Fungal Pathogens Affecting the Production and Quality of Medical Cannabis in Israel


Abstract
:1. Introduction
2. Results
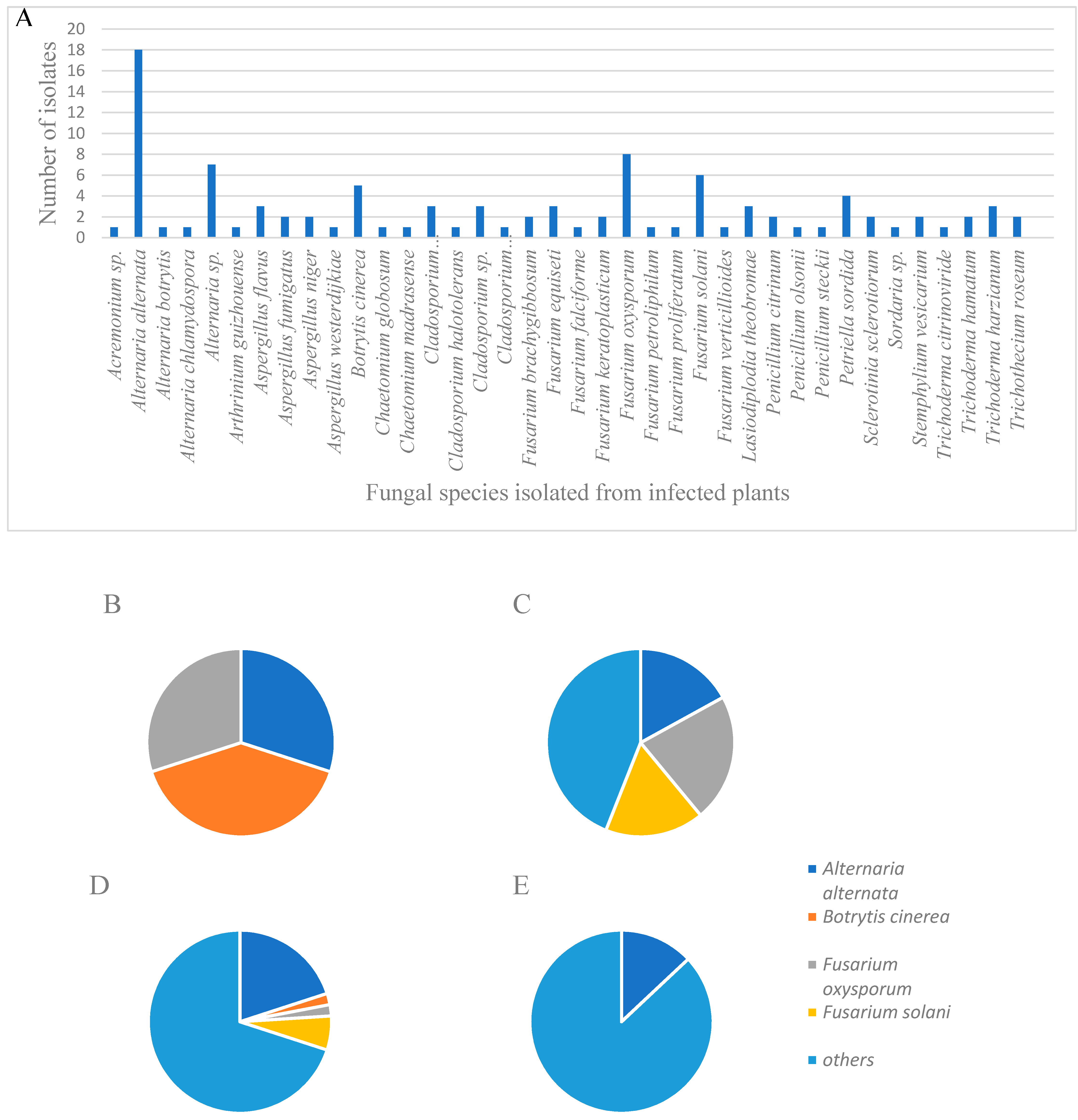
2.1. Fungal Isolations and Characterization
2.2. Koch Postulate Assays on Detached Tissues
2.3. Koch Postulate Assays on Intact Plants
2.4. Koch Postulate Assays of Soilborne Pathogens on Seedlings
3. Discussion
4. Materials and Methods
4.1. Isolation of Fungi and Growth Media
4.2. Fungal Identification and Characterization
4.3. Koch Postulate Assays
4.4. Production of Plant Material
4.5. Koch Postulate Assays on Detached Plant Tissues
4.6. Koch Postulate Assays on Intact Plants
4.7. Koch Postulate Assays on Seedlings for Soilborne Pathogens
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McPartland, J.M.; Clarke, R.C.; Watson, D.P. Hemp Diseases and Pests: Management and Biological Control—An Advanced Treatise; CABI: Wallingford, UK, 2017; ISBN 9780851994543. [Google Scholar]
- Ruchlemer, R.; Amit-Kohn, M.; Raveh, D.; Hanuš, L. Inhaled medicinal cannabis and the immunocompromised patient. Support. Care Cancer 2015, 23, 819–822. [Google Scholar] [CrossRef]
- Jerushalmi, S.; Maymon, M.; Dombrovsky, A.; Freeman, S. Effects of cold plasma, gamma and e-beam irradiations on reduction of fungal colony forming unit levels in medical cannabis inflorescences. J. Cannabis Res. 2020, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Vujanovic, V.; Korber, D.R.; Vujanovic, S.; Vujanovic, J.; Jabaji, S. Scientific prospects for cannabis-microbiome research to ensure quality and safety of products. Microorganisms 2020, 8, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, R.C.; Merlin, M.D. Cannabis domestication, breeding history, present-day genetic diversity, and future prospects. CRC. Crit. Rev. Plant Sci. 2016, 35, 293–327. [Google Scholar] [CrossRef]
- Small, E.; Marcus, D. Hemp: A New Crop with New Uses for North America. In Trends in New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 2019; pp. 1–39. [Google Scholar]
- Russo, E.B. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- Ben Amar, M. Cannabinoids in medicine: A review of their therapeutic potential. J. Ethnopharmacol. 2006, 105, 1–25. [Google Scholar] [CrossRef]
- Punja, Z.K. Flower and foliage-infecting pathogens of marijuana (Cannabis sativa L.) plants. Can. J. Plant Pathol. 2018, 40, 514–527. [Google Scholar] [CrossRef] [Green Version]
- Punja, Z.K.; Scott, C.; Chen, S. Root and crown rot pathogens causing wilt symptoms on field-grown marijuana (Cannabis sativa L.) plants. Can. J. Plant Pathol. 2018, 40, 528–541. [Google Scholar] [CrossRef] [Green Version]
- Jürgensen, C.W.; Madsen, A.M. Exposure to the airborne mould Botrytis and its health effects. Ann. Agric. Environ. Med. 2009, 16, 183–196. [Google Scholar]
- Lemons, A.R.; Nayak, A.P.; Couch, J.R.; Victory, K.R.; Beezhold, D.H.; Burton, N.C.; Green, B.J. Microbial hazards during harvesting and processing at an outdoor United States cannabis farm. J. Occup. Environ. Hyg. 2018. [Google Scholar] [CrossRef]
- Gargani, Y.; Bishop, P.; Denning, D.W. Too many mouldy joints—Marijuana and chronic pulmonary aspergillosis. Mediterr. J. Hematol. Infect. Dis. 2011, 3, e2011005. [Google Scholar] [CrossRef]
- Cascio, M.G.; Pertwee, R.G.; Marini, P. The pharmacology and therapeutic potential of plant cannabinoids. In Cannabis Sativa L.—Botany and Biotechnology; Springer International Publishing: Cham, Switzerland, 2017; pp. 207–225. ISBN 9783319545646. [Google Scholar]
- Sirikantaramas, S.; Taura, F. Cannabinoids: Biosynthesis and biotechnological applications. In Cannabis Sativa L.—Botany and Biotechnology; Springer International Publishing: Cham, Switzerland, 2017; pp. 183–206. ISBN 9783319545646. [Google Scholar]
- Akimitsu, K.; Tsuge, T.; Kodama, M.; Yamamoto, M.; Otani, H. Alternaria host-selective toxins: Determinant factors of plant disease. J. Gen. Plant Pathol. 2014, 80, 109–122. [Google Scholar] [CrossRef]
- Lawrence, C.B.; Mitchell, T.K.; Craven, K.D.; Cho, Y.; Cramer, R.A.; Kim, K.H. At death’s door: Alternaria pathogenicity mechanisms. Plant Pathol. J. 2008, 24, 101–111. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef]
- LaMondia, J.A. Outbreak of Brown Spot of Tobacco Caused by Alternaria alternata in Connecticut and Massachusetts. Plant Dis. 2001, 85, 230. [Google Scholar] [CrossRef]
- Gabriel, M.F.; Postigo, I.; Tomaz, C.T.; Martínez, J. Alternaria alternata allergens: Markers of exposure, phylogeny and risk of fungi-induced respiratory allergy. Environ. Int. 2016, 89–90, 71–80. [Google Scholar] [CrossRef]
- Pulimood, T.B.; Corden, J.M.; Bryden, C.; Sharples, L.; Nasser, S.M. Epidemic asthma and the role of the fungal mold Alternaria alternata. J. Allergy Clin. Immunol. 2007, 120, 610–617. [Google Scholar] [CrossRef]
- Scott, P.M.; Stoltz, D.R. Mutagens produced by Alternaria alternata. Mutat. Res. Toxicol. 1980, 78, 33–40. [Google Scholar] [CrossRef]
- Fillinger, S.; Elad, Y. Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems; Springer International Publishing: Cham, Switzerland, 2016; pp. 413–486. [Google Scholar]
- Petrasch, S.; Knapp, S.J.; van Kan, J.A.L.; Blanco-Ulate, B. Grey mould of strawberry, a devastating disease caused by the ubiquitous necrotrophic fungal pathogen Botrytis cinerea. Mol. Plant Pathol. 2019, 20, 877–892. [Google Scholar] [CrossRef] [Green Version]
- Pande, S.; Singh, G.; Rao, J.N.; Bakr, M.A.; Chaurasia, P.C.P.; Joshi, S.; Johansen, C.; Singh, S.D.; Kumar, J.; Rahman, M.M. Integrated Management of Botrytis Gray Mold of Chickpea; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; ISBN 9290664347. [Google Scholar]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium wilt of banana: Current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci. 2018, 871, 1468. [Google Scholar] [CrossRef] [Green Version]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.G. Influence of agricultural practices on Fusarium infection of cereals and subsequent contamination of grain by trichothecene mycotoxins. Toxicol. Lett. 2004, 153, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Placinta, C.M.; D’Mello, J.P.F.; MacDonald, A.M.C. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar] [CrossRef]
- da Rocha, M.E.B.; da Chagas Oliveira Freire, F.; Maia, F.E.F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Piacentini, K.C.; Iwase, C.H.T.; de Oliveira Rocha, L. Toxigenic Alternaria species: Impact in cereals worldwide. Curr. Opin. Food Sci. 2018, 23, 57–63. [Google Scholar] [CrossRef]
- Choi, Y.W.; Hyde, K.D.; Ho, W.W.H. Single spore isolation of fungi. Fungal Divers. 1999, 3, 29–38. [Google Scholar]
- Freeman, S.; Sharon, M.; Maymon, M.; Mendel, Z.; Protasov, A.; Aoki, T.; Eskalen, A.; O’Donnell, K. Fusarium euwallaceae sp. nov.—A symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia 2013, 105, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Gueidan, C.; Sink, S.; Johnston, P.R.; Crous, P.W.; Glenn, A.; Riley, R.; Zitomer, N.C.; Colyer, P.; Waalwijk, C.; et al. A two-locus DNA sequence database for typing plant and human pathogens within the Fusarium oxysporum species complex. Fungal Genet. Biol. 2009, 46, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dori-Bachash, M.; Avrahami-Moyal, L.; Protasov, A.; Mendel, Z.; Freeman, S. The occurrence and pathogenicity of Geosmithia spp. and common blue-stain fungi associated with pine bark beetles in planted forests in Israel. Eur. J. Plant Pathol. 2015, 143, 627–639. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Koch Postulate Assay a | |||
|---|---|---|---|
| Isolate | Species | Intact Plant | Detached Tissue |
| 22A | Alternaria alternata | + | + |
| 49A | Aspergillus flavus | ND b | + |
| 106A | Aspergillus fumigatus | ND | − |
| 53A | Aspergillus niger | ND | − |
| 71B | Aspergillus westerdijkiae | ND | + |
| 63A | Botrytis cinerea | + | + |
| 36B | Cladosporium cladosporioides | ND | + |
| 35A | Cladosporium halotolerans | ND | − |
| 32A | Cladosporium spp. | ND | + |
| 42A | Cladosporium sphaerospermum | ND | − |
| 119B | Cladosporium tenuissimum | ND | + |
| 10A | Fusarium equiseti | ND | + |
| 64B | Fusarium oxysporum | + | ND |
| 99A | Fusarium proliferatum | + | ND |
| 109A-3 | Fusarium solani | + | ND |
| 25A | Penicillium citrinum | ND | + |
| 110A | Penicillium olsonii | ND | − |
| 33B | Penicillium steckii | ND | + |
| 16A | Trichothecium roseum | ND | + |
| Primers Used for Fungal Identification with Corresponding Accession Numbers | |||||
|---|---|---|---|---|---|
| Isolate Designation | Fungal Species | ITS 4-5 | EF1 | RPB2 | IGS |
| 22A | Alternaria alternata | MT180724 | MT186261 | MT186265 | – a |
| 62A | Fusarium oxysporum | MT180726 | MT186263 | – | MT199205 |
| 63A | Botrytis cinerea | MT180725 | – | – | – |
| 64B | Fusarium oxysporum | MT180727 | MT186262 | – | MT199206 |
| 109A-3 | Fusarium solani | MT229135 | MT254546 | – | – |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jerushalmi, S.; Maymon, M.; Dombrovsky, A.; Freeman, S. Fungal Pathogens Affecting the Production and Quality of Medical Cannabis in Israel. Plants 2020, 9, 882. https://doi.org/10.3390/plants9070882
Jerushalmi S, Maymon M, Dombrovsky A, Freeman S. Fungal Pathogens Affecting the Production and Quality of Medical Cannabis in Israel. Plants. 2020; 9(7):882. https://doi.org/10.3390/plants9070882
Chicago/Turabian StyleJerushalmi, Shachar, Marcel Maymon, Aviv Dombrovsky, and Stanley Freeman. 2020. "Fungal Pathogens Affecting the Production and Quality of Medical Cannabis in Israel" Plants 9, no. 7: 882. https://doi.org/10.3390/plants9070882