First Come, First Served: Sui Generis Features of the First Intron


Abstract
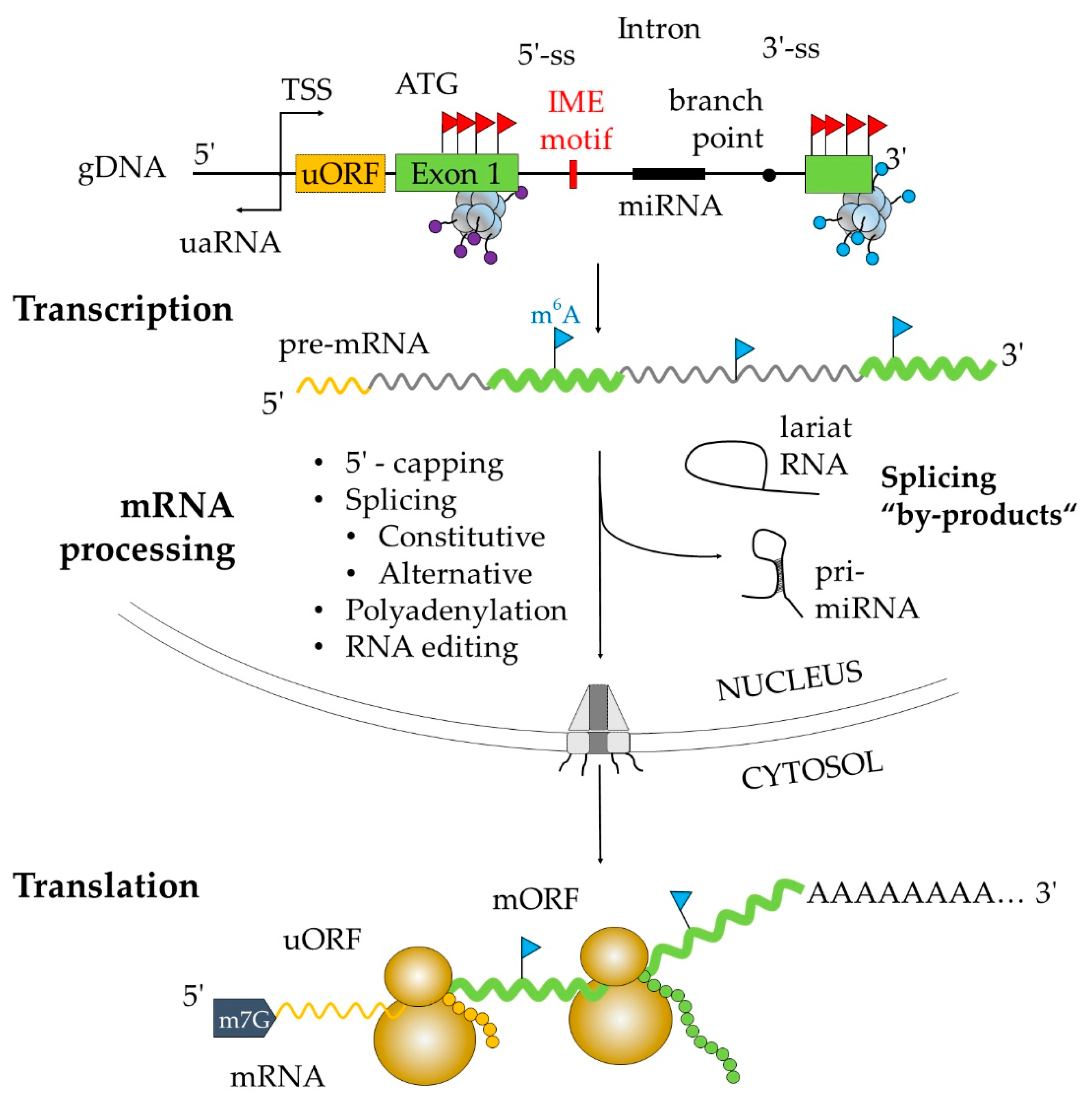
:1. Introduction
2. First Intronic DNA Sequence
3. Splicing is Predominantly Co-Transcriptional
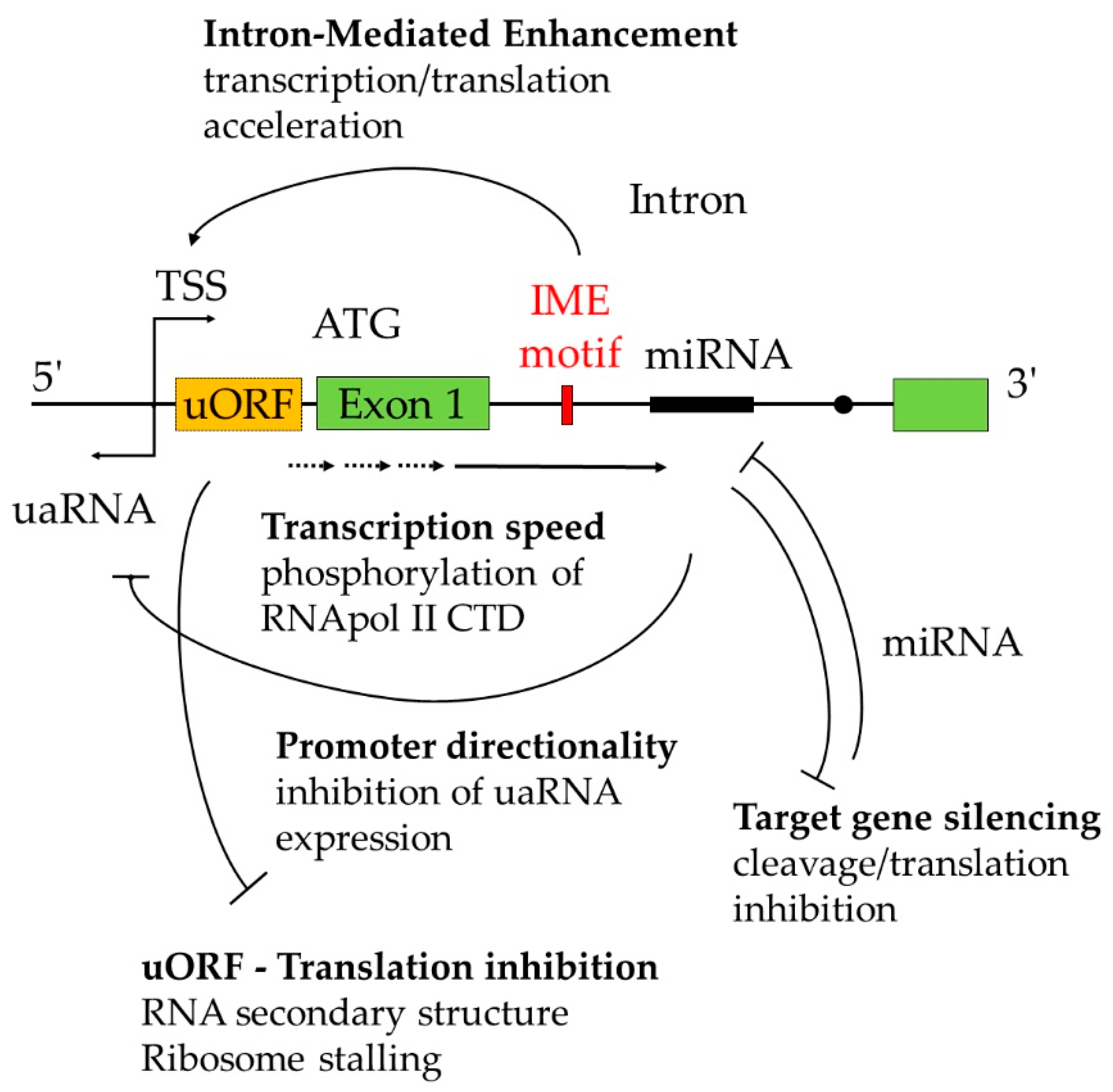
4. Intron-Mediated Enhancement (IME), a Mysterious Phenomenon
4.1. Splicing-Dependent IME
4.2. Splicing-Independent IME in Plants
5. Post-Transcriptional Regulation by Intron Retention (IR) within 5′-UTR
6. Regulation of First Intron Retention Located Downstream of ATG
7. AS in Response to Stresses and Developmental Cues
8. Intron Regulation beyond the Host Gene
9. Conclusions and Future Aspects
Author Contributions
Funding
Conflicts of Interest
References
- Berget, S.M.; Moore, C.; Sharp, P.A. Spliced segments at the 5′ terminus of adenovirus 2 late mRNA. Proc. Natl. Acad. Sci. USA 1977, 74, 3171–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, L.T.; Gelinas, R.E.; Broker, T.R.; Roberts, R.J. An amazing sequence arrangement at the 5′ ends of adenovirus 2 messenger RNA. Cell 1977, 12, 1–8. [Google Scholar] [CrossRef]
- Gilbert, W. Why genes in pieces? Nature 1978, 271, 501. [Google Scholar] [CrossRef]
- Catania, F.; Lynch, M. Where do introns come from? PLoS Biol. 2008, 6, 2354–2361. [Google Scholar] [CrossRef] [Green Version]
- Rogozin, I.B.; Carmel, L.; Csuros, M.; Koonin, E.V. Origin and evolution of spliceosomal introns. Biol. Direct 2012, 7, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irimia, M.; Roy, S.W. Origin of spliceosomal introns and alternative splicing. Cold Spring Harb. Perspect. Biol. 2014, 6, a016071. [Google Scholar] [CrossRef]
- Koonin, E.V.; Csuros, M.; Rogozin, I.B. Whence genes in pieces: Reconstruction of the exon-intron gene structures of the last eukaryotic common ancestor and other ancestral eukaryotes. Wiley Interdiscip. Rev. RNA 2013, 4, 93–105. [Google Scholar] [CrossRef]
- Csuros, M.; Rogozin, I.B.; Koonin, E.V. A detailed history of intron-rich eukaryotic ancestors inferred from a global survey of 100 complete genomes. PLoS Comput. Biol. 2011, 7, e1002150. [Google Scholar] [CrossRef] [Green Version]
- Braunschweig, U.; Gueroussov, S.; Plocik, A.M.; Graveley, B.R.; Blencowe, B.J. Dynamic integration of splicing within gene regulatory pathways. Cell 2013, 152, 1252–1269. [Google Scholar] [CrossRef] [Green Version]
- Barbazuk, W.B.; Fu, Y.; McGinnis, K.M. Genome-wide analyses of alternative splicing in plants: Opportunities and challenges. Genome Res. 2008, 18, 1382–1391. [Google Scholar] [CrossRef] [Green Version]
- Alló, M.; Schor, I.E.; Muñoz, M.J.; De La Mata, M.; Agirre, E.; Valcárcel, J.; Eyras, E.; Kornblihtt, A.R. Chromatin and alternative splicing. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Hir, H.; Nott, A.; Moore, M.J. How introns influence and enhance eukaryotic gene expression. Trends Biochem. Sci. 2003, 28, 215–220. [Google Scholar] [CrossRef]
- Szakonyi, D.; Duque, P. Alternative splicing as a regulator of early plant development. Front. Plant Sci. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, X.; Cao, Y.; Ma, L. Alternative splicing in plant genes: A means of regulating the environmental fitness of plants. Int. J. Mol. Sci. 2017, 18, 432. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, A.M.; Marone, D.; Laidò, G.; De Leonardis, A.M.; De Vita, P. Alternative splicing: Enhancing ability to cope with stress via transcriptome plasticity. Plant Sci. 2012, 185–186, 40–49. [Google Scholar] [CrossRef]
- Jabre, I.; Reddy, A.S.N.; Kalyna, M.; Chaudhary, S.; Khokhar, W.; Byrne, L.J.; Wilson, C.M.; Syed, N.H. Does co-transcriptional regulation of alternative splicing mediate plant stress responses? Nucl. Acids Res. 2019, 47, 2716–2726. [Google Scholar] [CrossRef]
- Park, S.G.; Hannenhalli, S.; Choi, S.S. Conservation in first introns is positively associated with the number of exons within genes and the presence of regulatory epigenetic signals. BMC Genomics 2014, 15, 526. [Google Scholar] [CrossRef] [Green Version]
- Jo, S.S.; Choi, S.S.; Hurst, L. Analysis of the functional relevance of epigenetic chromatin marks in the first intron associated with specific gene expression patterns. Genome Biol. Evol. 2019, 11, 786–797. [Google Scholar] [CrossRef]
- Zhang, X.; Bernatavichute, Y.V.; Cokus, S.; Pellegrini, M.; Jacobsen, S.E. Genome-wide analysis of mono-, di- and trimethylation of histone H3 lysine 4 in Arabidopsis thaliana. Genome Biol. 2009, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Meshorer, E.; Ast, G. Chromatin organization marks exon-intron structure. Nat. Struct. Mol. Biol. 2009, 16, 990–995. [Google Scholar] [CrossRef]
- Chodavarapu, R.K.; Feng, S.; Bernatavichute, Y.V.; Chen, P.Y.; Stroud, H.; Yu, Y.; Hetzel, J.A.; Kuo, F.; Kim, J.; Cokus, S.J.; et al. Relationship between nucleosome positioning and DNA methylation. Nature 2010, 466, 388–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pass, D.A.; Sornay, E.; Marchbank, A.; Crawford, M.R.; Paszkiewicz, K.; Kent, N.A.; Murray, J.A.H. Genome-wide chromatin mapping with size resolution reveals a dynamic sub-nucleosomal landscape in Arabidopsis. PLoS Genet. 2017, 13, e1006988. [Google Scholar] [CrossRef] [PubMed]
- Deremetz, A.; Le Roux, C.; Idir, Y.; Brousse, C.; Agorio, A.; Gy, I.; Parker, J.E.; Bouché, N. Antagonistic actions of FPA and IBM2 regulate transcript processing from genes containing heterochromatin. Plant Physiol. 2019, 180, 392–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieberstein, N.I.; Oesterreich, F.C.; Straube, K.; Neugebauer, K.M. First exon length controls active chromatin signatures and transcription. Cell Rep. 2012, 2, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Gelfman, S.; Cohen, N.; Yearim, A.; Ast, G. DNA-methylation effect on cotranscriptional splicing is dependent on GC architecture of the exon-intron structure. Genome Res. 2013, 23, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Lev Maor, G.; Yearim, A.; Ast, G. The alternative role of DNA methylation in splicing regulation. Trends Genet. 2015, 31, 274–280. [Google Scholar] [CrossRef]
- Wong, J.J.L.; Gao, D.; Nguyen, T.V.; Kwok, C.T.; Van Geldermalsen, M.; Middleton, R.; Pinello, N.; Thoeng, A.; Nagarajah, R.; Holst, J.; et al. Intron retention is regulated by altered MeCP2-mediated splicing factor recruitment. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ullah, F.; Hamilton, M.; Reddy, A.S.N.; Ben-Hur, A. Exploring the relationship between intron retention and chromatin accessibility in plants. BMC Genomics 2018, 19, 21. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hu, L.; Wang, X.; Li, N.; Xu, C.; Gong, L.; Liu, B. DNA methylation affects gene alternative splicing in plants: An example from rice. Mol. Plant 2016, 9, 305–307. [Google Scholar] [CrossRef] [Green Version]
- Chorev, M.; Joseph Bekker, A.; Goldberger, J.; Carmel, L. Identification of introns harboring functional sequence elements through positional conservation. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Lu, D.; Mason, A.S.; Li, B.; Xiao, M.; An, S.; Fu, D. Non-coding RNAs and transposable elements in plant genomes: Emergence, regulatory mechanisms and roles in plant development and stress responses. Planta 2019, 250, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Espinas, N.A.; Tu, L.N.; Furci, L.; Shimajiri, Y.; Harukawa, Y.; Miura, S.; Takuno, S.; Saze, H. Transcriptional regulation of genes bearing intronic heterochromatin in the rice genome. PLoS Genet. 2020, 16, e1008637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, T.N.; Miyazaki, Y.; Takuno, S.; Saze, H. Epigenetic regulation of intragenic transposable elements impacts gene transcription in Arabidopsis thaliana. Nucl. Acids Res. 2015, 43, 3911–3921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, K.; Liu, P.; Wu, C.A.; Yang, G.D.; Xu, R.; Guo, Q.H.; Huang, J.G.; Zheng, C.C. Stress-induced alternative splicing provides a mechanism for the regulation of microRNA processing in Arabidopsis thaliana. Mol. Cell 2012, 48, 521–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Shao, C.; Ma, X.; Wang, H. Introns targeted by plant microRNAs: A possible novel mechanism of gene regulation. Rice 2013, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Fan, X.; Lin, F.; He, G.; Terzaghi, W.; Zhu, D.; Deng, X.W. Arabidopsis noncoding RNA mediates control of photomorphogenesis by red light. Proc. Natl. Acad. Sci. USA 2014, 111, 10359–10364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qüesta, J.I.; Song, J.; Geraldo, N.; An, H.; Dean, C. Arabidopsis transcriptional repressor VAL1 triggers Polycomb silencing at FLC during vernalization. Science 2016, 353, 485–488. [Google Scholar] [CrossRef]
- Churchman, L.S.; Weissman, J.S. Nascent transcript sequencing visualizes transcription at nucleotide resolution. Nature 2011, 469, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Nojima, T.; Gomes, T.; Grosso, A.R.F.; Kimura, H.; Dye, M.J.; Dhir, S.; Carmo-Fonseca, M.; Proudfoot, N.J. Mammalian NET-seq reveals genome-wide nascent transcription coupled to RNA processing. Cell 2015, 161, 526–540. [Google Scholar] [CrossRef] [Green Version]
- Leng, X.; Ivanov, M.; Kindgren, P.; Malik, I.; Thieffry, A.; Brodersen, P.; Sandelin, A.; Kaplan, C.D.; Marquardt, S. Organismal benefits of transcription speed control at gene boundaries. EMBO Rep. 2020, 21, 1–16. [Google Scholar] [CrossRef]
- Godoy Herz, M.A.; Kornblihtt, A.R. Alternative splicing and transcription elongation in plants. Front. Plant Sci. 2019, 10, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Fong, N.; Kim, H.; Zhou, Y.; Ji, X.; Qiu, J.; Saldi, T.; Diener, K.; Jones, K.; Fu, X.D.; Bentley, D.L. Pre-mRNA splicing is facilitated by an optimal RNA polymerase II elongation rate. Genes Dev. 2014, 28, 2663–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujardin, G.; Lafaille, C.; de la Mata, M.; Marasco, L.E.; Muñoz, M.J.; Le Jossic-Corcos, C.; Corcos, L.; Kornblihtt, A.R. How slow RNA polymerase II elongation favors alternative exon skipping. Mol. Cell 2014, 54, 683–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Liu, M.; Liu, X.; Dong, Z. RNA polymerase II activity revealed by GRO-seq and pNET-seq in Arabidopsis. Nat. Plants 2018, 4, 1112–1123. [Google Scholar] [CrossRef]
- Wallace, E.W.J.; Beggs, J.D. Extremely fast and incredibly close: Cotranscriptional splicing in budding yeast. RNA 2017, 23, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nojima, T.; Rebelo, K.; Gomes, T.; Grosso, A.R.; Proudfoot, N.J.; Carmo-Fonseca, M. RNA Polymerase II phosphorylated on CTD serine 5 interacts with the spliceosome during co-transcriptional splicing. Mol. Cell 2018, 72, 369–379.e4. [Google Scholar] [CrossRef] [Green Version]
- Fong, N.; Saldi, T.; Sheridan, R.M.; Cortazar, M.A.; Bentley, D.L. RNA pol II dynamics modulate co-transcriptional chromatin modification, CTD phosphorylation, and transcriptional direction. Mol. Cell 2017, 66, 546–557.e3. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Long, Y.; Zhang, H.; Li, Z.; Liu, Z.; Zhao, Y.; Lu, D.; Jin, X.; Deng, X.; Xia, R.; et al. Post-transcriptional splicing of nascent RNA contributes to widespread intron retention in plants. Nat. Plants 2020, 6, 780–788. [Google Scholar] [CrossRef]
- Harlen, K.M.; Churchman, L.S. The code and beyond: Transcription regulation by the RNA polymerase II carboxy-terminal domain. Nat. Rev. Mol. Cell Biol. 2017, 18, 263–273. [Google Scholar] [CrossRef]
- Izaurralde, E.; Lewis, J.; McGuigan, C.; Jankowska, M.; Darzynkiewicz, E.; Mattaj, I.W. A nuclear cap binding protein complex involved in pre-mRNA splicing. Cell 1994, 78, 657–668. [Google Scholar] [CrossRef]
- Raczynska, K.D.; Simpson, C.G.; Ciesiolka, A.; Szewc, L.; Lewandowska, D.; McNicol, J.; Szweykowska-Kulinska, Z.; Brown, J.W.S.; Jarmolowski, A. Involvement of the nuclear cap-binding protein complex in alternative splicing in Arabidopsis thaliana. Nucl. Acids Res. 2009, 38, 265–278. [Google Scholar] [CrossRef]
- Laubinger, S.; Sachsenberg, T.; Zeller, G.; Busch, W.; Lohmann, J.U.; Rätsch, G.; Weigel, D. Dual roles of the nuclear cap-binding complex and SERRATE in pre-mRNA splicing and microRNA processing in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 8795–8800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raczynska, K.D.; Stepien, A.; Kierzkowski, D.; Kalak, M.; Bajczyk, M.; McNicol, J.; Simpson, C.G.; Szweykowska-Kulinska, Z.; Brown, J.W.S.; Jarmolowski, A. The SERRATE protein is involved in alternative splicing in Arabidopsis thaliana. Nucl. Acids Res. 2014, 42, 1224–1244. [Google Scholar] [CrossRef] [Green Version]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.S.; Crespi, M. Long noncoding RNA modulates alternative splicing regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callis, J.; Fromm, M.; Walbot, V. Introns increase gene expression in cultured maize cells. Genes Dev. 1987, 1, 1183–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, M.; Curtis Hannah, L. Splicing of the maize Sh1 first intron is essential for enhancement of gene expression, and a T-rich motif increases expression without affecting splicing. Plant Physiol. 2002, 130, 918–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Core, L.J.; Waterfall, J.J.; Lis, J.T. Nascent RNA sequencing reveals widespread pausing and divergent initiation at human promoters. Science 2008, 322, 1845–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Sharp, P.A. Divergent transcription: A driving force for new gene origination? Cell 2013, 155, 990–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, A.; Di Iulio, J.; Maleri, S.; Eser, U.; Vierstra, J.; Reynolds, A.; Sandstrom, R.; Stamatoyannopoulos, J.A.; Churchman, L.S. Native elongating transcript sequencing reveals human transcriptional activity at nucleotide resolution. Cell 2015, 161, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Hetzel, J.; Duttke, S.H.; Benner, C.; Chory, J. Nascent RNA sequencing reveals distinct features in plant transcription. Proc. Natl. Acad. Sci. USA 2016, 113, 12316–12321. [Google Scholar] [CrossRef] [Green Version]
- Furger, A.; O’Sullivan, J.M.; Binnie, A.; Lee, B.A.; Proudfoot, N.J. Promoter proximal splice sites enhance transcription. Genes Dev. 2002, 16, 2792–2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, N.; Ansari, A. Enhancement of transcription by a splicing-competent intron is dependent on promoter directionality. PLoS Genet. 2016, 12, 1006047. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.M.; Tan-Wong, S.M.; Morillon, A.; Lee, B.; Coles, J.; Mellor, J.; Proudfoot, N.J. Gene loops juxtapose promoters and terminators in yeast. Nat. Genet. 2004, 36, 1014–1018. [Google Scholar] [CrossRef]
- Moabbi, A.M.; Agarwal, N.; El Kaderi, B.; Ansari, A. Role for gene looping in intron-mediated enhancement of transcription. Proc. Natl. Acad. Sci. USA 2012, 109, 8505–8510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallegos, J.E.; Rose, A.B. The enduring mystery of intron-mediated enhancement. Plant Sci. 2015, 237, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.B. Introns as gene regulators: A brick on the accelerator. Front. Genet. 2019, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Vasil, V.; Clancy, M.; Ferl, R.J.; Vasil, I.K.; Hannah, L.C. Increased gene expression by the first intron of maize shrunken-1 locus in grass species. Plant. Physiol. 1989, 91, 1575–1579. [Google Scholar] [CrossRef] [Green Version]
- Clancy, M.; Vasil, V.; Curtis Hannah, L.; Vasil, I.K. Maize Shrunken-1 intron and exon regions increase gene expression in maize protoplasts. Plant Sci. 1994, 98, 151–161. [Google Scholar] [CrossRef]
- Donath, M.; Mendel, R.; Cerff, R.; Martin, W. Intron-dependent transient expression of the maize GapA1 gene. Plant Mol. Biol. 1995, 28, 667–676. [Google Scholar] [CrossRef]
- Vain, P.; Finer, K.R.; Engler, D.E.; Pratt, R.C.; Finer, J.J. Intron-mediated enhancement of gene expression in maize (Zea mays L.) and bluegrass (Poa pratensis L.). Plant. Cell Rep. 1996, 15, 489–494. [Google Scholar] [CrossRef]
- Morita, S.; Tsukamoto, S.; Sakamoto, A.; Makino, H.; Nakauji, E.; Kaminaka, H.; Masumura, T.; Ogihara, Y.; Satoh, S.; Tanaka, K. Differences in intron-mediated enhancement of gene expression by the first intron of cytosolic superoxide dismutase gene from rice in monocot and dicot plants. Plant Biotechnol. 2012, 29, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.B.; Last, R.L. Introns act post-transcriptionally to increase expression of the Arabidopsis thaliana tryptophan pathway gene PAT1. Plant J. 1997, 11, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Norris, S.R.; Meyer, S.E.; Callis, J. The intron of Arabidopsis thaliana polyubiquitin genes is conserved in location and is a quantitative determinant of chimeric gene expression. Plant Mol. Biol. 1993, 21, 895–906. [Google Scholar] [CrossRef] [PubMed]
- David-Assael, O.; Berezin, I.; Shoshani-Knaani, N.; Saul, H.; Mizrachy-Dagri, T.; Chen, J.; Brook, E.; Shaul, O. AtMHX is an auxin and ABA-regulated transporter whose expression pattern suggests a role in metal homeostasis in tissues with photosynthetic potential. Funct. Plant. Biol. 2006, 33, 661–672. [Google Scholar] [CrossRef]
- Akua, T.; Shaul, O. The Arabidopsis thaliana MHX gene includes an intronic element that boosts translation when localized in a 5′ UTR intron. J. Exp. Bot. 2013, 64, 4255–4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, A.B.; Elfersi, T.; Parra, G.; Korf, I. Promoter-proximal introns in Arabidopsis thaliana are enriched in dispersed signals that elevate gene expression. Plant Cell 2008, 20, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.B.; Carter, A.; Korf, I.; Kojima, N. Intron sequences that stimulate gene expression in Arabidopsis. Plant Mol. Biol. 2016, 92, 337–346. [Google Scholar] [CrossRef]
- Bradnam, K.R.; Korf, I. Longer first introns are a general property of eukaryotic gene structure. PLoS ONE 2008, 3. [Google Scholar] [CrossRef]
- Rose, A.B.; Beliakoff, J.A. Intron-mediated enhancement of gene expression independent of unique intron sequences and splicing. Plant Physiol. 2000, 122, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.B. Requirements for intron-mediated enhancement of gene expression in Arabidopsis. RNA 2002, 8, 1444–1453. [Google Scholar] [CrossRef] [Green Version]
- Emami, S.; Arumainayagam, D.; Korf, I.; Rose, A.B. The effects of a stimulating intron on the expression of heterologous genes in Arabidopsis thaliana. Plant Biotechnol. J. 2013, 11, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.M.; Mun, J.H.; Lee, I.; Woo, J.C.; Hong, C.B.; Kim, S.G. Distinct roles of the first introns on the expression of Arabidopsis profilin gene family members. Plant Physiol. 2006, 140, 196–209. [Google Scholar] [CrossRef] [Green Version]
- Gallegos, J.E.; Rose, A.B. Intron DNA sequences can be more important than the proximal promoter in determining the site of transcript initiation. Plant Cell 2017, 29, 843–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, A.B. The effect of intron location on intron-mediated enhancement of gene expression in Arabidopsis. Plant. J. 2004, 40, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, J.E.; Rose, A.B. An intron-derived motif strongly increases gene expression from transcribed sequences through a splicing independent mechanism in Arabidopsis thaliana. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Parra, G.; Bradnam, K.; Rose, A.B.; Korf, I. Comparative and functional analysis of intron-mediated enhancement signals reveals conserved features among plants. Nucl. Acids Res. 2011, 39, 5328–5337. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.B.; Emami, S.; Bradnam, K.; Korf, I. Evidence for A DNA-based mechanism of intron-mediated enhancement. Front. Plant Sci. 2011, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ner-Gaon, H.; Halachmi, R.; Savaldi-Goldstein, S.; Rubin, E.; Ophir, R.; Fluhr, R. Intron retention is a major phenomenon in alternative splicing in Arabidopsis. Plant J. 2004, 39, 877–885. [Google Scholar] [CrossRef]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.K.; Mockler, T.C. Genome-wide mapping of alternative splicing in Arabidopsis thaliana. Genome Res. 2010, 20, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Fesenko, I.; Khazigaleeva, R.; Kirov, I.; Kniazev, A.; Glushenko, O.; Babalyan, K.; Arapidi, G.; Shashkova, T.; Butenko, I.; Zgoda, V.; et al. Alternative splicing shapes transcriptome but not proteome diversity in Physcomitrella patens. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, S.; Khokhar, W.; Jabre, I.; Reddy, A.S.N.; Byrne, L.J.; Wilson, C.M.; Syed, N.H. Alternative splicing and protein diversity: Plants versus animals. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Tian, C.; Yu, Y.; Jiao, Y. Transcriptome Survey of the Contribution of Alternative Splicing to Proteome Diversity in Arabidopsis thaliana. Mol. Plant. 2016, 9, 749–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melamud, E.; Moult, J. Stochastic noise in splicing machinery. Nucl. Acids Res. 2009, 37, 4873–4886. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Lu, Y.; Zinta, G.; Lang, Z.; Zhu, J.K. UTR-dependent control of gene expression in plants. Trends Plant Sci. 2018, 23, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Kalyna, M.; Simpson, C.G.; Syed, N.H.; Lewandowska, D.; Marquez, Y.; Kusenda, B.; Marshall, J.; Fuller, J.; Cardle, L.; McNicol, J.; et al. Alternative splicing and nonsense-mediated decay modulate expression of important regulatory genes in Arabidopsis. Nucl. Acids Res. 2012, 40, 2454–2469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laxa, M.; Müller, K.; Lange, N.; Doering, L.; Pruscha, J.T.; Peterhänsel, C. The 5′UTR intron of arabidopsis GGT1 aminotransferase enhances promoter activity by recruiting RNA polymerase II. Plant. Physiol. 2016, 172, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Remy, E.; Cabrito, T.R.; Batista, R.A.; Hussein, M.A.M.; Teixeira, M.C.; Athanasiadis, A.; Sá-Correia, I.; Duque, P. Intron retention in the 5′UTR of the novel ZIF2 transporter enhances translation to promote zinc tolerance in Arabidopsis. PLoS Genet. 2014, 10, e1004375. [Google Scholar] [CrossRef] [Green Version]
- Sathe, L.; Bolinger, C.; Amin-Ul Mannan, M.; Dever, T.E.; Dey, M. Evidence that base-pairing interaction between intron and mRNA leader sequences inhibits initiation of HAC1 mRNA translation in yeast. J. Biol. Chem. 2015, 290, 21821–21832. [Google Scholar] [CrossRef] [Green Version]
- Von Arnim, A.G.; Jia, Q.; Vaughn, J.N. Regulation of plant translation by upstream open reading frames. Plant. Sci. 2014, 214, 1–12. [Google Scholar] [CrossRef]
- Hsu, P.Y.; Calviello, L.; Wu, H.Y.L.; Li, F.W.; Rothfels, C.J.; Ohler, U.; Benfey, P.N. Super-resolution ribosome profiling reveals unannotated translation events in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, E7126–E7135. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, A.G.; Ivanov, I.P.; Sonenberg, N. Translational control by 5′-untranslated regions of eukaryotic mRNAs. Science 2016, 352, 1413–1416. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.; Wardell, S.J.T.; Kleffmann, T.; Brown, C.M. The exon-intron gene structure upstream of the initiation codon predicts translation efficiency. Nucleic Acids Res. 2018, 46, 4575–4591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnstone, T.G.; Bazzini, A.A.; Giraldez, A.J. Upstream ORF s are prevalent translational repressors in vertebrates. EMBO J. 2016, 35, 706–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebina, I.; Takemoto-Tsutsumi, M.; Watanabe, S.; Koyama, H.; Endo, Y.; Kimata, K.; Igarashi, T.; Murakami, K.; Kudo, R.; Ohsumi, A.; et al. Identification of novel Arabidopsis thaliana upstream open reading frames that control expression of the main coding sequences in a peptide sequence-dependent manner. Nucl. Acids Res. 2015, 43, 1562–1576. [Google Scholar] [CrossRef] [Green Version]
- Calvo, S.E.; Pagliarini, D.J.; Mootha, V.K. Upstream open reading frames cause widespread reduction of protein expression and are polymorphic among humans. Proc. Natl. Acad. Sci. USA 2009, 106, 7507–7512. [Google Scholar] [CrossRef] [Green Version]
- Noh, A.L.; Watanabe, S.; Takahashi, H.; Naito, S.; Onouchi, H. An upstream open reading frame represses expression of a tomato homologue of arabidopsis ANAC096, a NAC domain transcription factor gene, in a peptide sequence-dependent manner. Plant Biotechnol. 2015, 32, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Henkin, T.M. Riboswitch RNAs: Using RNA to sense cellular metabolism. Genes Dev. 2008, 22, 3383–3390. [Google Scholar] [CrossRef] [Green Version]
- Sudarsan, N.; Barrick, J.E.; Breaker, R.R. Metabolite-binding RNA domains are present in the genes of eukaryotes. RNA 2003, 9, 644–647. [Google Scholar] [CrossRef] [Green Version]
- Bocobza, S.; Adato, A.; Mandel, T.; Shapira, M.; Nudler, E.; Aharoni, A. Riboswitch-dependent gene regulation and its evolution in the plant kingdom. Genes Dev. 2007, 21, 2874–2879. [Google Scholar] [CrossRef] [Green Version]
- Cheah, M.T.; Wachter, A.; Sudarsan, N.; Breaker, R.R. Control of alternative RNA splicing and gene expression by eukaryotic riboswitches. Nature 2007, 447, 497–500. [Google Scholar] [CrossRef]
- Álvarez, D.; Voß, B.; Maass, D.; Wüst, F.; Schaub, P.; Beyer, P.; Welsch, R. Carotenogenesis is regulated by 5′UTR-mediated translation of phytoene synthase splice variants. Plant. Physiol. 2016, 172, 2314–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grojean, J.; Downes, B. Riboswitches as hormone receptors: Hypothetical cytokinin-binding riboswitches in Arabidopsis thaliana. Biol. Direct 2010, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.S.N. Alternative splicing of pre-messenger RNAs in plants in the genomic era. Annu. Rev. Plant Biol. 2007, 58, 267–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.S.N.; Marquez, Y.; Kalyna, M.; Barta, A. Complexity of the alternative splicing landscape in plants. Plant Cell 2013, 25, 3657–3683. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Salvador, Ó.; Sánchez-Giménez, L.; López-Galiano, M.J.; Fernández-Crespo, E.; Schalschi, L.; García-Robles, I.; Rausell, C.; Real, M.D.; González-Bosch, C. The histone marks signature in exonic and intronic regions is relevant in early response of tomato genes to Botrytis cinerea and in miRNA regulation. Plants 2020, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Egawa, C.; Kobayashi, F.; Ishibashi, M.; Nakamura, T.; Nakamura, C.; Takumi, S. Differential regulation of transcript accumulation and alternative splicing of a DREB2 homolog under abiotic stress conditions in common wheat. Genes Genet. Syst. 2006, 81, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Chong, G.L.; Foo, M.H.; Lin, W.D.; Wong, M.M.; Verslues, P.E. Highly ABA-induced 1 (HAI1)-interacting protein HIN1 and drought acclimation-enhanced splicing efficiency at intron retention sites. Proc. Natl. Acad. Sci. USA 2019, 116, 22376–22385. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Li, J.; Gao, Z.; Lu, Y.; Yu, J.; Zheng, Q.; Yan, S.; Zhang, W.; He, H.; Ma, L.; et al. SKIP confers osmotic tolerance during salt stress by controlling alternative gene splicing in Arabidopsis. Mol. Plant 2015, 8, 1038–1052. [Google Scholar] [CrossRef] [Green Version]
- Leviatan, N.; Alkan, N.; Leshkowitz, D.; Fluhr, R. Genome-wide survey of cold stress regulated alternative splicing in Arabidopsis thaliana with tiling microarray. PLoS ONE 2013, 8, e66511. [Google Scholar] [CrossRef] [Green Version]
- Huertas, R.; Catalá, R.; Jiménez-Gómez, J.M.; Castellano, M.M.; Crevillén, P.; Piñeiro, M.; Jarillo, J.A.; Salinas, J. Arabidopsis SME1 regulates plant development and response to abiotic stress by determining spliceosome activity Specificity. Plant. Cell 2019, 31, 537–554. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Ryu, J.Y.; Baek, K.; Park, C.M. High temperature attenuates the gravitropism of inflorescence stems by inducing SHOOT GRAVITROPISM 5 alternative splicing in Arabidopsis. New Phytol. 2016, 209, 265–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, R.; Kathare, P.K.; Huq, E. Coordinated regulation of pre-mRNA splicing by the SFPS-RRC1 complex to promote photomorphogenesis. Plant Cell 2019, 31, 2052–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Chen, H.; Deng, X.W.; Irish, V.F.; Wei, N. Phytochrome B induces intron retention and translational inhibition of PHYTOCHROME-INTERACTING FACTOR3. Plant. Physiol. 2020, 182, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Ling, Y.; Serrano, N.; Gao, G.; Atia, M.; Mokhtar, M.; Woo, Y.H.; Bazin, J.; Veluchamy, A.; Benhamed, M.; Crespi, M.; et al. Thermopriming triggers splicing memory in Arabidopsis. J. Exp. Bot. 2018, 69, 2659–2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, A.B.; Syed, N.H.; Bordage, S.; Marshall, J.; Nimmo, G.A.; Jenkins, G.I.; Herzyk, P.; Brown, J.W.S.; Nimmo, H.G. Alternative splicing mediates responses of the Arabidopsis circadian clock to temperature changes. Plant. Cell 2012, 24, 961–981. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Y.; Lu, W.C.; Ko, S.S.; Sun, C.M.; Hung, J.C.; Chiou, T.J. Upstream open reading frame and phosphate-regulated expression of rice OsNLA1 controls phosphate transport and reproduction. Plant Physiol. 2020, 182, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Tsugeki, R.; Tanaka-Sato, N.; Maruyama, N.; Terada, S.; Kojima, M.; Sakakibara, H.; Okada, K. CLUMSY VEIN, the Arabidopsis DEAH-box Prp16 ortholog, is required for auxin-mediated development. Plant J. 2015, 81, 183–197. [Google Scholar] [CrossRef]
- Petrillo, E.; Godoy Herz, M.A.; Fuchs, A.; Reifer, D.; Fuller, J.; Yanovsky, M.J.; Simpson, C.; Brown, J.W.S.; Barta, A.; Kalyna, M.; et al. A chloroplast retrograde signal regulates nuclear alternative splicing. Science 2014, 344, 427–430. [Google Scholar] [CrossRef] [Green Version]
- Daras, G.; Rigas, S.; Alatzas, A.; Samiotaki, M.; Chatzopoulos, D.; Tsitsekian, D.; Papadaki, V.; Templalexis, D.; Banilas, G.; Athanasiadou, A.M.; et al. LEFKOTHEA Regulates Nuclear and Chloroplast mRNA Splicing in Plants. Dev. Cell 2019, 50, 767–779.e7. [Google Scholar] [CrossRef]
- Wahl, M.C.; Will, C.L.; Lührmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.W.S.; Marshall, D.F.; Echeverria, M. Intronic noncoding RNAs and splicing. Trends Plant Sci. 2008, 13, 335–342. [Google Scholar] [CrossRef]
- Rosado, A.; Li, R.; Van De Ven, W.; Hsu, E.; Raikhel, N.V. Arabidopsis ribosomal proteins control developmental programs through translational regulation of auxin response factors. Proc. Natl. Acad. Sci. USA 2012, 109, 19537–19544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallari, N.; Nibau, C.; Fuchs, A.; Dadarou, D.; Barta, A.; Doonan, J.H. The cyclin-dependent kinase G group defines a thermo-sensitive alternative splicing circuit modulating the expression of Arabidopsis ATU2AF65A. Plant J. 2018, 94, 1010–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parenteau, J.; Abou Elela, S. Introns: Good Day Junk Is Bad Day Treasure. Trends Genet. 2019, 35, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Munding, E.M.; Shiue, L.; Katzman, S.; Donohue, J.P.; Ares, M. Competition between Pre-mRNAs for the splicing machinery drives global regulation of splicing. Mol. Cell 2013, 51, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parenteau, J.; Maignon, L.; Berthoumieux, M.; Catala, M.; Gagnon, V.; Abou Elela, S. Introns are mediators of cell response to starvation. Nature 2019, 565, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.T.; Fink, G.R.; Bartel, D.P. Excised linear introns regulate growth in yeast. Nature 2019, 565, 606–611. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, S.; Cheng, J.; Su, C.; Zhong, S.; Liu, Q.; Fang, Y.; Yu, Y.; Lv, H.; Zheng, Y.; et al. Intron lariat RNA inhibits microRNA biogenesis by sequestering the dicing complex in Arabidopsis. PLoS Genet. 2016, 12, e1006422. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Category | Gene Name (Gene ID) | Position Relative to ATG | Function | Organism | Reference |
|---|---|---|---|---|---|
| IME | UBQ10 (At4g05320) | downstream | IME | Arabidopsis | [80] |
| PAT1/TRP1 (At5g17990) | downstream | IME | Arabidopsis | [72] | |
| ADH1 (GRMZM2G442658) | downstream | IME | maize | [55] | |
| Sh1 (GRMZM2G089713) | downstream | IME, splicing-dependent | maize | [56,68] | |
| PRF1 (At2g19760) | downstream | IME | Arabidopsis | [82] | |
| PRF2 (At4g29350) | downstream | IME | Arabidopsis | [82] | |
| miRNA/ | OsCLV1 (Os1g07060) | upstream | target of miR2123abc | rice | [35] |
| lncRNA | YSL3 (At5g53550) | upstream | miRNA-binding site | Arabidopsis | [35] |
| SLK2 (At5g62090) | upstream | miRNA-binding site | Arabidopsis | [35] | |
| SlyWRKY75 (Solyc05g015850) | downstream | target of sly-miR1127 | tomato | [115] | |
| n.a. (At1g32583) | upstream | source of miR400 | Arabidopsis | [34] | |
| n.a. (At5g08185) | downstream | source of miR162a | Arabidopsis | [131] | |
| NPF2.9/NRT1.9 (At1g18880) | downstream | source of miR837 | Arabidopsis | [131] | |
| n.a. (At2g23348) | upstream | source of miR844 | Arabidopsis | [131] | |
| PIF3 (At1g09530) | upstream | target of HID1 | Arabidopsis | [36] | |
| IR | |||||
| secondarystructure | ZIF2 (At2g48020) | upstream | translation enhancement | Arabidopsis | [97] |
| riboswitch | PSY (At5g17230) | upstream | translation inhibition | Arabidopsis | [111] |
| riboswitch | CRK10 (At4g23180) | downstream | putative cytokinin-binding riboswitch | Arabidopsis | [112] |
| uORF | PIF3 (At1g09530) | upstream | translation inhibition | Arabidopsis | [123] |
| uORF | ARF3 (At2g24765) | upstream | translation inhibition | Arabidopsis | [132] |
| uORF | OsNLA1 (Os07g0673200) | upstream | confers phosphate-dependent induction | Rice | [126] |
| IR | ATGGT-IB (At2g39550) | downstream | enhanced splicing upon cold | Arabidopsis | [119] |
| n.a. (At2g43160) | downstream | enhanced splicing upon cold | Arabidopsis | [119] | |
| n.a. (At3g47630) | downstream | enhanced splicing upon cold | Arabidopsis | [119] | |
| CDKG1 (At5g63370) | downstream | altered subcellular localization | Arabidopsis | [133] | |
| n.a. (At3g48070) | downstream | target of the LEF | Arabidopsis | [129] | |
| n.a. (At2g37510) | downstream | target of the LEF | Arabidopsis | [129] | |
| n.a. (At3g19920) | downstream | target of the LEF | Arabidopsis | [129] | |
| n.a. (At4g30993) | downstream | target of SME1 | Arabidopsis | [120] | |
| RD28/PIP2C (At2g37180) | downstream | target of SME1 | Arabidopsis | [120] | |
| n.a. (At3g26360) | downstream | target of SME1 | Arabidopsis | [120] | |
| TTA2/OBE4 (At3g63500) | downstream | target of SME1 | Arabidopsis | [120] | |
| IPGAM2 (At3g08590) | downstream | target of SME1 | Arabidopsis | [120] | |
| SPL2 (At5g43270) | downstream | target of SE | Arabidopsis | [53] | |
| APE2 (At5g46110) | downstream | target of SE | Arabidopsis | [53] | |
| HsfA2 (At2g26150) | downstream | target of SE | Arabidopsis | [53] | |
| SnRK2.8 (At1g78290) | downstream | target of SE | Arabidopsis | [53] | |
| YUC3 (At1g04610) | downstream | target of CUV | Arabidopsis | [127] | |
| SHY2/IAA3 (At1g04240) | downstream | target of CUV | Arabidopsis | [127] | |
| AXR3/IAA17 (At1g04250) | downstream | target of CUV | Arabidopsis | [127] | |
| NPH4/ARF7 (At5g20730) | downstream | target of CUV | Arabidopsis | [127] | |
| SLR/IAA14 (At4g14550) | downstream | target of CUV | Arabidopsis | [127] | |
| n.a. (At4G27050) | downstream | target of AtNSRa/b | Arabidopsis | [54] | |
| AtDRM2, DAP2 (At2G33830) | downstream | target of AtNSRa/b | Arabidopsis | [54] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalabák, D.; Ikeda, Y. First Come, First Served: Sui Generis Features of the First Intron. Plants 2020, 9, 911. https://doi.org/10.3390/plants9070911
Zalabák D, Ikeda Y. First Come, First Served: Sui Generis Features of the First Intron. Plants. 2020; 9(7):911. https://doi.org/10.3390/plants9070911
Chicago/Turabian StyleZalabák, David, and Yoshihisa Ikeda. 2020. "First Come, First Served: Sui Generis Features of the First Intron" Plants 9, no. 7: 911. https://doi.org/10.3390/plants9070911