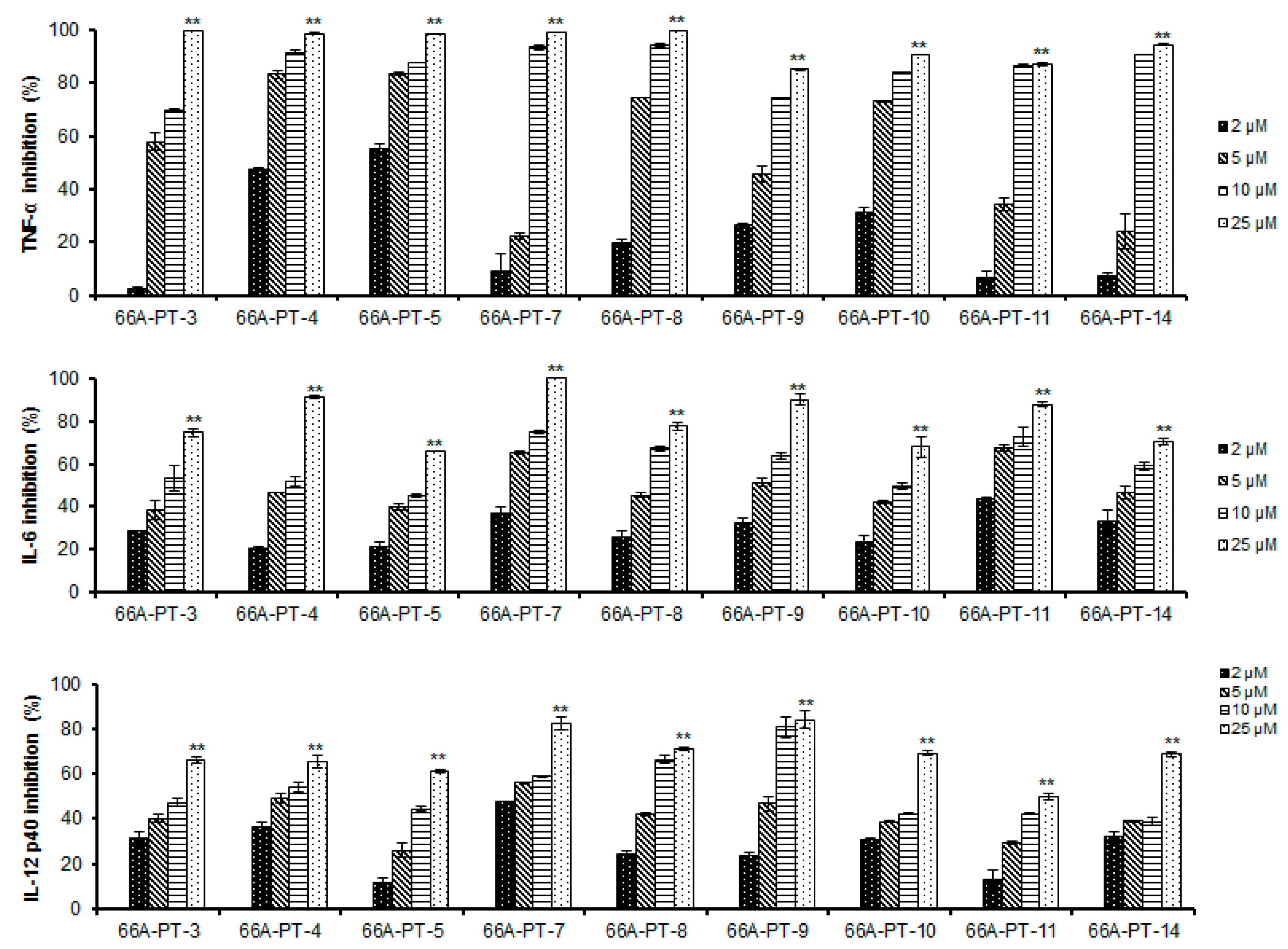
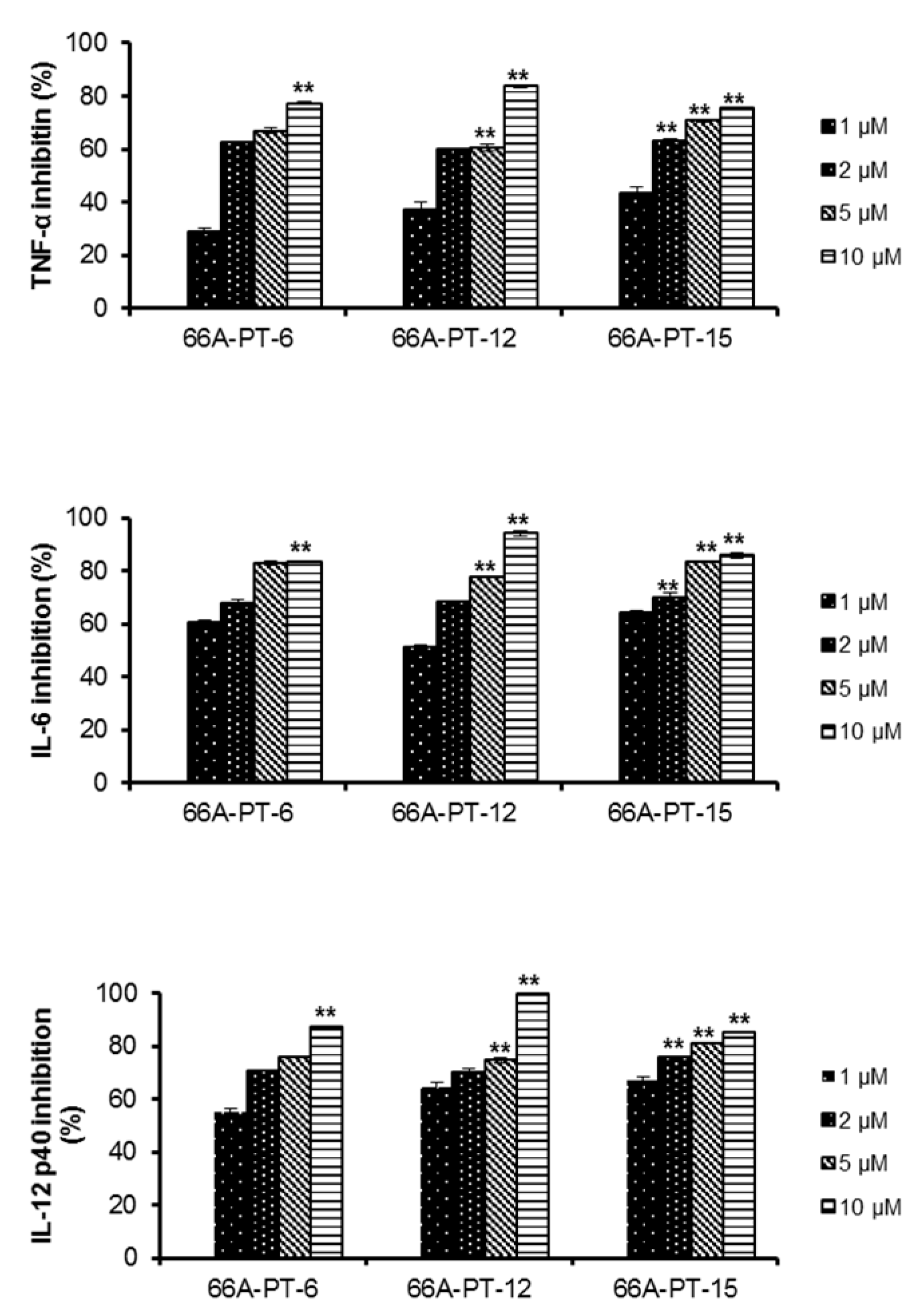
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Extraction and Isolation
Physical Properties and Key Spectroscopic Data of Isolated Compounds:
3.4. Cell Culture and Reagents
3.5. Cytokine Production Measurements
3.6. Cell Viability Assay
3.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, J.-M.; An, J. Cytokines, inflammation and pain. Int. Anesthesiol. Clin. 2007, 45, 27. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Tang, Y.; Hua, S. Immunological approaches towards cancer and inflammation: A cross talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Vinh, L.B.; Lee, Y.; Han, Y.K.; Kang, J.S.; Park, J.U.; Kim, Y.R.; Yang, S.Y.; Kim, Y.H. Two new dammarane-type triterpene saponins from Korean red ginseng and their anti-inflammatory effects. Bioorg. Med. Chem. Lett. 2017, 27, 5149–5153. [Google Scholar] [CrossRef] [PubMed]
- Vinh, L.B.; Jo, S.J.; Nguyen Viet, P.; Gao, D.; Cho, K.W.; Koh, E.-J.; Park, S.S.; Kim, Y.H.; Yang, S.Y. The chemical constituents of ethanolic extract from Stauntonia hexaphylla leaves and their anti-inflammatory effects. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef]
- Vinh, L.B.; Jang, H.-J.; Phong, N.V.; Cho, K.W.; Park, S.S.; Kang, J.S.; Kim, Y.H.; Yang, S.Y. Isolation, structural elucidation, and insights into the anti-inflammatory effects of triterpene saponins from the leaves of Stauntonia hexaphylla. Bioorg. Med. Chem. Lett. 2019, 29, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Cheong, M.-H.; Lee, S.-R.; Yoo, H.-S.; Jeong, J.-W.; Kim, G.-Y.; Kim, W.-J.; Jung, I.-C.; Choi, Y.H. Anti-inflammatory effects of Polygala tenuifolia root through inhibition of NF-κB activation in lipopolysaccharide-induced BV2 microglial cells. J. Ethnopharmacol. 2011, 137, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, J.; Yu, S.; Chen, N.; Xue, W.; Hu, J.; Zhang, D. Triterpenoid saponins with neuroprotective effects from the roots of Polygala tenuifolia. Planta Med. 2008, 74, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Tu, P.-F. Xanthone O-glycosides from Polygala tenuifolia. Phytochemistry 2002, 60, 813–816. [Google Scholar] [CrossRef]
- Shi, T.-X.; Wang, S.; Zeng, K.-W.; Tu, P.-F.; Jiang, Y. Inhibitory constituents from the aerial parts of Polygala tenuifolia on LPS-induced NO production in BV2 microglia cells. Bioorg. Med. Chem. Lett. 2013, 23, 5904–5908. [Google Scholar] [CrossRef]
- Dong, X.-Z.; Huang, C.-L.; Yu, B.-Y.; Hu, Y.; Mu, L.-H.; Liu, P. Effect of Tenuifoliside A isolated from Polygala tenuifolia on the ERK and PI3K pathways in C6 glioma cells. Phytomedicine 2014, 21, 1178–1188. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.; Dang, T.T.; Nguyen, P.H.; Kim, E.; Thuong, P.T.; Oh, W.K. Xanthones from Polygala karensium inhibit neuraminidases from influenza A viruses. Bioorg. Med. Chem. Lett. 2012, 22, 3688–3692. [Google Scholar] [CrossRef] [PubMed]
- Vinh, L.B.; Jang, H.-J.; Phong, N.V.; Dan, G.; Cho, K.W.; Kim, Y.H.; Yang, S.Y. Bioactive triterpene glycosides from the fruit of Stauntonia hexaphylla and insights into the molecular mechanism of its inflammatory effects. Bioorg. Med. Chem. Lett. 2019, 29, 2085–2089. [Google Scholar] [CrossRef] [PubMed]
- Vinh, L.B.; Nguyet, N.T.M.; Ye, L.; Dan, G.; Phong, N.V.; Anh, H.L.T.; Kim, Y.H.; Kang, J.S.; Yang, S.Y.; Hwang, I. Enhancement of an in vivo anti-inflammatory activity of oleanolic acid through glycosylation occurring naturally in Stauntonia hexaphylla. Molecules 2020, 25, 3699. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Doan, H.T.; Vu, T.V.; Pham, Q.T.; Khoi, N.M.; Huu, T.N.; Thuong, P.T. Oligosaccharide and glucose esters from the roots of Polygala arillata. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef]
- Song, Y.-L.; Zhou, G.-S.; Zhou, S.-X.; Jiang, Y.; Tu, P.-F. Polygalins D–G, four new flavonol glycosides from the aerial parts of Polygala sibirica L.(Polygalaceae). Nat. Prod. Res. 2013, 27, 1220–1227. [Google Scholar] [CrossRef]
- Quang, T.H.; Yen, D.T.H.; Nhiem, N.X.; Tai, B.H.; Ngan, N.T.T.; Anh, H.L.T.; Oh, H.; Van Kiem, P.; Van Minh, C. Oleanane-type triterpenoid saponins from the roots of Polygala aureocauda Dunn. Phytochem. Let. 2019, 34, 59–64. [Google Scholar] [CrossRef]
- Ali, I.; Manzoor, Z.; Koo, J.-E.; Kim, J.-E.; Byeon, S.-H.; Yoo, E.-S.; Kang, H.-K.; Hyun, J.-W.; Lee, N.-H.; Koh, Y.-S. 3-Hydroxy-4, 7-megastigmadien-9-one, isolated from Ulva pertusa, attenuates TLR9-mediated inflammatory response by down-regulating mitogen-activated protein kinase and NF-κB pathways. Pharm. Biol. 2017, 55, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Vinh, L.B.; Nguyet, N.T.M.; Yang, S.Y.; Kim, J.H.; Thanh, N.V.; Cuong, N.X.; Nam, N.H.; Minh, C.V.; Hwang, I.; Kim, Y.H. Cytotoxic triterpene saponins from the mangrove Aegiceras corniculatum. Nat. Prod. Res. 2017, 33, 628–634. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, H.Y.; Yang, S.Y.; Kim, J.-B.; Jin, C.H.; Kim, Y.H. Inhibitory activity of (−)-epicatechin-3, 5-O-digallate on α-glucosidase and in silico analysis. Int. J. Biol. Macromol. 2018, 107, 1162–1167. [Google Scholar] [CrossRef]
- Sun, Y.N.; Kim, J.H.; Li, W.; Jo, A.R.; Yan, X.T.; Yang, S.Y.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of anthraquinone components from Aloe. Bioorg. Med. Chem. 2015, 23, 6659–6665. [Google Scholar] [CrossRef] [PubMed]
- Tung, N.H.; Song, G.Y.; Nhiem, N.X.; Ding, Y.; Tai, B.H.; Jin, L.G.; Lim, C.-M.; Hyun, J.W.; Park, C.J.; Kang, H.K. Dammarane-type saponins from the flower buds of Panax ginseng and their intracellular radical scavenging capacity. J. Agric. Food Chem. 2009, 58, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.; Cuong, N.; Luyen, B.; Quang, T.; Hanh, T.; Kim, S.; Koh, Y.-S.; Nam, N.; Van Kiem, P.; Van Minh, C. Anti-inflammatory components of the starfish Astropecten polyacanthus. Mar. Drugs 2013, 11, 2917–2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ba Vinh, L.; Thi Minh Nguyet, N.; Young Yang, S.; Hoon Kim, J.; Thi Vien, L.; Thi Thanh Huong, P.; Van Thanh, N.; Xuan Cuong, N.; Hoai Nam, N.; Van Minh, C. A new rearranged abietane diterpene from Clerodendrum inerme with antioxidant and cytotoxic activities. Nat. Prod. Res. 2018, 32, 2001–2007. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | IC50 ± SD (µg/mL) a | |||
|---|---|---|---|---|
| IL-12 p40 | IL-6 | TNF-α | ||
| 1 | MeOH extract | 3.38 ± 0.02 | 1.65 ± 0.16 | 3.09 ± 0.14 |
| 2 | Water-layer | 0.94 ± 0.04 | 0.24 ± 0.08 | 2.43 ± 0.22 |
| 3 | EtOAc fraction | 0.05 ± 0.01 | 5.83 ± 0.26 | 3.92 ± 0.13 |
| 4 | CH2Cl2 fraction | 0.37 ± 0.10 | 0.89 ± 0.12 | 0.005 ± 0.001 |
| SB203580 b | 5.00 ± 0.08 | 3.50 ± 0.08 | 7.20 ± 0.06 | |
| Compounds | IC50 (µM) a | |||
|---|---|---|---|---|
| IL-12 p40 | IL-6 | TNF-α | ||
| 3 | 9.25 ± 0.06 | 7.54 ± 0.08 | 5.77 ± 0.12 | |
| 4 | 6.21 ± 0.27 | 6.44 ± 0.32 | 1.50 ± 0.25 | |
| 5 | 14.34 ± 0.03 | 2.36 ± 0.08 | 1.04 ± 0.12 | |
| 6 | 0.55 ± 0.03 | 0.35 ± 0.03 | 1.97 ± 0.03 | |
| 7 | 2.99 ± 0.60 | 3.19 ± 0.03 | 6.08 ± 0.10 | |
| 8 | 6.65 ± 0.03 | 5.82 ± 0.03 | 3.51 ± 0.03 | |
| 9 | 5.89 ± 0.08 | 4.64 ± 0.08 | 5.08 ± 0.10 | |
| 10 | 9.78 ± 0.09 | 9.04 ± 0.05 | 2.99 ± 0.29 | |
| 11 | 21.05 ± 0.40 | 2.42 ± 0.03 | 6.34 ± 0.12 | |
| 12 | 0.43 ± 0.05 | 0.83 ± 0.09 | 1.75 ± 0.02 | |
| 14 | 10.52 ± 0.10 | 6.05 ± 0.03 | 2.05 ± 0.02 | |
| 15 | 0.08 ± 0.01 | 0.24 ± 0.06 | 1.17 ± 0.13 | |
| SB203580 b | 5.00 ± 0.01 | 3.50 ± 0.02 | 7.20 ± 0.02 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinh, L.B.; Heo, M.; Phong, N.V.; Ali, I.; Koh, Y.S.; Kim, Y.H.; Yang, S.Y. Bioactive Compounds from Polygala tenuifolia and Their Inhibitory Effects on Lipopolysaccharide-Stimulated Pro-inflammatory Cytokine Production in Bone Marrow-Derived Dendritic Cells. Plants 2020, 9, 1240. https://doi.org/10.3390/plants9091240
Vinh LB, Heo M, Phong NV, Ali I, Koh YS, Kim YH, Yang SY. Bioactive Compounds from Polygala tenuifolia and Their Inhibitory Effects on Lipopolysaccharide-Stimulated Pro-inflammatory Cytokine Production in Bone Marrow-Derived Dendritic Cells. Plants. 2020; 9(9):1240. https://doi.org/10.3390/plants9091240
Chicago/Turabian StyleVinh, Le Ba, Myungsook Heo, Nguyen Viet Phong, Irshad Ali, Young Sang Koh, Young Ho Kim, and Seo Young Yang. 2020. "Bioactive Compounds from Polygala tenuifolia and Their Inhibitory Effects on Lipopolysaccharide-Stimulated Pro-inflammatory Cytokine Production in Bone Marrow-Derived Dendritic Cells" Plants 9, no. 9: 1240. https://doi.org/10.3390/plants9091240