1. Introduction
Plant endophytes widely exist in plants [
1]. Dark septate endophytes (DSE) represent a major group on endophytes within plants characterized by a dark mycelium color and distinct septum. They colonize the epidermis, cortex, and even the intercellular space of vascular tissue of healthy plant roots to form symbionts without causing plant diseases [
2,
3]. The host range of dark septate endophytic fungi covers nearly 600 species of plants from 114 families and 320 genera. DSE colonization has been found in mycorrhizal plants as well as roots of traditional non-mycorrhizal plants such as Cyperaceae, Cruciferae, and Chenopodiaceae [
4]. In conifers, the main DSE fungi are the group of the
Phialocephala fortinii s.l.—
acephala applanata species complex (PAC) belonging to the ascomycetes. PACs can be found in the Northern Hemisphere from polar to tropical areas and play a leading role in the root system of conifers [
5,
6]. Several studies have reported that DSE fungi exhibit a positive effect on plant growth [
7,
8,
9,
10]. They enhance the mineralization process of insoluble phosphorus in soil [
11,
12], promote the uptake and utilization of nutrients like nitrogen (N) or phosphorus (P) by the plants [
13] and increase the stress tolerance of the host plant [
14,
15,
16,
17]. In addition, DSE fungi can colonize the host roots and effectively inhibit the occurrence of soil-borne diseases [
18,
19,
20,
21].
Mongolian pine (
P. sylvestris var.
mongolica) is a geographical variety of the Scots pine (
P. sylvestris). Its natural habitats are the Daxinganling mountains in China and parts of Russia and Mongolia [
22]. Due to its advantages of rapid growth, cold tolerance, drought resistance, and potent adaptability [
23], it is now the major coniferous tree species used for the “Sand-Control Project” in China, which has a vital role in environmental restoration and ecological conservation. Damping-off is the most widespread and destructive disease that affects young conifer seedlings in forest nurseries around the world [
24] Several soil-borne pathogens such as different species of
Pythium,
Fusarium, and
Rhizoctonia coexist in most nursery soils and are responsible for severe damage in nurseries [
25]. Damping-off damage caused by
Rhizoctonia, particularly
R. solani, is widely reported in forest nurseries around the world [
26,
27,
28]. Like other conifer seedlings, the soil-borne diseases of
P. sylvestris var.
Mongolica are managed in the nursery by chemical control. However, pesticides and chemical fertilizers have often been excessively applied, which leads to a variety of negative impacts, like critical illness, damage to the soil environment, or retarded seedling growth [
29]. At present, the use of microbial metabolites produced by beneficial microbiota is a novel, environmentally friendly approach to manage plant health in comparison with the use of chemical pesticides. It is advantageous in that it is free of pollution or residues, it shows pathogen specificity, difficulty in resistance generation and has human or animal health benefits, while providing environmental protection [
30].
P. fortinii s.l.—
acephala applanata species complex (PAC) members have been noted to suppress pathogens [
31].
P. sphareoides could effectively inhibit
Heterobasidion parviporum infection of Norway spruce by producing a variety of bioactive substances, as well as reduce mortality and the disease intensity [
32]. PAC strains with low toxicity were also verified by inoculation experiments to control high toxicity PAC pathogens using niche competition [
33]. Christoph [
34] isolated 85 PAC strains from Norway spruce (
Picea abies) roots, among which
P. europaea dramatically decreased
Phytophthora citricola growth in vitro.
P. bamuru A024 (isolated from the root of
P. sylvestris var.
Mongolica collected on the Jiagedaqi Experimental Forest Farm in Hei Longjiang Province, China) is one of the PAC members that has a strong ability to inhibit the growth of
R. solani in vitro (unpublished data). In this study,
P. bamuru A024 was utilized for investigating the impacts of DSEs on damping-off biocontrol in vivo, root structure and growth promotion for the
P. sylvestris var.
Mongolica annual seedlings, rhizosphere soil physicochemical characters, as well as the microbial community structure. Thus, our study aimed to assess the impacts of
P. bamuru A024 on: (1) Seedling damping-off disease of
P. sylvestris var.
mongolica; (2) growth promotion as well as root structure in the seedlings in
P. sylvestris var.
mongolica; (3) rhizosphere soil physical characteristics together with enzymatic activities of the seedlings in
P. sylvestris var.
mongolica; (4) rhizosphere microbial structure in the seedlings of
P. sylvestris var.
mongolica.
2. Materials and Methods
2.1. Organisms and Growing Environment
Dark septate endophytes strain, P. bamuru A024 (NCBI accession number: MN006137), was isolated from the root of P. sylvestris var. Mongolica, collected at the Jiagedaqi Experimental Forest Farm in Hei Longjiang Province of China. The pathogenic fungus, R. solani SH01, was isolated from roots of diseased conifer seedlings, collected at the Weihe seedling nursery of Hei Longjiang Province, China. The above two strains were then inoculated on potato dextrose agar (PDA) medium supplemented with 12 g/L potato extract, 20 g/L dextrose and 14 g/L agar (pH 6.0, Haibo Biotechnology, Qingdao, China).
After 20 days of growth on PDA medium,
P. bamuru A024 mycelium was collected using a 5 mm sterile puncher. The mycelia disks were transferred to solid cottonseed shell medium [
35] (cottonseed shell 200 g, glucose 2 g, MgSO
4·7H
2O 3 g, KH
2PO
4 3 g, (NH
4)
2HPO
4 3 g, Ca
3(PO
4)
2 2 g, vitamin B
1 1 mg, 60% water content, and steam-sterilized at 0.1 MPa at 121 °C for 1 h), followed by 30 days of incubation at 25 °C in the dark.
The pathogenic fungi,
R. solani SH01, after five days of growth on the PDA medium, was sampled using the 5 mm sterile puncher. A suspension of
R. solani SH01 was prepared by placing the mycelium inoculant in liquid PD medium (agar-free PDA medium) and incubating for 7 days under 25 °C in dark with 150 rpm of agitation. The culture suspension was mixed with sterile culture substrate (vermiculite:river sand:peat soil = 1:1:3, v:v:v) in a ratio of 1:10 (v:v), and subsequently used for seedling inoculation [
35].
Potassium permanganate (0.5% v/v) was used to sterilize the surface of P. sylvestris var. mongolica experimental seeds (Zhanggutai Experimental Forest Farm, Zhangwu County, Liaoning Province, China) for 30 min, later, distilled water was used to rinse the seeds five times. Thereafter, seeds were further subjected to 5 days of germination on sterilized wet gauze at 25 °C.
A mixture consisting of peat soil, sand and vermiculite in a volume ratio of 2:1:1 was used as the substrate for culturing. First it was disinfected for 2 h in a high-temperature autoclave at 121 °C, and after cooling, it was transferred into plastic pots (15 cm × 13 cm). Each pot was first filled with 600 g culture substrate followed by 100 g of solid inoculum (for the control treatment, the same amount of sterile cottonseed shell culture medium was used), ensuring the coverage using the culture substrate was 1 cm in thickness. Germinated seeds (30 seeds per pot) were covered with 2 cm thick culture substrate, and then the substrate was watered and grown in the dark. After the seedlings emerged from the soil they were placed in a greenhouse at 10 plants per pot and 20 pots per treatment. Each treatment was divided into two groups and randomly subjected to the different treatments. The plants were watered once every two days, and Hoagland nutrient solution was poured on them once a week [
23].
2.2. Design of Experiments and Inoculation of Seedlings
In each treatment (also for control), 20 pots (about 10 seedlings/pot) were selected so altogether 200 seedlings were prepared in each treatment. The four treatments included: (1) inoculation with sterile cottonseed shell medium (CK); (2) P. bamuru A024 (DSE) inoculation alone; (3) R. solani SH01 (CK + SH) inoculation alone; and (4) inoculation with P. bamuru A024 and R. solani SH01 (DSE + SH). One month after sowing, the seedlings of P. sylvestris var. mongolica were incubated using the pathogen (50 mL per pot), spread flat around to the seedlings. Each treatment was randomly assigned in the greenhouse environment, as mentioned previously.
2.3. Damping-Off Control Together with Physiological Index Determination
Damping-off rate in seedlings was investigated after 45 days of inoculation with R. solani SH01, and the relative control effect was also calculated. The survival rate of seedlings was counted three months after sowing. About 100 seedlings were investigated per treatment.
Forty five days after inoculation with R. solani SH01, 40 P. sylvestris var. mongolica seedlings were collected for the physiological index measurements. The roots of seedlings were rinsed using sterile water, then sterile filter paper was used to absorb excess water. The seedlings were further cut into 1 cm pieces, ground in liquid nitrogen and transferred to a 5 mL centrifuge tube. An acid ninhydrin colorimetry approach was applied to measure proline. The thiobarbituric acid chromogenic method was applied to measure malonaldehyde (MDA) content. Anthrone colorimetry was applied in measuring soluble sugar content. The Coomassie brilliant blue G-250 staining method was applied to measure soluble protein content. The nitroblue tetrazolium colorimetry approach was applied to measure superoxide dismutase (SOD) activity. The guaiacol method was applied to measure peroxidase (POD) activity. A hydrogen peroxide ultraviolet absorption method was applied to measure catalase (CAT) activity. The 3,5-dinitrosalicylic acid (DNS) method was applied to measure chitinase activity. The reducing sugar colorimetry method was applied to measure β-1,3-glucanase activity. The deamination of phenylalanine method was applied to measure phenylalanine ammonia lyase (PAL) activity. All the above indices were determined with detection kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.4. Sample Collection and Seedling Analysis
At 3 months after sowing, all seedlings were collected, and soil was carefully removed by washing so as to not damage the root system. Altogether 100 seedlings were randomized for every treatment, among which the initial 30 seedlings were adopted for measuring biomass indexes. In every seedling, the biomass indexes included plant height, dry weight, fresh weight upon collection, and ground level diameter. After measuring fresh weight, seedlings were dried in an oven for 5 h under 85 °C to measure their dry weight [
36].
When sampling, after washing the root system to remove the soil trying to not damage the roots system of seedlings, 10 seedlings were randomly selected and an Epson v700 root scanner (Seiko Epson Corporation, Nagano, Japan) was used to scan and grade the root system of the seedlings. The indexes of mean diameter, root tip number, bifurcation number, root volume, root length and surface area were thus analyzed.
2.5. Soil Character Analysis
Soil was sampled from the place where the 100 seedlings were harvested for each treatment. Rhizosphere soil samples were obtained within the root zone at a depth of 5 mm by using a brush and then filtered by a 1-mm mesh sieve. Thereafter, those soil samples adopted for measuring enzymatic activities and determining physicochemical characters were dried in the air at 25 °C, packaged in the sterile sample bags, and finally preserved at 5 °C for later analyses.
The Kjeldahl approach was applied in measuring total nitrogen (TN) and the alkaline hydrolysis diffusion assay was conducted to determine the available nitrogen (AN). After sulfuric acid digestion, total phosphorus (TP) was determined by the anti-colorimetry of MO-SB method and total potassium (TK) was determined by flame photometry. The sulfuric acid-hydrochloric acid (double acid) extraction and anti-colorimetry of MO-SB method was applied to measure available phosphorus (AP). NH
4OAc leaching and flame photometry was used to determine available potassium (AK). In addition, the potassium dichromate oxidation-external heating approach was employed for measuring organic matter (OM). The soil pH (1:2.5) was determined by using a pH meter [
37]. The sodium phenol colorimetry approach was employed to measure urease activity. The glucose colorimetry approach was employed to measure saccharase activity. The disodium phenylphosphate colorimetry approach was employed to measure acid phosphatase activity. The hydrogen peroxide ultraviolet absorption method was applied to measure catalase activity. These four soil enzyme activities were determined using detection kits (Nanjing Jiancheng Bioengineering Institute).
2.6. Analysis of Bacterial Diversities
Rhizosphere soils were sampled according to the abovementioned method, then a 5.0 g sample was prepared for each biological replicate, which was put into a 50 mL sterilized centrifuge tube before it was sent to our laboratory in a cooler containing ice bags. In addition, soil that was utilized in high-throughput sequencing was preserved in a centrifuge tube at –80°C for further assessment.
Then, total genome DNA was extracted from the 0.5 g soil sample using the EZNA Soil DNA Kit (Omega Bio-Tek, Norcross, GA, USA) in accordance with manufacturer’s protocol. The sample was then subjected to high-throughput sequencing for soil microbiota. Thereafter, 100 µL elution solution was obtained from the literature to elute the obtained DNA. The DNA quality (A260/A280) and content were determined using a NanoDrop2000 spectrophotometer (Thermo Scientific, Waltham, MA, USA). All treatments were repeated three times. Meanwhile, the 16S rRNA region was subjected to high-throughput sequencing for determining the bacterial community in the soil. The V3 + V4 region in the 16S rRNA gene of bacteria was amplified using the universal 338F (5′-ACTCCTACGGAGGCAGCAG-3′) and 806R (5′-GGACTA CHVGGGTWTCTAAT-3′) primers.
The reaction system prepared for PCr consisted of 5×FastPfu buffer (4 µL), each primer (0.8 µL, 5 µM), 2.5 mM dNTPs (2 µL), bovine serum albumin (BSA, 0.2 µL), template DNA (10 ng), FastPfu polymerase (0.4 µL) together with double-distilled water (11.6 µL) in a total volume of 20 µL. Moreover, PCR was conducted by the following conditions: 3 min at 95 °C; 30 s at 95 °C, 30 s at 50 °C, 30 s at 72 °C for 27 cycles, followed by 10 min of extension at 72 °C. Later, the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA) was used to purify the resultant products, whereas QuantiFluor-ST (Promega, Madison, WI, USA) was used for quantification. Later, those amplicons that were purified were mixed at the equimolar dose to form a single aliquot, which was used later to construct the library. An Illumina MiSeq sequencer (Majorbio Biotechnology Co., Ltd., Shanghai, China) was applied in performing sequencing. The raw fastqfu files were filtered and merged using Trimmomatic and FLASH (version1.2.11) [
38,
39], whereas pyrosequencing data were analyzed using UPARSE (version 7.1,
http://drive5.com/uparse/). Subsequently, all sequences were classified as operational taxonomic units (OTUs) at the similarity threshold of 97%, while UCHIME (version 4.1) was used to remove chimeras [
40]. For all sequences, their taxonomic annotations were analyzed using RDP Classifier (
http://rdp.cme.msu.edu/) at the 0.7 confidence threshold.
2.7. Data Analyses
WPS 2016 (Kingsoft Corporation, Beijing, China) was adopted to process data. Differences in index of root structure, biomass of plant, soil pH, soil enzymes and chemical characteristics were analyzed through one-way ANOVA (Tukey test) by adopting IBM SPSS 22.0 (IBM Corporation, New York, NY, USA). Moreover, correlation analysis was conducted by Pearson’s approach. A difference of p < 0.05 indicated statistical significance.
The Mothur software (The University of Michigan, Michigan, MI, USA) was used for analyzing α-diversity indexes as well as rarefaction. At the same time, the sequencing depth was represented by the coverage index. Moreover, both Ace and Chao1 indices were adopted for describing abundances of microbiota, whereas Shannon and Simpson indices were applied in representing species richness as well as microbial community diversity. Unifrac distance was calculated to compare the bacterial β-diversity. Differences in the phylum and genus relative abundance between four treatments were analyzed through one-way ANOVA (Tukey test) by adopting IBM SPSS 22.0. Environmental factor correlation was analyzed using RDA methods. The Origin 2019b (Origin Lab Corporation, Northampton, MA, USA) software was employed for figure generation.
4. Discussion
The plant rhizosphere is a complex micro-ecological system wherein the interaction between beneficial microorganisms and host plants can promote the growth and stress resistance of plants. Dark septate endophytes (DSEs) can colonize the cortex and epidermis spaces inside and outside cells, or even the vessel tissue of healthy plant roots, to form symbionts. DSEs play an important part in promoting plant development and enhancing nutrient absorption, along with stress resistance. Strain A024 isolated and screened from the roots of P. sylvestris var. Mongolica was found to effectively suppress R. solani in vitro (unpublished data). To further study the interaction mechanism between the strain A024 and P. sylvestris var. mongolica and its control effect on R. solani, we have mainly focused on: (1) The control effect of strain A024 on R. solani; (2) the effect on promoting P. sylvestris var. mongolica; (3) the impacts on microbial community structure in rhizosphere soil.
This study reported a damping-off rate of 28.15% and a survival rate of 80.15% for DSE + SH treatment. Whereas for CK + SH treatment, the incidence rate of damping-off was 72.15%, and the survival rate was 33.35%. However, the relative control effect of strain A024 treatment against damping-off was 60.98%. These results of the seedling inoculation test showed that
P. bamuru A024 could effectively control the occurrence of damping-off caused by
R. solani on
P. sylvestris var.
mongolica seedlings. These results are similar to those of other
Phialocephala studies. Christoph [
34] isolated 85 PAC strains from Norway spruce (
Picea abies) roots, among which
P. europaea dramatically decreased
Phytophthora citricola growth in vitro. The main inhibitory agents were identified as sclerotinin A and B, sclerolide, and sclerin, however, no seedling inoculation test was done. The result of the sterile seedlings inoculation experiment done by Terhonen indicated that
P. sphareoides could prevented Norway spruce seedling root infection by the pathogen,
Heterobasidion parviporum under
in-vitro conditions. These results indicated that DSE fungi could colonize the host root system and effectively inhibit the occurrence of soil-borne diseases. Beside this, DSE fungi can also enhance seedlings’ disease resistance against soil borne pathogens. In our research, under the stress of
R. solani, the β–1,3-glucanase and chitinase activity, together with the contents of proline and soluble protein in seedlings from DSE + SH treatment increased by 18.41%, 92.54%, 101.27% and 30.48% respectively in comparison to the CK + SH treatment (
Table 1). Su [
41] found that the DSE strain
Harpophora oryzae enhanced disease resistance in rice and reduced root blast disease caused by
Magnaporthe oryzae by up-regulation of expression of key genes of salicylic acid (SA) signaling pathway. Therefore, improving plant resistance is one of the main reasons to reduce the occurrence of soil-borne disease.
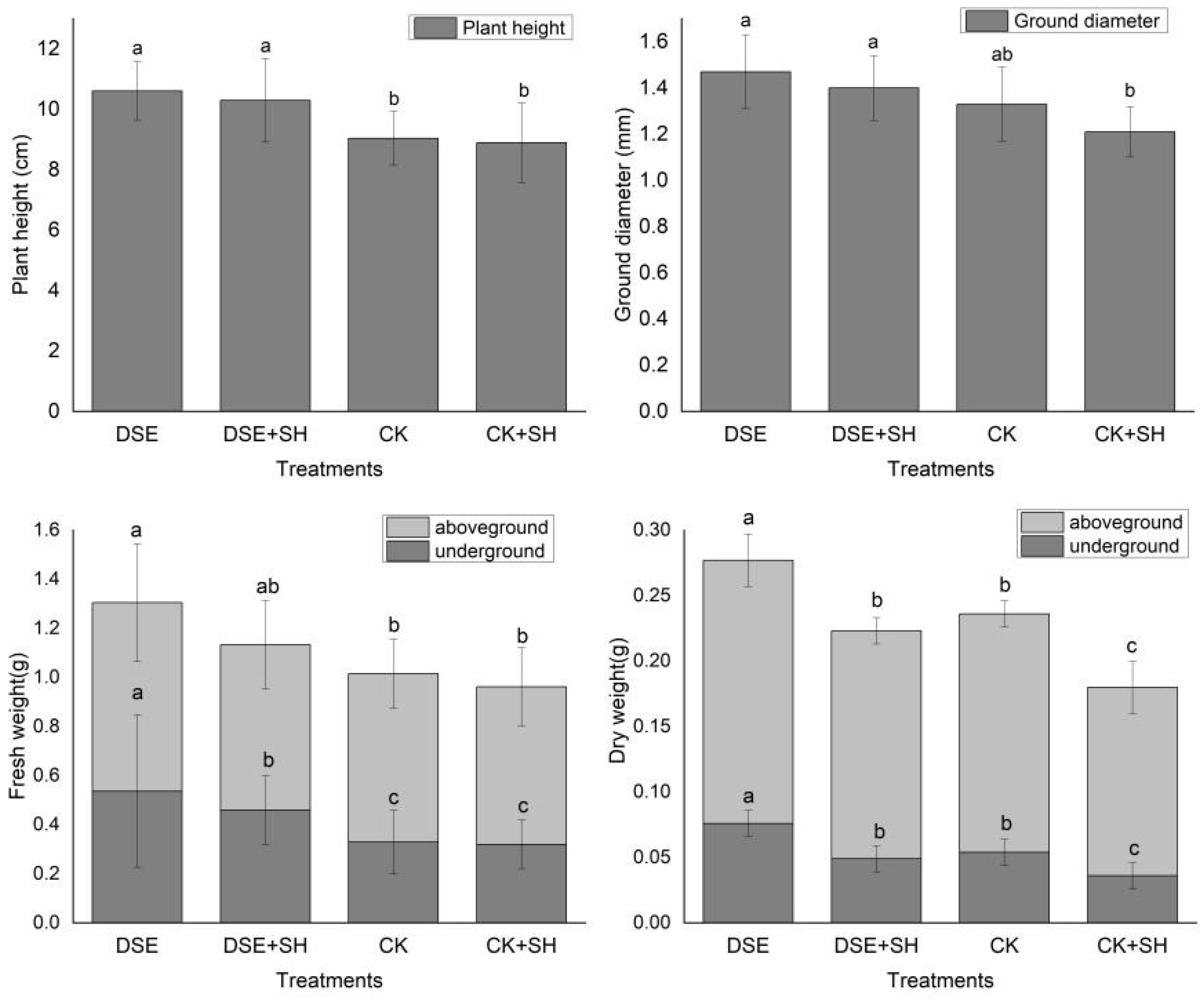
Dark septate endophytes (DSE) also has an effect on plant growth. The direct effects include promoted plant growth, an increase in biomass, as well as boosting the root structure and development of plants. In our research, dark septate endophytes strain A024 enhanced annual seedling growth, compared to CK treatment, underground and aboveground fresh weight, seedling height, aboveground and underground dry weight of seedlings from DSE treatment increased by 17.31%, 62.91%, 12.09%, 40.74%, 10.43%, respectively. Similarly, fresh underground weight, ground diameter, seedling height, underground and aboveground dry weights under DSE + SH treatment increased by 15.82%, 15.71%, 43.43%, 36.11%, 36.11%, and 20.83%, respectively, compared to CK + SH treatment (
Figure 1). These results indicated that DSE has an effect on plant growth even under the stress of
R. solani. Andrade [
42] inoculated tomato plants with
Leptodontidium orchidicola liquid inoculum, that increased the biomass by 20%, and number of fruits by two-fold, in comparison with control. The same result was found in an experiment that involved inoculation of
Harpophora radicicola with
Vulpia ciliate [
43]. DSE fungi can secrete a variety of extracellular hydrolases, such as amylase, pectinase, laccase, cellulase, lipase, protease, tyrosinase, polyphenol oxidase, and xylanase. Thus, the presence of a variety of hydrolases ensures the utilization of various nutrients [
44]. Dark septate root endophytic fungi,
Phialocephala fortinii, increased the growth of the seedlings for Scots pine in the presence of increased CO
2 content via enhancing the utilization efficiency of nitrogen [
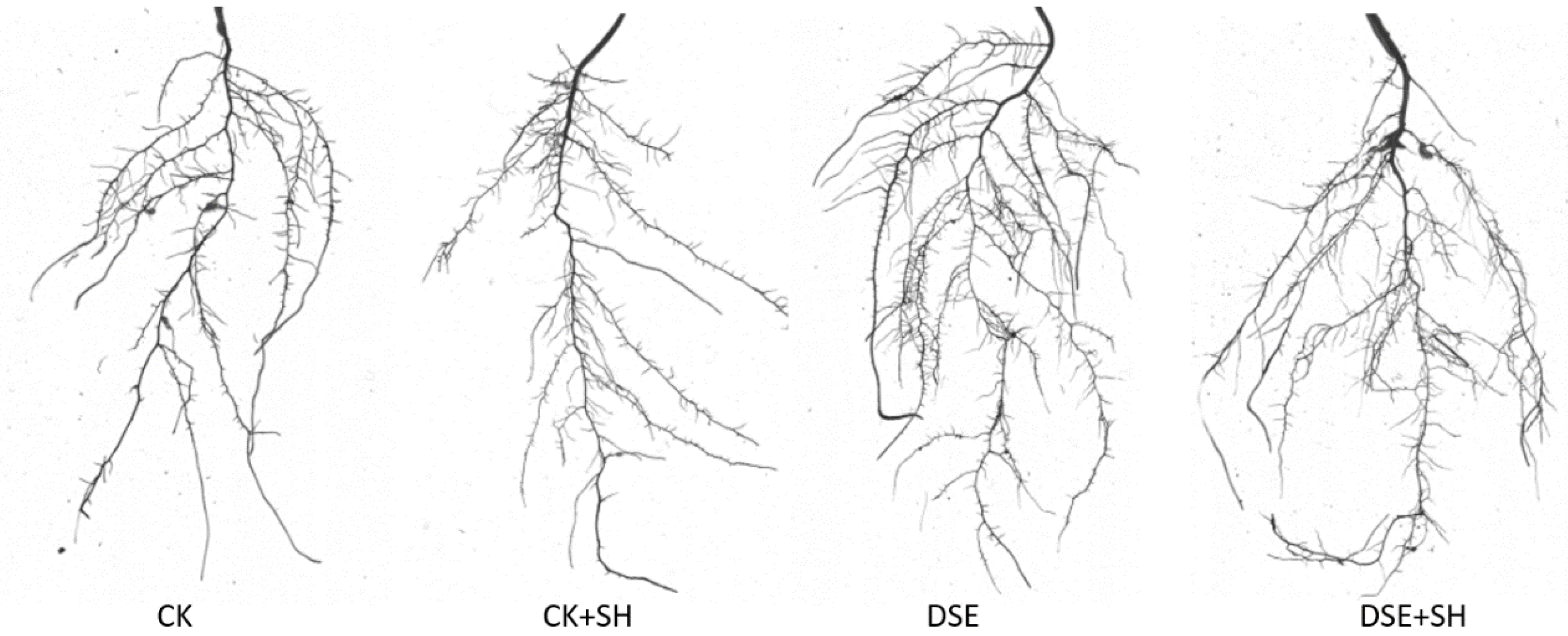
45]. Surface area and root length account for the vital factors to measure root distribution, meanwhile, root volume, average root diameter, fork number and tip number are also the vital factors to measure the absorption efficiency of root. Inoculation with DSEs can regulate the plant root architecture and facilitate the growth of root systems. In our research, DSE A024 significantly (
p < 0.05) enhanced root volume and length, fork number and root surface area in
P. sylvestris var.
mongolica in comparison with control treatment. Although, differences in the root architecture indexes were not significant between DSE and DSE + SH treatments, but CK and CK + SH treatments showed a significant difference (
Table 2 and
Figure 2). These results showed that DSE inoculation could counteract the negative effect of the pathogen on the root structure. The change of root structure promoted the growth of annual seedlings.These results are similar to other studies, Li [
46] inoculated
Leptosphaeria sp.,
knufia sp., and
Darksidea sp., with
A. mongolicus, and the plant root biomass and length remarkably elevated compared to control.
P. graminicola inoculation can also change the specific root length, root diameter, and root hair number of
Vulpia ciliata ssp.
Ambigua [
47].
A number of soil nutrient elements show low or no solubility, thus limiting soil nutrient circulation [
48,
49]. The existence of P and N within soil limits absorption and utilization efficiency of plants. Conversion of these organic nutrients to the inorganic ones to be easily taken up by plants depends on the microbes-derived extracellular enzymes, such as bacteria or fungi. DSE can improve the efficiency of plant uptake and utilization of soil nutrients. In our research, the seedlings of
P. sylvestris var.
mongolica, rhizosphere soil nutrient indices including AN, TP, AP, TK, AK increased by 57.57%, 32.53%, 103.84%, 33.11%, and 23.52%, respectively under the DSE treatment compared to the CK treatment (
Table 3). Soil enzymes play a vital part in conserving energy and circulating organic matter in soil.
P. bamuru A024 inoculation significantly elevated enzyme activities in seedling rhizosphere soil. In comparison to CK treatment, activities of catalase, urease, and acid phosphatase under DSE treatment increased by 12.97%, 20.93% and 19.67%, respectively (
Table 3). Therefore, DSE can improve the content of soil available K and P, thereby improving plant absorption efficiency while increasing the accumulation of biomass. In other research, Jumpponen et al. combining DSE inoculation with nitrogen fertilizer application increased the biomass accumulation of seedlings by 50% compared to the application of nitrogen fertilizer alone, which proved that the existence of DSE improved the utilization efficiency of the nitrogen fertilizer [
50]. The inoculation of
Atriplex canescens (Pursh) Nutt. with
Aspergillus ustus increased the P utilization efficiency, especially for the unavailable P [
51]. Spagnoletti measured the ability of nine DSE strains for solubilizing aluminum and iron phosphates, and DSE could solubilize insoluble phosphate [
52]. Knapp [
53] used Biolog FF together with API-ZYM tests for analyzing 15 DSE’s metabolic diversity. Differences were detected across diverse species, but each of our tested substrate was absorbed by DSE fungi. So the improvement of soil nutrients is one of important reasons to seedlings growth promotion.
According to our high-throughput sequencing analysis results, the rhizosphere bacterial community of
P. sylvestris var.
mongolica is dominated by Proteobacteria, with the highest relative abundance among the four treatments. However, the differences between the four treatments are not significant. The relative abundance of the Actinobacteria, Gemmatimonadetes were higher in CK and CK + SH treatments, whereas Bacteroidetes, Chloroflexi and Firmicutes were higher in DSE and DSE + SH treatments (
Table 5). The results of α-diversity index analysis showed that DSE had a significant effect on rhizosphere soil richness for
P. sylvestris var.
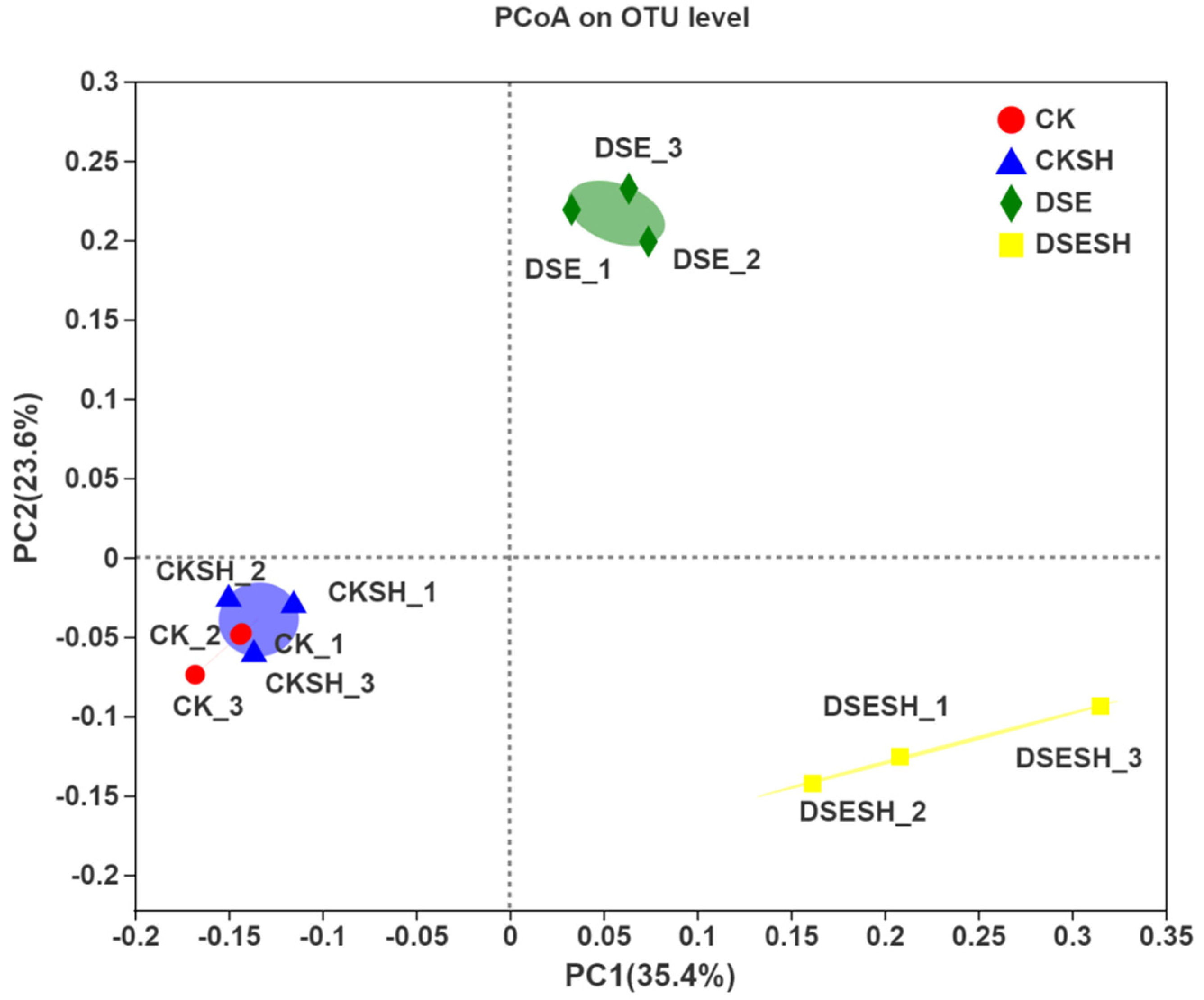
mongolica, but little effect on the diversity. Moreover, it was found through analyzing β-Diversity Indexes that, there were higher differences between bacteria groups under CK and CK + SH, DSE, and DSE + SH treatments than those inside each treatment (
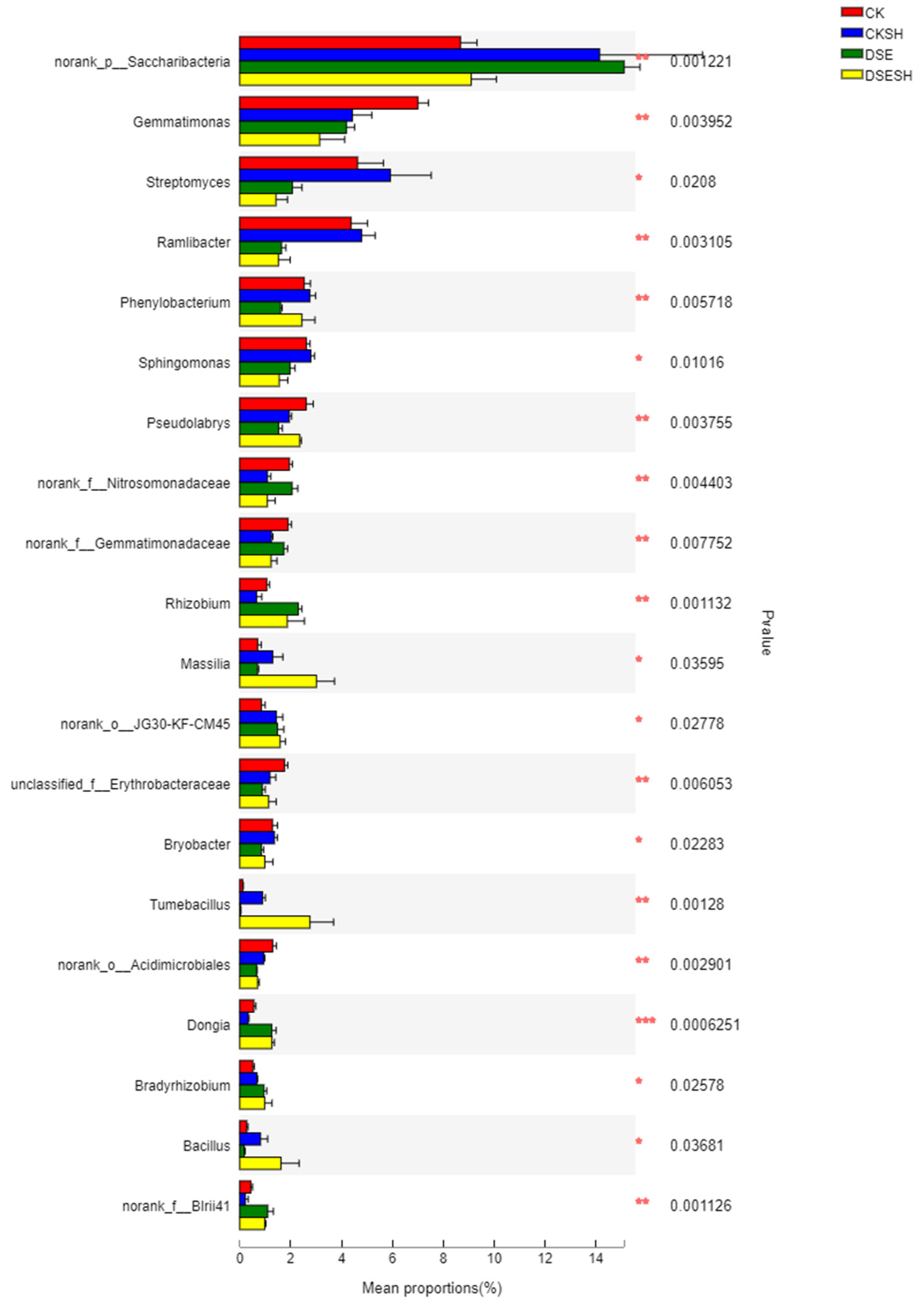
Figure 5). And some genus include
Massilia,
Rhizobium,
Bacillus,
Tumebacillus,
Dongia and
Bradyrhizobium were higher in DSE treatments, in contrast, the relative abundance of
Streptomyces,
Gemmatimonas,
Ramlibacter,
Sphingomonas and
Bryobacter were higher in CK treatments (
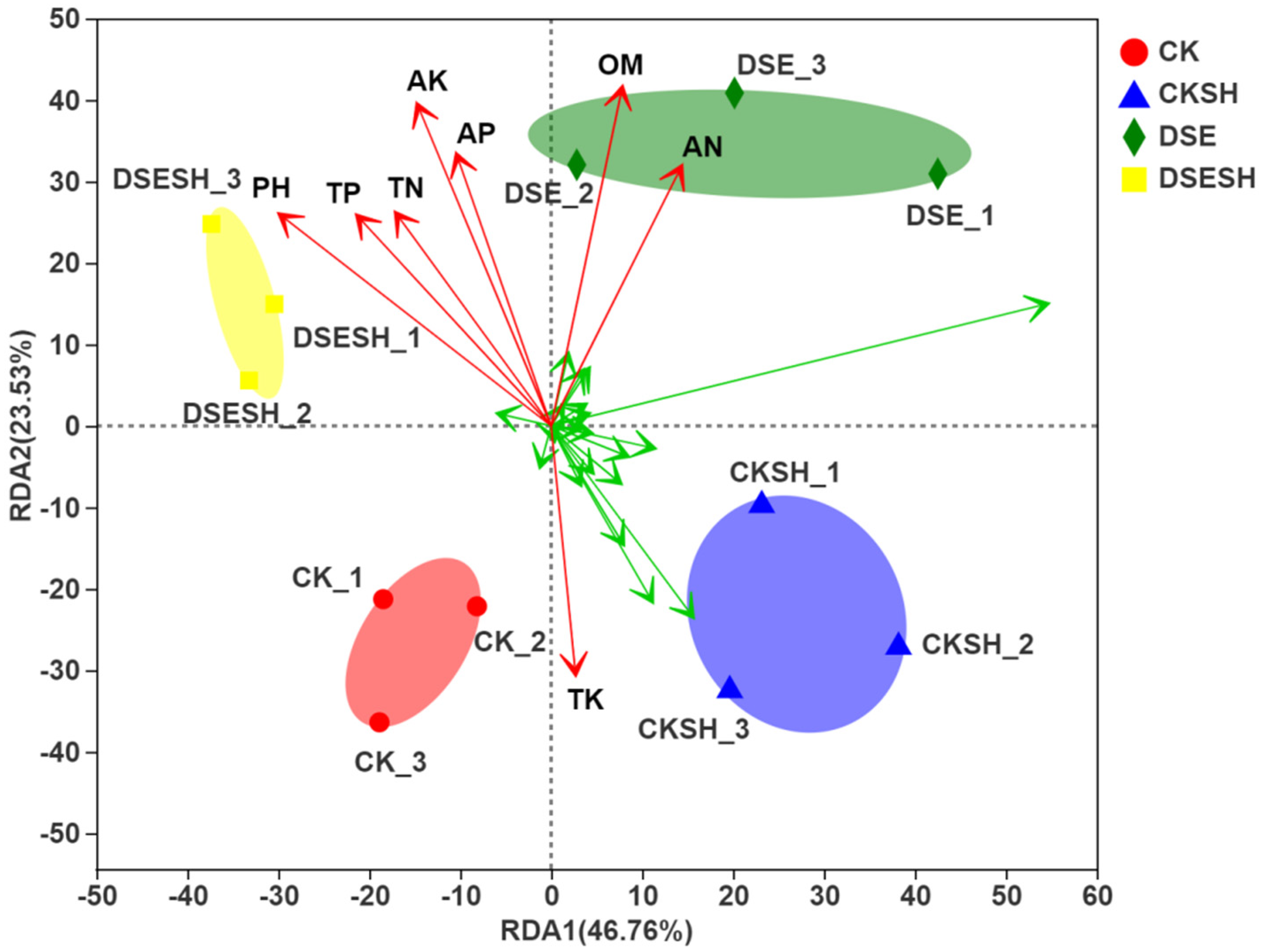
Figure 4). And the RDA analysis result indicated that DSE treatments soil bacterial community structure were positive correlated with AN, OM, AP, AK, TN, TP (
Figure 6), and this mean after DSE inoculation, enhance the soil nutrition, and then promote the growth of annual seedlings.
In conclusion, dark septate endophyte (DSE) A024 can control the rate of incidence of seedling damping-off disease and promote annual seedling growth after inoculation, and change the root structure of the P. sylvestris var. Mongolica seedlings, thereby elevating the seedling growth potential as well as absorption area. DSE A024 increased the activities of soil enzymes and contents of soil nutrients in the rhizosphere soil of P. sylvestris var. mongolica., particularly, the rhizosphere soil bacterial community structure of DSE A024 inoculation was positively correlated to the main soil nutrition indices.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}