Inhibitory Mechanism of Trichoderma virens ZT05 on Rhizoctonia solani

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Antagonistic Effect of T. virens ZT05 against R. solani
2.3. Inhibitory Effect of the Metabolites of T. virens ZT05 on R. solani
2.3.1. Inhibitory Effect of Nonvolatile Metabolites of T. virens ZT05 on R. solani
2.3.2. Effects of Nonvolatile Metabolites of T. virens ZT05 on the Stress Resistance of R. solani
2.3.3. Inhibitory Effect of Volatile Metabolites of T. virens ZT05 on R. solani
2.4. Transcriptome Sequencing Analysis of the Side-by-Side Cultured T. virens and R. solani
2.4.1. Mycelium Collection and Total RNA Extraction
2.4.2. Library Preparation and Illumina Sequencing
2.4.3. Differential Expression Analysis and Function Gene Selection
2.5. Validation of Quantitative qRT-PCR of the Transcriptome Data
2.6. Data Analysis
3. Results
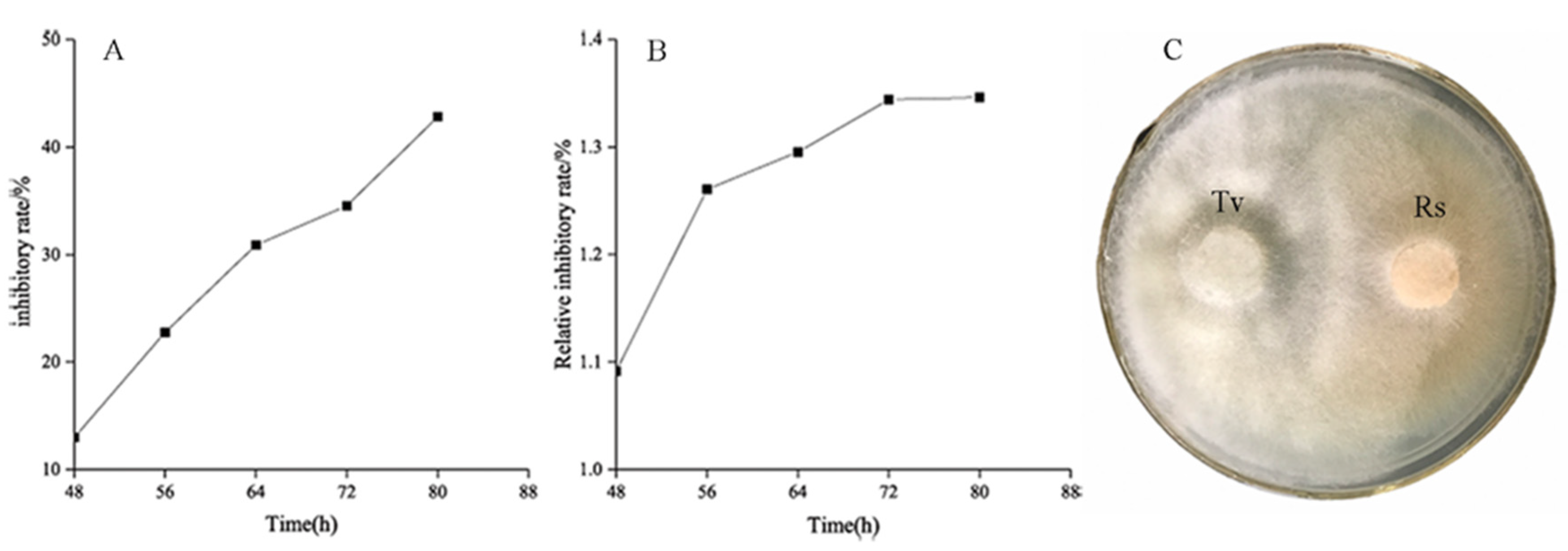
3.1. Antagonistic Effect of T. virens ZT05 against R. solani
3.2. Inhibitory Effect of the Metabolites of T. virens ZT05 on R. solani
3.2.1. Inhibitory Effect of Nonvolatile Metabolites of T. virens ZT05 on R. solani
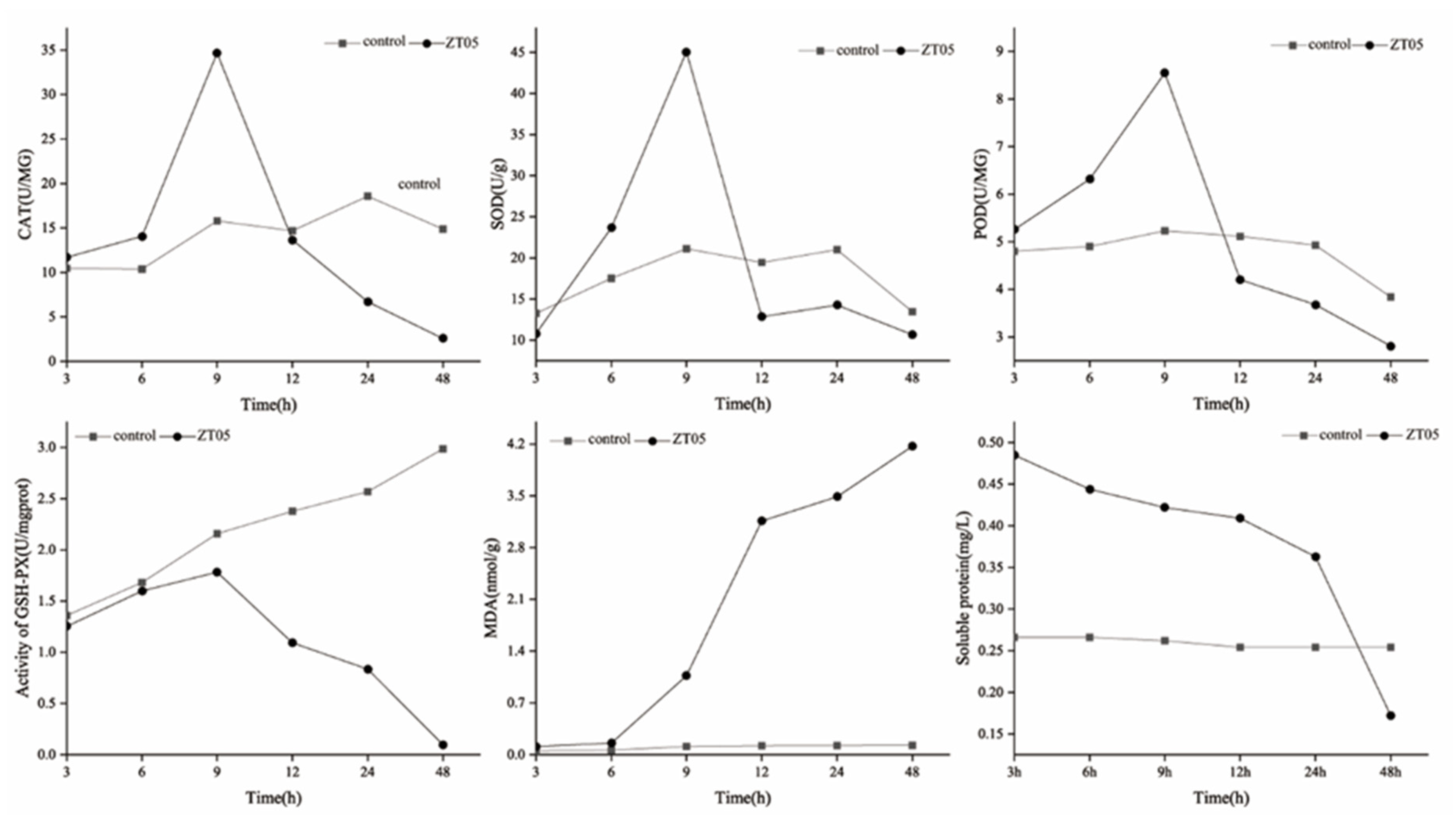
3.2.2. Inhibitory Effect of Nonvolatile Metabolites of T. virens ZT05 on Enzyme Activities of R. solani
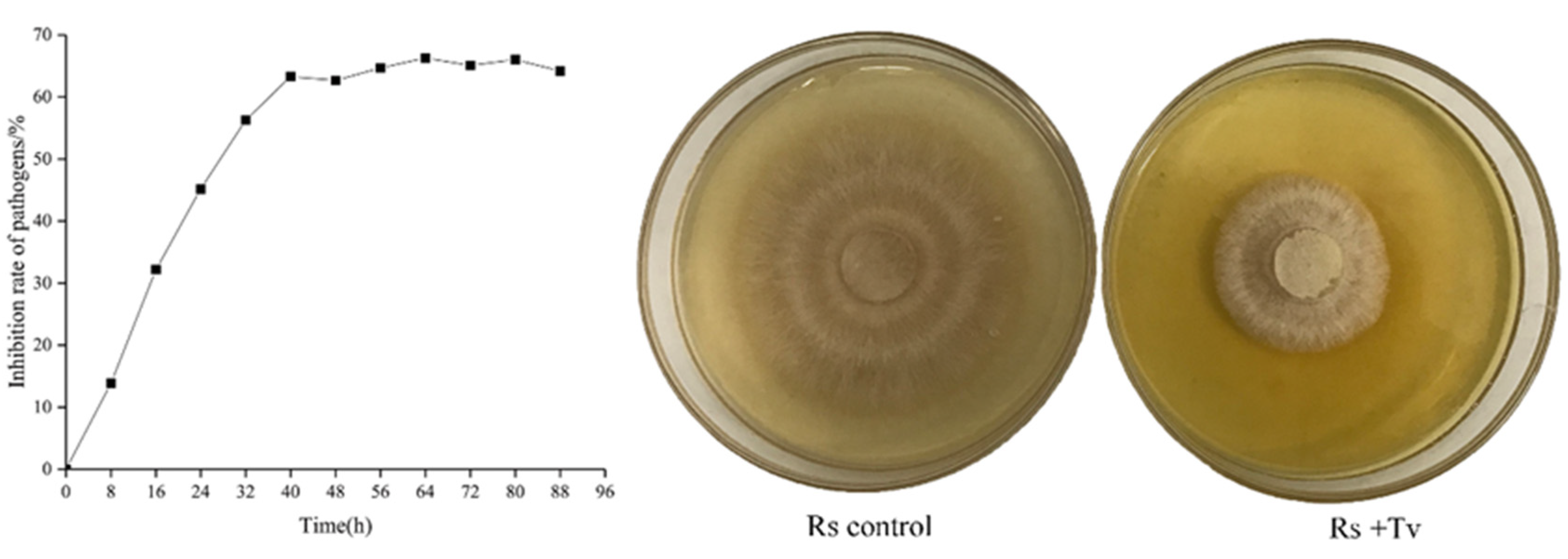
3.2.3. Inhibitory Effect of Volatile Metabolites of T. virens ZT05 on R. solani
3.3. Transcriptome Sequencing Analysis of the Side-by-Side Cultured T. virens ZT05 and R. solani
3.3.1. Analysis of Transcriptome Sequencing Data
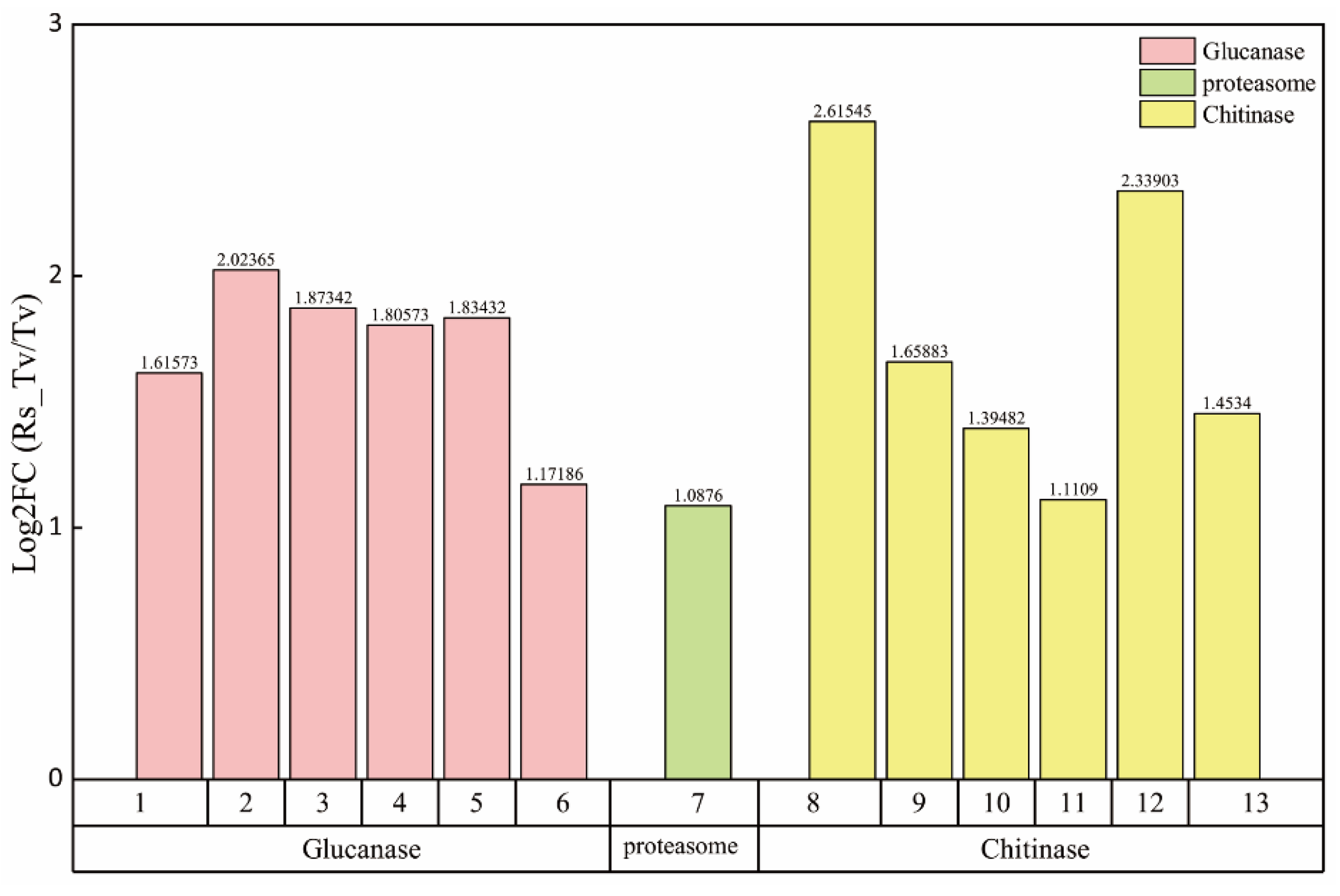
3.3.2. Screening of Hyperparasitic Functional Genes
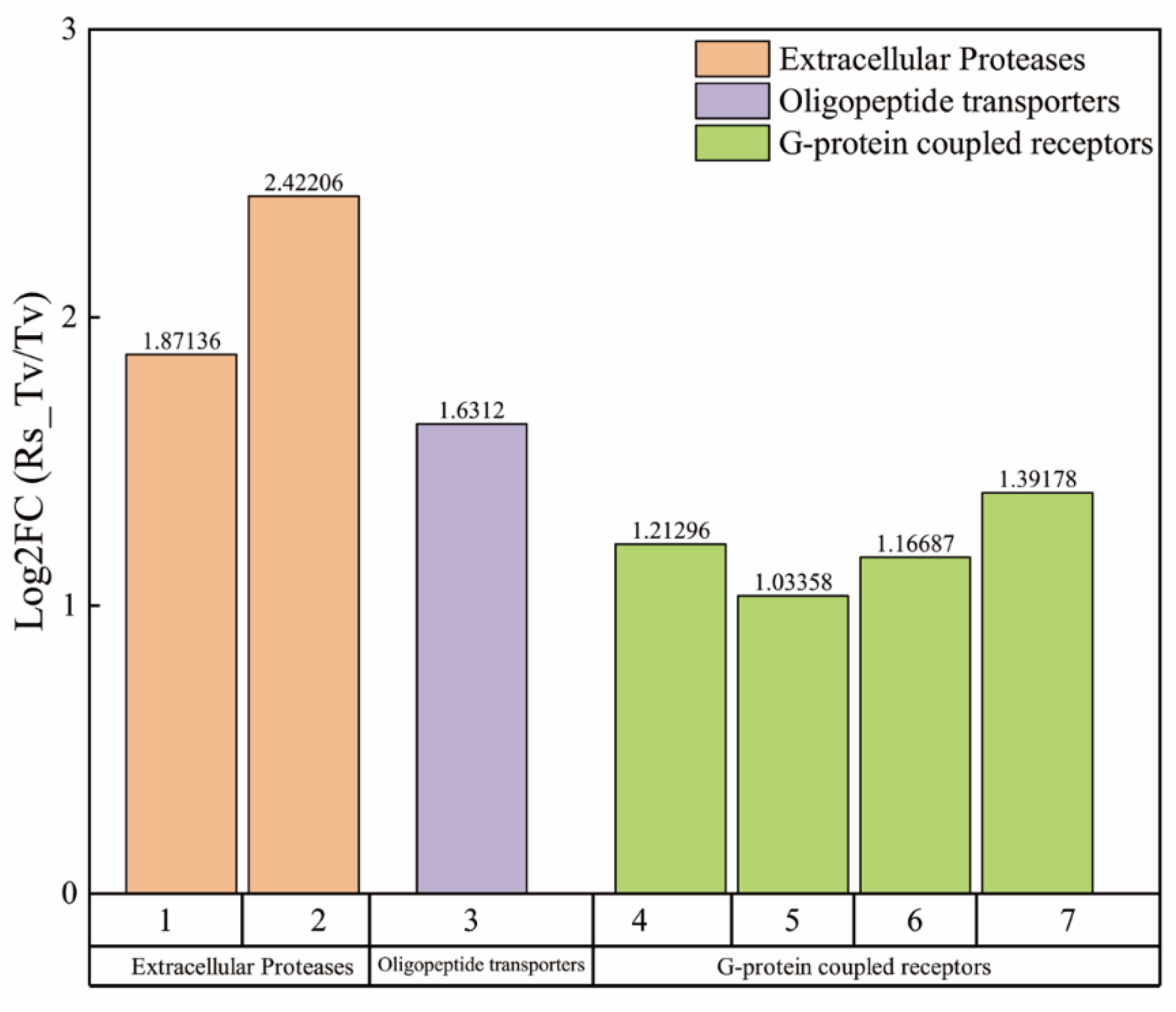
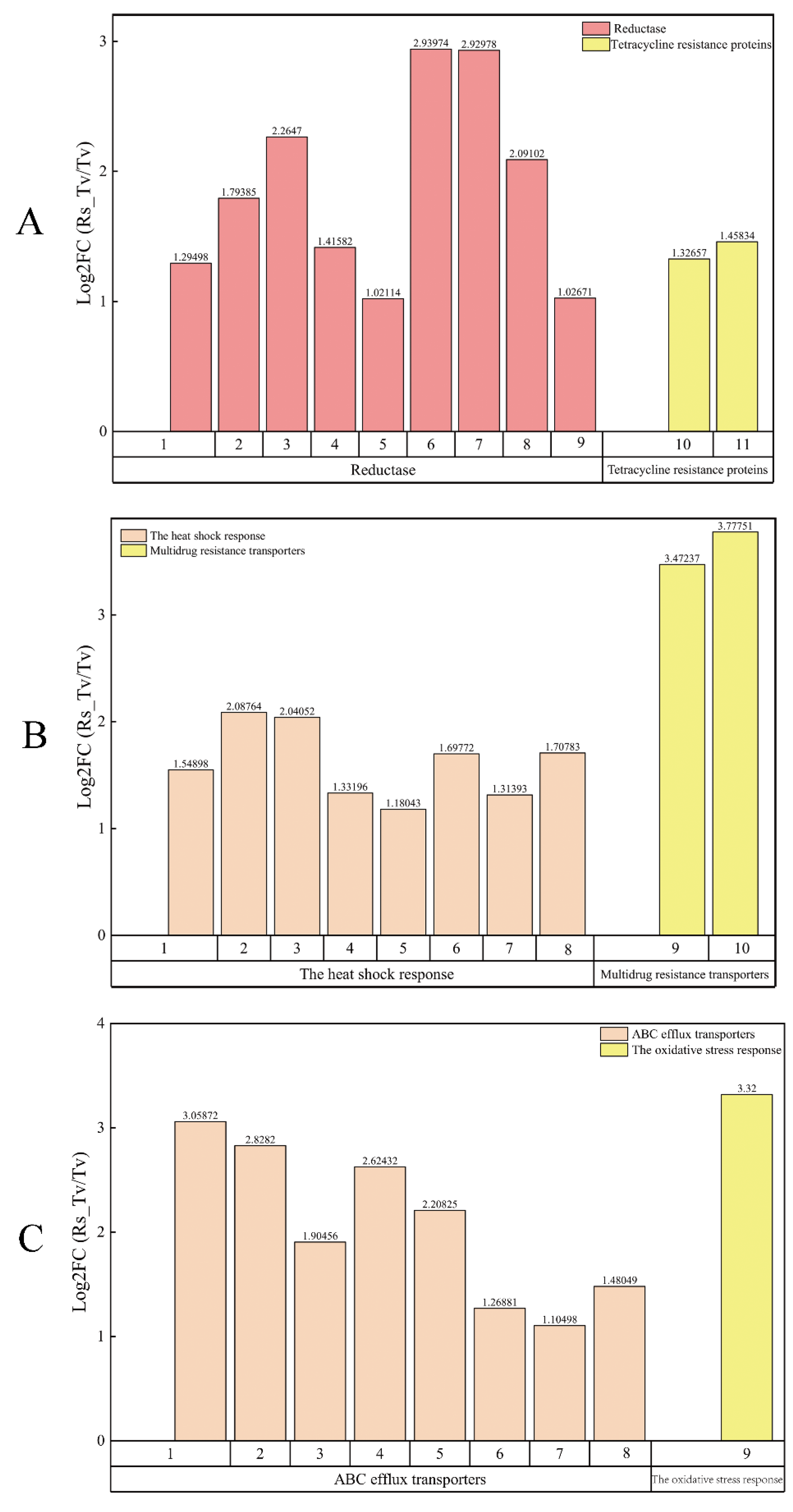
3.3.3. Screening of Antibiosis Functional Genes
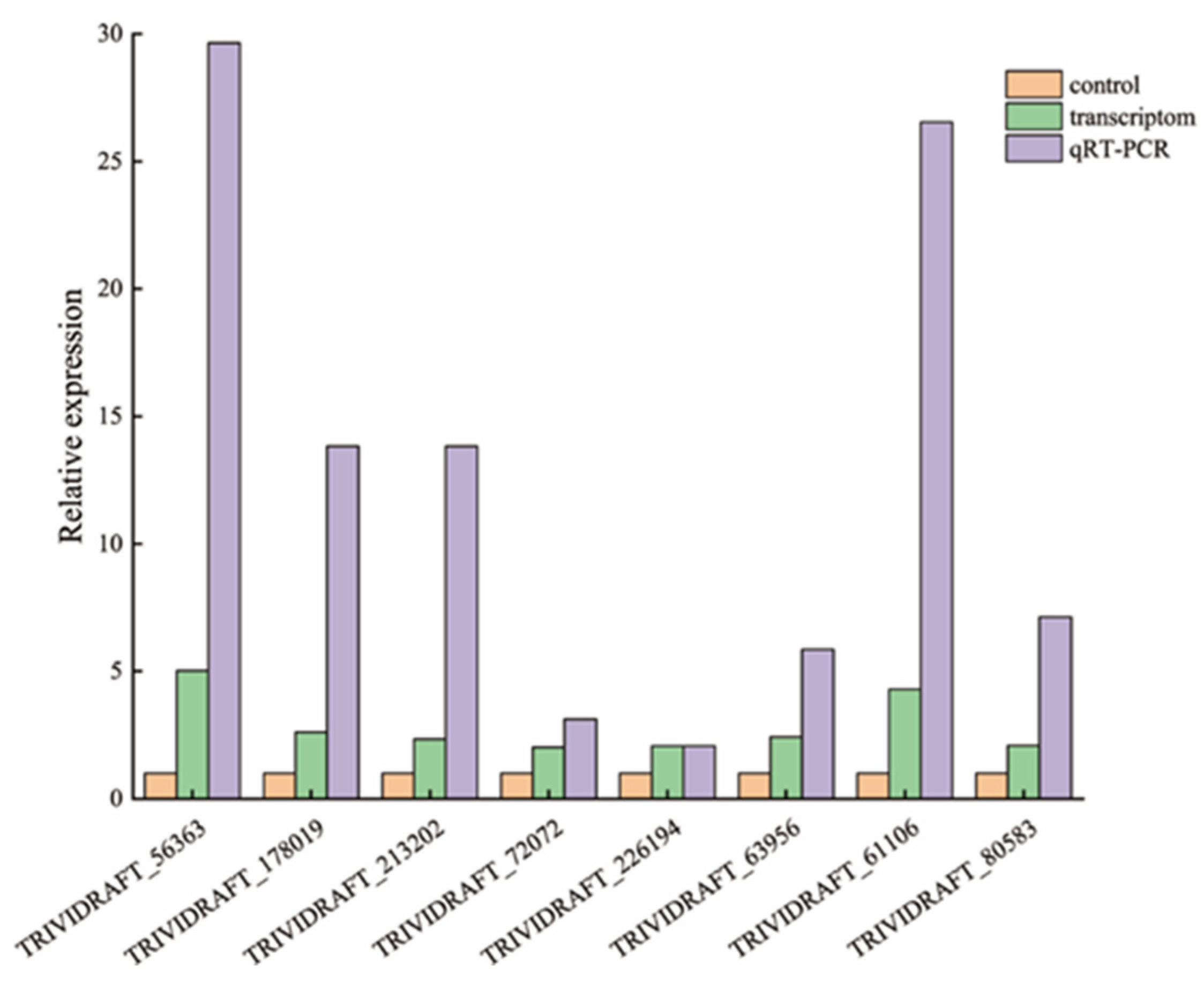
3.3.4. qRT-PCR Quantitative Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Halifu, S.; Deng, X.; Song, X.S.; Song, R.Q. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef] [Green Version]
- Kacprzak, M.; Asiegbu, F.O.; Daniel, G.; Stenlid, J.; ManKa, M.; Johansson, M. Resistance reaction of conifer species (european larch, norway spruce, scots pine) to infection by selected necrotrophic damping-off pathogens. Eur. J. Plant Pathol. 2001, 107, 191–207. [Google Scholar] [CrossRef]
- Martín-Pinto, P.; Pajares, J.; Díez, J. In vitro effects of four ectomycorrhizal fungi, Boletus edulis, Rhizopogon roseolus, Laccaria laccataand Lactarius deliciosuson Fusarium damping off in Pinus nigra seedlings. New For. 2006, 32, 323–334. [Google Scholar] [CrossRef]
- John, C.J.; Jishma, P.; Karthika, N.R.; Nidheesh, K.S.; Ray, G.J.; Mathew, J.; Radhakrishnan, K.E. Pseudomonas fluorescens R68 assisted enhancement in growth and fertilizer utilization of Amaranthus tricolor (L.). Biotech 2017, 7, 256. [Google Scholar]
- Fengge, Z.; Yunqian, H.; Cobb, A.B.; Gongwen, L.; Jiqiong, Z.; Gaowen, Y.; Zhang, J.Q.; Yang, G.W.; Gai, W.T.W.; Zhang, Y.J. Trichoderma biofertilizer links to altered soil chemistry, altered microbial communities, and improved grassland biomass. Front. Microbiol. 2018, 9, 848. [Google Scholar]
- Nunzio, F.; Valeria, V.; Woo, S.L.; Pepe, O.; Rosa, A.D.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-Based Biostimulants Modulate Rhizosphere Microbial Populations and Improve N Uptake Efficiency, Yield, and Nutritional Quality of Leafy Vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. Biocontrol 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Migheli, Q.; Steinberg, C. Microbiological control of soil-borne phytopathogenic fungi with special emphasis on wilt-inducing Fusarium oxysporum. New Phytol. 2009, 184, 529–544. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; He, J.Z.; Wang, Q.; Shen, C.C.; Ge, Y. Ectomycorrhizal fungi inoculation alleviates simulated acid rain effects on soil ammonia oxidizers and denitrifiers in Masson pine forest. Environ. Microbiol. 2019, 21, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Zimand, G. Effect of Trichoderma harzianum on Botrytis cinerea Pathogenicity. Phytopathology 1996, 86, 1255–1260. [Google Scholar] [CrossRef]
- Harman, G.E. Myths and Dogmas of Biocontrol Changes in Perceptions Derived from Research on Trichoderma harzinum T-22. Plant Dis. 2000, 84, 377–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, G.R.; Dandurand, L.C. Ecological Complexity and the Success of Fungal Biological Control Agents. Adv. Agric. 2014, 2014, 542703. [Google Scholar] [CrossRef]
- Zapata-Sarmiento, D.H.; Palacios-Pala, E.F.; Rodríguez-Hernández, A.; Melchor, D.L.M.M.; Rodríguez-Monroy, M.; Sepúlveda-Jiménez, G. Trichoderma asperellum, a potential biological control agent of Stemphylium vesicarium, on onion (Allium cepa L.). Biol. Control 2019, 140, 104–105. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Morán-Diez, E.; Hermosa, R.; Ambrosino, P.; Cardoza, R.E.; Gutiérrez, S.; Lorito, M.; Monte, E. The ThPG1 Endopolygalacturonase Is Required for the Trichoderma harzianum –Plant Beneficial Interaction. Mol. Plant-Microbe Interact. 2009, 22, 1021–1031. [Google Scholar] [CrossRef] [Green Version]
- Rubio, M.B.; Pardal, A.J.; Cardoza, R.E.; Gutiérrez, S.; Monte, E.; Hermosa, R. Involvement of the Transcriptional Coactivator ThMBF1 in the Biocontrol Activity of Trichoderma harzianum. Front. Microbiol. 2017, 8, 2273. [Google Scholar] [CrossRef] [Green Version]
- Nawrocka, J.; Małolepsza, U.; Szymczak, K.; Szezech, M. Involvement of metabolic components, volatile compounds, PR proteins, and mechanical strengthening in multilayer protection of cucumber plants against Rhizoctonia solani activated by Trichoderma atroviride TRS25. Protoplasma 2018, 255, 359–373. [Google Scholar] [CrossRef] [Green Version]
- Tijerino, A.; Cardoza, R.E.; Moraga, J.; Malmierca, G.M.; Vicente, F.; Aleu, J.; Collado, I.G.; Gutiérrez, S.; Monte, E.; Hermosa, R. Overexpression of the trichodiene synthase gene tri5 increases trichodermin production and antimicrobial activity in Trichoderma brevicompactum. Fungal Genet. Biol. 2011, 48, 285–296. [Google Scholar] [CrossRef]
- Leylaie, S.; Zafari, D. Antiproliferative and Antimicrobial Activities of Secondary Metabolites and Phylogenetic Study of Endophytic Trichoderma Species from Vinca Plants. Front. Microbiol. 2018, 9, 1484. [Google Scholar] [CrossRef] [Green Version]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Antonio, C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Howell, C.R. Mechanisms Employed by Trichoderma Species in the Biological Control of Plant Diseases: The History and Evolution of Current Concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, W.A.; Mandawe, J.C.; Kenerley, C.M. Multifunctional fungal plant symbionts: New tools to enhance plant growth and productivity. New Phytol. 2011, 190, 806. [Google Scholar]
- Woo, S.L.; Lorito, M. Exploiting the interactions between fungal antagonists, pathogens and the plant for biocontrol. In Novel Biotechnologies for Biocontrol Agent Enhancement and Management; Vurro, M., Gressel, J., Eds.; Springer: Dordrecht, The Netherlands; Portici, NA, Italy, 2007. [Google Scholar]
- Sharma, P. Biocontrol genes from Trichoderma species: A review. Afr. J. Biotechnol. 2011, 10, 19898–19907. [Google Scholar]
- Suárez, M.B.; Vizcaíno, J.A.; Llobell, A.; Monte, E. Characterization of genes encoding novel peptidases in the biocontrol fungus Trichoderma harzianum CECT 2413 using the TrichoEST functional genomics approach. Curr. Genet. 2007, 51, 331–342. [Google Scholar] [CrossRef]
- Flores, A.; Chet, I.; Herrera-Estrella, A. Improved biocontrol activity of Trichoderma harzianum by over-expression of the proteinase-encoding gene prb1. Curr. Genet. 1997, 31, 30–37. [Google Scholar] [CrossRef]
- Cardoza, R.E.; Malmierca, M.G.; Gutiérrez, S. Overexpression of erg1 gene in Trichoderma harzianum CECT 2413: Effect on the induction of tomato defence-related genes. J. Appl. Microbiol. 2014, 117, 812–832. [Google Scholar] [CrossRef]
- Galarza, L.; Akagi, Y.; Takao, K.; Kim, C.S.; Maekawa, N.; Itail, A.; Peralta, E.; Santos, E.; Kodama, M. Characterization of Trichoderma species isolated in Ecuador and their antagonistic activities against phytopathogenic fungi from Ecuador and Japan. J. Gen. Plant Pathol. 2015, 81, 201–210. [Google Scholar] [CrossRef]
- Ichiro, M.; Sunhwa, K.; Yamamoto, Y.C.; Ajisaka, K.; Maruyama, J.I.; Nakajima, H.; Kitamoto, K. Cloning and Overexpression of β-N-Acetylglucosaminidase Encoding Genenag A from A spergillus oryzaeand Enzyme-catalyzed Synthesis of Human Milk Oligosaccharide. Biosci. Biotechnol. Biochem. 2014, 20, 646–650. [Google Scholar]
- Aires, R.D.S.; Steindorff, A.S.; Ramada, M.H.S.; Siqueira, S.J.L.D.; Ulhoa, C.J. Biochemical characterization of a 27 kDa 1,3-β-d-glucanase from Trichoderma asperellum induced by cell wall of Rhizoctonia solani. Carbohydr. Polym. 2012, 87, 1219–1223. [Google Scholar] [CrossRef] [Green Version]
- Olmedo-Monfil, V.; Mendoza-Mendoza, A.I.; Gómez, C.C.; Herrera-Estrella, A. Multiple environmental signals determine the transcriptional activation of the mycoparasitism related gene prb1 in Trichoderma atrovirid. Mol. Genet. Genom. 2002, 267, 703–712. [Google Scholar] [CrossRef]
- Reithner, B.; Brunner, K.; Schuhmacher, R.; Peissl, I.; Seidl, V.; Krska, R.; Zeilinger, S. The G protein α-subunit Tga1 of Trichoderma atroviride is involved in chitinase formation and differential production of antifungal metabolites. Fungal Genet. Biol. 2005, 42, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Reithner, B.; Schuhmacher, R.; Stoppacher, N.; Pucher, M.; Brunner, K.; Zeilinger, S. Signaling via the Trichoderma atroviride mitogen-activated protein kinase Tmk1 differentially affects mycoparasitism and plant protection. Fungal Genet. Biol. 2007, 44, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivasithamparam, K.; Ghisalberti, E.L. Secondary metabolism in Trichoderma and Gliocladium. In Trichoderma and Gliocladium; Kubicek, C.P., Harman, G.E., Eds.; Taylor & Francis: London, UK, 1998; Volume 1, pp. 139–191. [Google Scholar]
- Doan, T.L.; Hajji, M.E.; Rebuffat, S.; Rajesvari, M.R.; Bodo, B. Fluorescence studies of the interaction of trichorzianine A IIIc with model membranes. Biochim. Biophys. Acta 1986, 858, 1–5. [Google Scholar] [CrossRef]
- Bae, S.J.; Mohanta, T.K.; Chung, J.Y.; Ryu, M.J.; Park, G.; Shim, S.; Hong, S.B.; Seo, H.C.; Bae, D.W.; Bae, I.; et al. Trichoderma metabolites as Biological Control Agents against Phytophthora Pathogens. Biol. Control 2015, 92, 128–138. [Google Scholar] [CrossRef]
- Seidl, V.; Song, L.; Lindquist, E.; Gruber, S.; Koptchinskiy, A.; Sun, J.B.; Grigoriev, I.; Herrera-Estrella, A.; Baker, S.E.; Kubicek, C.P. Transcriptomic response of the mycoparasitic fungus Trichoderma atroviride to the presence of a fungal prey. BMC Genom. 2009, 10, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational research on Trichoderma: From ‘omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395. [Google Scholar] [CrossRef] [Green Version]
- Lorito, M.; Woo, S.L. Trichoderma: A Multi-Purpose Tool for Integrated Pest Management. In Principles of Plant-Microbe Interactions; Springer International Publishing: Portici, NA, Italy, 2015. [Google Scholar]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2010, 10, 57–63. [Google Scholar] [CrossRef]
- Clark, T.A. Genomewide Analysis of mRNA Processing in Yeast Using Splicing-Specific Microarrays. Science 2002, 296, 907–910. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Huber, W.; Granovskaia, M.; Toedling, J.; Palm, C.J.; Bofkin, L.; Jones, T.; Davis, R.W.; Steinmetz, L.M. A high-resolution map of transcription in the yeast genome. Proc. Natl. Acad. Sci. USA 2006, 103, 5320–5325. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Benedito, V.A.; Wang, M.; Wuray, J.D.; Zhao, P.X.; Tang, Y.H.; Udvardi, M.K. The Medicago truncatula gene expression atlas web server. BMC Bioinform. 2009, 10, 441. [Google Scholar] [CrossRef] [Green Version]
- Cloonan, N.; Forrest, A.R.R.; Kolle, G.; Gardiner, B.B.A.; Faulkner, G.J.; Brown, M.K.; Taylor, D.F.; Steptoe, A.L.; Wani, S.; Bethel, G. Stem cell transcriptome profiling via massive-scale mRNA sequencing. Nat. Methods 2008, 5, 613–619. [Google Scholar] [CrossRef]
- Morin, R.; Bainbridge, M.; Fejes, A.; Martin, H.; Krzywinski, M.; Pugh, T.J.; Mcdonald, H.; Varhol, R.; Jones, S.J.M.; Marra, M.A. Profiling the HeLa S3 transcriptome using randomly primed cDNA and massively parallel short-read sequencing. Biotechniques 2008, 4, 81–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagalakshmi, U.; Wang, Z.; Waern, K.; Shou, c.; Raha, D.; Gerstein, M.; Snyder, M. The Transcriptional Landscape of the Yeast Genome Defined by RNA Sequencing. Science 2008, 320, 1344–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, Y.; Huang, Y.Y.; Jia, Z.H.; Ge, W.; Zhang, L.; Zhao, Q.; Song, S.S.; Huang, Y.L. Whole RNA-sequencing and gene expression analysis of Trichoderma harzianum Tr-92 under chlamydospore-producing condition. Genes Genom. 2019, 41, 689–699. [Google Scholar]
- Laura, L.; Susan, P.M.; Rosa, E.C.; Daren, W.B.; Hye, S.K.; Nancy, J.A.; Robert, H.P.; Santiago, G. Effect of deletion of a trichothecene toxin regulatory gene on the secondary metabolism transcriptome of the saprotrophic fungus Trichoderma arundinaceum. Fungal Genet. Biol. 2018, 8, 26–49. [Google Scholar]
- Pradeep, K.D.; Crespo, A.; Wedin, M. Evolution of complex symbiotic relationships in a morphologically derived family of lichen-forming fungi. New Phytol. 2015, 208, 1217–1226. [Google Scholar]
- Boddy, L.; Hiscox, J. Fungal Ecology: Principles and Mechanisms of Colonization and Competition by Saprotrophic Fungi. Microbiol. Spectr. 2016, 4, 293–308. [Google Scholar]
- Terhonen, E.; Sipari, N.; Asiegbu, F.O. Inhibition of phytopathogens by fungal root endophytes of Norway spruce. Biol. Control 2016, 99, 53–63. [Google Scholar] [CrossRef]
- Yin, D.C.; Deng, X.; Ilan, C.H.E.T.; Song, R.Q. Inhibiting effect and mechanism of Trichoderma virens T43 on four major species of forest pathogen. Chin. J. Ecol. 2014, 33, 1911–1919. [Google Scholar]
- Deng, X.; Song, X.S.; Yin, D.C.; Song, R.Q.; Ma, X.Q.; Zhang, X. Study on Screening of High Efficient Trichoderma Strains and Its Biocontrol of Seedling Blight. J. Jilin Agric. Univ. 2013, 35, 282–287. [Google Scholar]
- Bell, D.K. In Vitro Antagonism of Trichoderma species Against Six Fungal Plant Pathogens. Phytopathology 1982, 72, 379–382. [Google Scholar] [CrossRef]
- Qu, Y.; Lian, B. Bioleaching of rare earth and radioactive elements from red mud using Penicillium tricolor RM-10. Bioresour. Technol. 2013, 136, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Bao, P.; Liu, A.; Wang, M.; Shen, L.; Yu, R.; Liu, Y.; Chen, M.; Li, J.K.; Wu, X.L.; et al. Bioleaching of low-grade waste printed circuit boards by mixed fungal culture and its community structure analysis. Resour. Conserv. Recycl. 2018, 136, 267–275. [Google Scholar] [CrossRef]
- Hu, L.F. Studies on antifungal actions of cuminum cyminum L. In Seed; Northwest A&F University: Yangling, China, 2008. [Google Scholar]
- Deng, X.; Song, X.S.; Yin, D.C.; Song, R.Q. Water-based and dry powder formulation preparation of Trichoderma virens T43 and its field Application. For. Pest Dis. 2014, 33, 8–12. [Google Scholar]
- Dubey, S.C.; Suresh, M.; Singh, B. Evaluation of Trichoderma species against Fusarium oxysporum f. sp. ciceris for integrated management of chickpea wilt. Biol. Control 2007, 40, 118–127. [Google Scholar] [CrossRef]
- Iquee, S.; Cheong, B.E.; Taslima, K.; Kausar, H.; Hansan, M.M. Separation and Identification of Volatile Compounds from Liquid Cultures of Trichoderma harzianum by GC-MS using Three Different Capillary Columns. J. Chromatogr. Sci. 2012, 50, 358–367. [Google Scholar]
- Liu, S.; Lv, Z.; Liu, Y.; Li, L.; Zhang, l.d. Network analysis of ABA-dependent and ABA-independent drought responsive genes in Arabidopsis thaliana. Genet. Mol. Biol. 2018, 41, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Bashir, W.; Anwar, S.; Zhao, Q.; Hussain, I.; Xie, F. Interactive effect of drought and cadmium stress on soybean root morphology and gene expression. Ecotoxicol. Environ. Saf. 2019, 175, 90–101. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.M.; Mazutis, L.; Akartuna, I.; Tallapragada, N.; Veres, A.; Li, V.; Kirschner, M.W. Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells. Cell 2015, 161, 1187–1201. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.; Wu, Y.F. An SVM-based approach for discovering splicing junctions with RNA-Seq. In Proceedings of the IEEE International Conference on Computational Advances in Bio and Medical Sciences (ICCABS), Miami, FL, USA, 2–4 June 2014. [Google Scholar]
- Dewey, C.N.; Bo, L. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar]
- Wang, X.; Cairns, M.J. SeqGSEA: A Bioconductor package for gene set enrichment analysis of RNA-Seq data integrating differential expression and splicing. Bioinformatics 2014, 30, 1777–1779. [Google Scholar] [CrossRef] [Green Version]
- Velmourougane, K.; Prasanna, R.; Supriya, P.; Ramakrishnan, B.; Thapa, S.; Saxena, A.K. Transcriptome profiling provides insights into regulatory factors involved in Trichoderma viride-Azotobacter chroococcum biofilm formation. Microbiol. Res. 2019, 227, 126–292. [Google Scholar] [CrossRef] [PubMed]
- Durechova, D.; Jopcik, M.; Rajninec, M.; Jana, M.; Jana, L. Expression of Drosera rotundifolia Chitinase in Transgenic Tobacco Plants Enhanced Their Antifungal Potential. Mol. Biotechnol. 2019, 61, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2 -(∆∆Ct) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Harwoko, H.; Daletos, G.; Stuhldreier, F.; Lee, J.; Wesselborg, S.; Feldbrügge, M.; Müller, W.E.G.; Kalscheuer, R.; Ancheeva, E.; Proksch, P. Dithiodiketopiperazine derivatives from endophytic fungi Trichoderma harzianum and Epicoccum nigrum. Nat. Prod. Res. 2019, 3, 1–9. [Google Scholar] [CrossRef]
- Mukherjee, M.; Mukherjee, P.K.; Horwitz, B.A.; Zachow, C.; Berg, G.; ZEILINGER, S. Trichoderma–Plant–Pathogen Interactions: Advances in Genetics of Biological Control. Indian J. Microbiol. 2012, 52, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Li, J.S.; Wu, Y.Z.; Chen, K.; Wang, Y.L.; Hu, J.D.; Wei, Y.L.; Yang, H.T. Trichoderma cyanodichotomus sp. nov. a new soil-inhabiting species with a potential for biological control. Can. J. Microbiol. 2018, 64, 1020–1029. [Google Scholar] [CrossRef]
- Macías-Rodríguez, L.; Guzmán-Gómez, A.; García-Juárez, P.; Contreras-Cornejo, H.A. Trichoderma atroviride promotes tomato development and alters the root exudation of carbohydrates, which stimulates fungal growth and the biocontrol of the phytopathogen Phytophthora cinnamomi in a tripartite interaction system. FEMS Microbiol. Ecol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Kredics, L.; Liqiong, C.; Orsolya, K.; Rita, B.; Lóránt, H.; Henrietta, A.; Viktor, N.; Khaled, J.M.; Naiyf, S.A.; Alharbi, N.S.; et al. Molecular Tools for Monitoring Trichoderma in Agricultural Environments. Front. Microbiol. 2018, 9, 1599. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Salwan, R.; Sharma, P.N. Differential Response of Extracellular Proteases of Trichoderma Harzianum against Fungal Phytopathogens. Curr. Microbiol. 2016, 73, 419–425. [Google Scholar] [CrossRef]
- Stappler, E.; Christoph, D.; Tisch, D.; Monika, S. Analysis of Light- and Carbon-Specific Transcriptomes Implicates a Class of G-Protein-Coupled Receptors in Cellulose Sensing. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francesca, B.; Silvia, C.; Franzetti, E.; Congiu, T.; Marinelli, F.; Casartelli, M.; Tettamanti, G. Effects of Trichoderma viride chitinases on the peritrophic matrix of Lepidoptera. Pest Manag. Sci. 2016, 72, 980–989. [Google Scholar]
- Xiong, H.; Xue, K.; Qin, W.; Chen, X.; Wang, H.F.; Shi, X.H.; Ma, T.; Sun, Z.H.; Chen, W.G.; Tian, X.Q.; et al. Does Soil Treated with Conidial Formulations of Trichoderma spp. Attract or Repel Subterranean Termites. J. Econ. Entomol. 2018, 111, 808–816. [Google Scholar] [CrossRef]
- Ravindra, B.; Mala, M.; Benjamin, A.H.; Prasun, K.M. Regulation of conidiation and antagonistic properties of the soil-borne plant beneficial fungus Trichoderma virens by a novel proline-, glycine-, tyrosine-rich protein and a GPI-anchored cell wall protein. Curr. Genet. 2019, 65, 953–964. [Google Scholar]
- Dautt-Castro, M.; Estrada-Rivera, M.; Olguin-Martínez, I.; Rocha-Medina, M.D.C.; Islas-Osuna, M.A.; Casas-Flores, S. TBRG-1 a Ras-like protein in Trichoderma virens involved in conidiation, development, secondary metabolism, mycoparasitism, and biocontrol unveils a new family of Ras-GTPases. Fungal Genet. Biol. 2019, 136, 103292. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Wang, M.H. Isolation and molecular identification of Trichoderma species from wetland soil and their antagonistic activity against phytopathogens. Physiol. Mol. Plant Pathol. 2020, 109, 101458. [Google Scholar] [CrossRef]
- Ngikoh, B.; Karim, N.A.A.; Jahim, J. Characterisation of Cellulases and Xylanase from Trichoderma virens UKM1 and its Potential in Oil Palm Empty Fruit Bunch (OPEFB) Saccharification. J. Phys. Ence 2017, 28, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Kovalchuk, A.; Driessen, A.J.M. Phylogenetic analysis of fungal ABC transporters. BMC Genom. 2010, 11, 177. [Google Scholar] [CrossRef] [Green Version]
- Lanzuise, S.; Ruocco, M.; Scala, V.; Woo, S. Cloning of ABC transporter-encoding genes in Trichoderma spp. to determine their involvement in biocontrol. J. Plant Pathol. 2002, 84, 184. [Google Scholar]
- Ruocco, M.; Lanzuise, S.; Vinale, F.; Marra, R.; Turrà, D.; Woo, S.L.; Lorito, M. Identification of a New Biocontrol Gene in Trichoderma atroviride: The Role of an ABC Transporter Membrane Pump in the Interaction with Different Plant-Pathogenic Fungi. Mol. Plant Microbe Interact. 2009, 22, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.; Shahid, M. Trichoderma Genome to Genomics: A Review. J. Data Min. Genom. Proteom. 2014, 5, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Montero-Barrientos, M.; Cardoza, R.E.; Santiago, G.; Lorito, S.; Monte, E.; Hermosa, R. The heterologous overexpression of hsp23, a small heat-shock protein gene from Trichoderma virens, confers thermotolerance to T. harzianum. Curr. Genet. 2007, 52, 45–53. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Sequence (5′–3′) |
|---|---|---|
| TRIVIDRAFT_56363 | 1-TRIVIDRAFT_56363-F 1-TRIVIDRAFT_56363-R | TATATGAGCGCTGCTAAGAT AAATTGGGACTTGTGAGTGT |
| TRIVIDRAFT_178019 | 2-TRIVIDRAFT_178019-3F 2-TRIVIDRAFT_178019-3R | GGATCCAGATTCAGTTCTAT GGATCCAGATTCAGTTCTAT |
| TRIVIDRAFT_213202 | 3-TRIVIDRAFT_213202-3F 3-TRIVIDRAFT_213202-3R | GGATCCAGATTCAGTTCTAT GGATCCAGATTCAGTTCTAT |
| TRIVIDRAFT_72072 | 4-TRIVIDRAFT_72072-F 4-TRIVIDRAFT_72072-R | GGATCCAGATTCAGTTCTAT GGATCCAGATTCAGTTCTAT |
| TRIVIDRAFT_226194 | 5-TRIVIDRAFT_226194-2F 5-TRIVIDRAFT_226194-2R | GGATCCAGATTCAGTTCTAT CAGAAATCTGCATTTGCAAG |
| TRIVIDRAFT_63956 | 6-TRIVIDRAFT_63956-F 6-TRIVIDRAFT_63956-R | CAGAAATCTGCATTTGCAAG CGTGCTTGCGATGTGTAAGT |
| TRIVIDRAFT_61106 | 7-TRIVIDRAFT_61106-F 7-TRIVIDRAFT_61106-R | CTCGGACAACAGCCAGTTTC CTCGGACAACAGCCAGTTTC |
| TRIVIDRAFT_80583 | 8-TRIVIDRAFT_80583-F 8-TRIVIDRAFT_80583-R | CTCGGACAACAGCCAGTTTC CTCGGACAACAGCCAGTTTC |
| Sample | Raw Reads | Clean Reads | Error Rate (%) | Q30 (%) | GC Content (%) | Total Mapped |
|---|---|---|---|---|---|---|
| Tv | 47,012,424.00 | 46,570,488.67 (99.00%) | 0.02 | 95.91 | 54.17 | 43,264,365.67 (92.91%) |
| Rs-Tv | 45,402,895.33 | 44,737,395.33 (99.00%) | 0.02 | 95.50 | 52.66 | 26,959,925.00 (60.12%) |
| Total | 92,415,319.33 | 91,307,884.00 (99.00%) | 0.02 | 95.71 | 53.42 | 70,224,291.00 (76.52%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halifu, S.; Deng, X.; Song, X.; Song, R.; Liang, X. Inhibitory Mechanism of Trichoderma virens ZT05 on Rhizoctonia solani. Plants 2020, 9, 912. https://doi.org/10.3390/plants9070912
Halifu S, Deng X, Song X, Song R, Liang X. Inhibitory Mechanism of Trichoderma virens ZT05 on Rhizoctonia solani. Plants. 2020; 9(7):912. https://doi.org/10.3390/plants9070912
Chicago/Turabian StyleHalifu, Saiyaremu, Xun Deng, Xiaoshuang Song, Ruiqing Song, and Xu Liang. 2020. "Inhibitory Mechanism of Trichoderma virens ZT05 on Rhizoctonia solani" Plants 9, no. 7: 912. https://doi.org/10.3390/plants9070912