Seed Total Protein Profiling in Discrimination of Closely Related Pines: Evidence from the Pinus mugo Complex

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
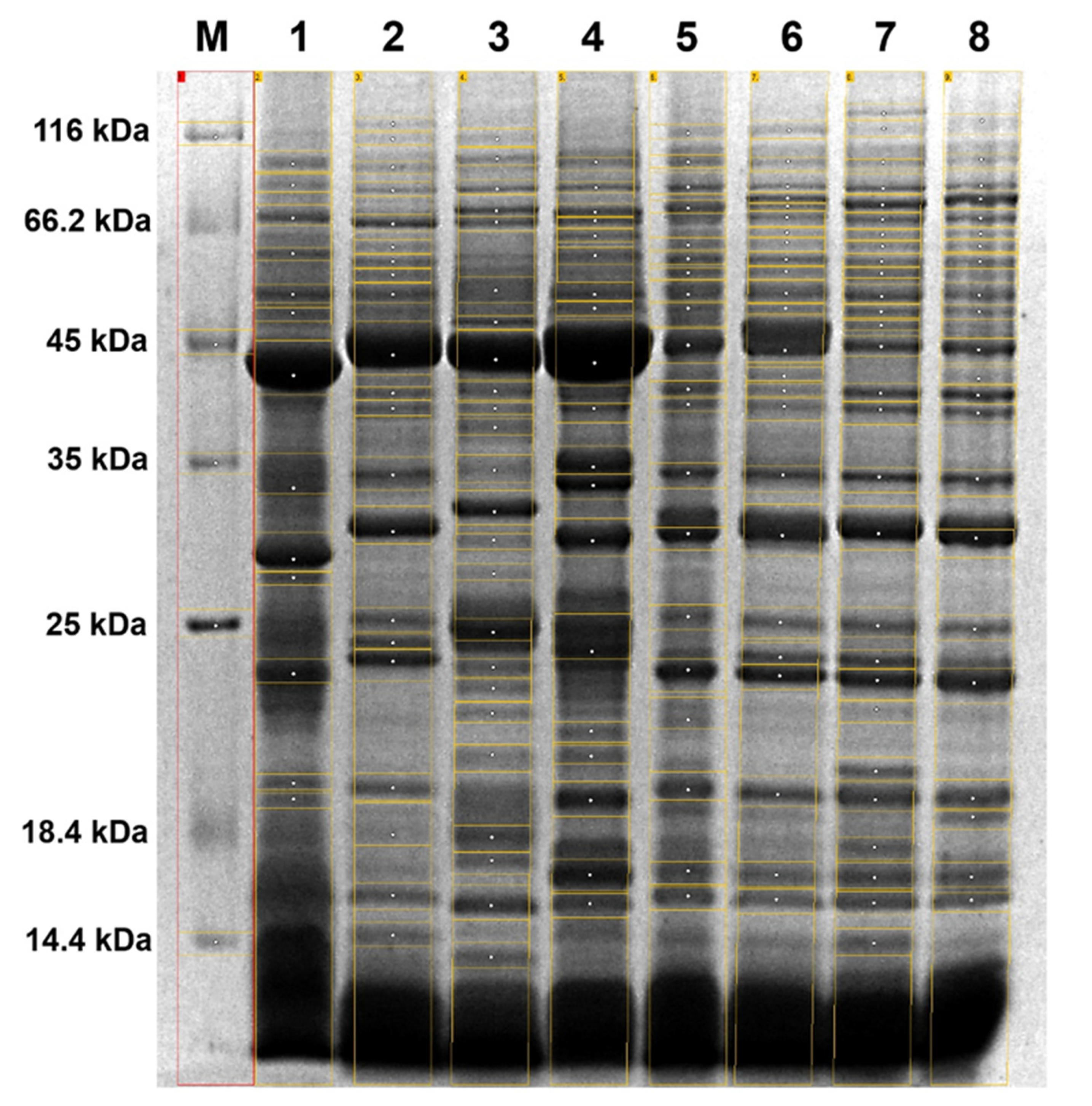
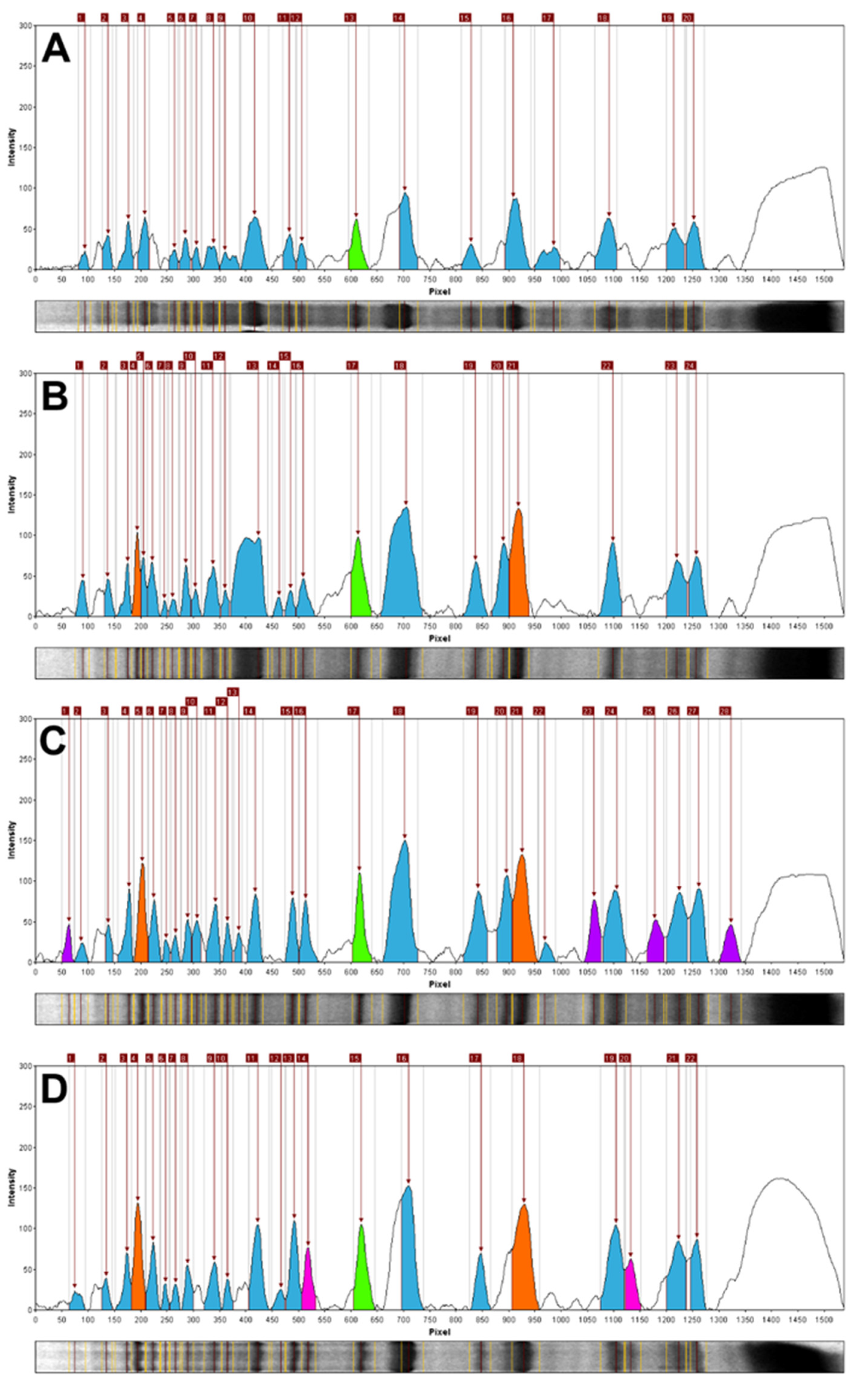
2.1. STPs Profiling—Interspecific Comparative Analysis
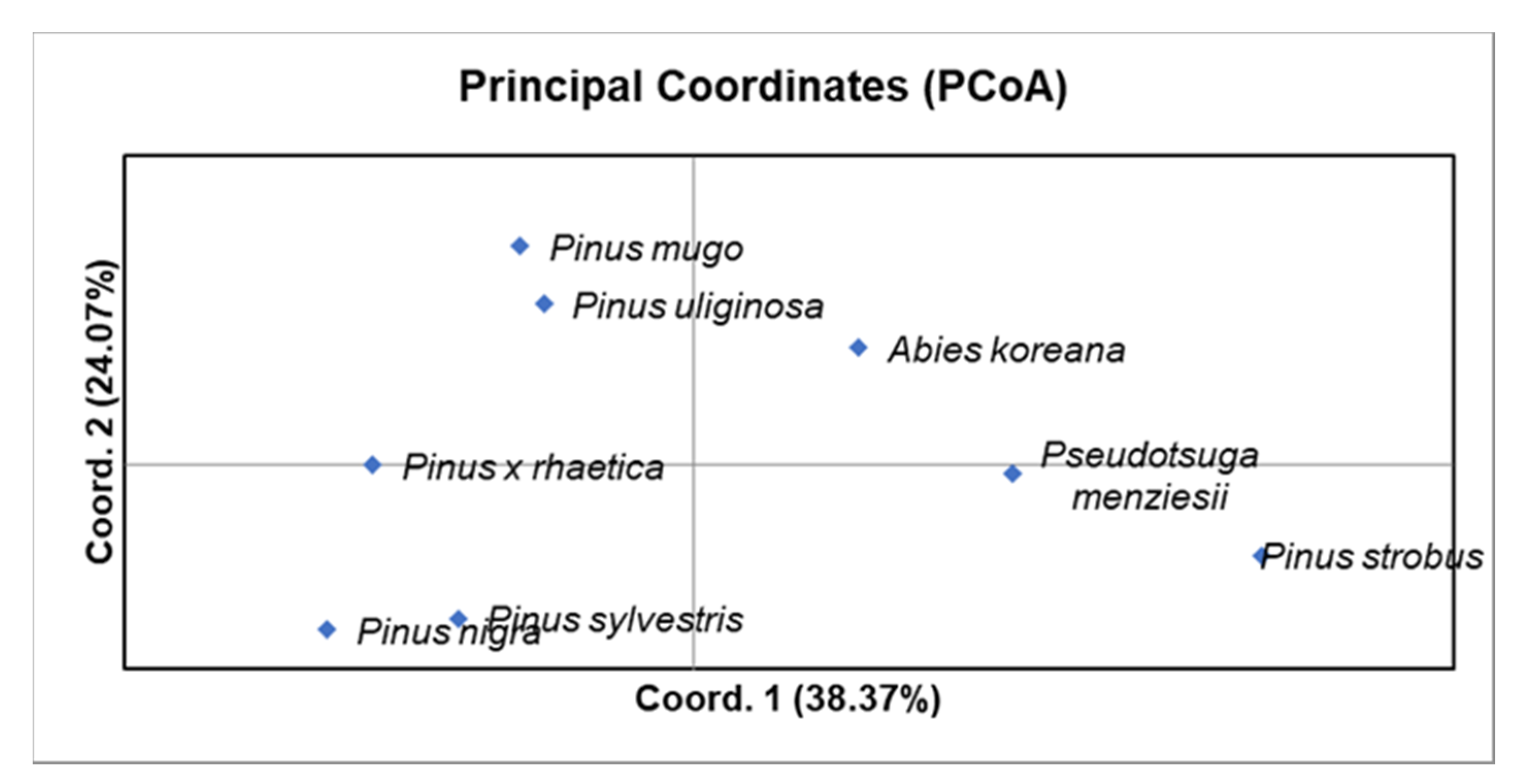
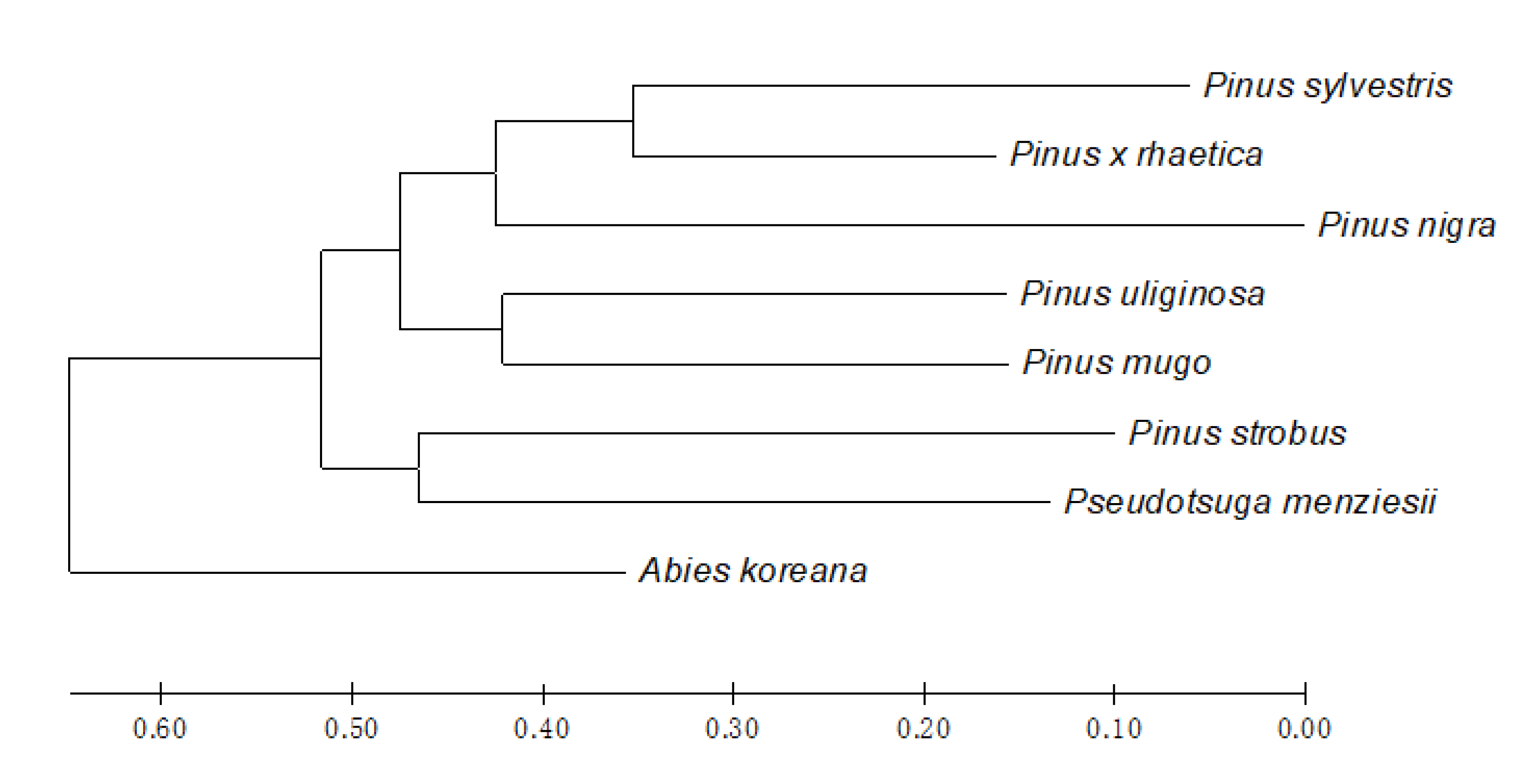
2.2. Genetic Distance and Cluster Analysis
3. Materials and Methods
3.1. Plant Material
3.2. Seed Total Protein (STP) Extraction
3.3. SDS-PAGE Electrophoresis
3.4. Band Scoring and Data Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Celiński, K.; Kijak, H.; Wojnicka-Półtorak, A.; Buczkowska-Chmielewska, K.; Sokołowska, J.; Chudzińska, E. Effectiveness of the DNA Barcoding Approach for Closely Related Conifers Discrimination: A Case Study of the Pinus Mugo Complex. Comptes Rendus Biol. 2017, 340, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Ladizinsky, G.; Hymowitz, T. Seed Protein Electrophoresis in Taxonomic and Evolutionary Studies. Theor. Appl. Genet. 1979, 54, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Slavov, G.T.; Zhelev, P. Allozyme Variation, Differentiation, and Inbreeding in Populations of Pinus Mugo in Bulgaria. Can. J. For. Res. 2004, 34, 2611–2617. [Google Scholar] [CrossRef] [Green Version]
- Goncharenko, G.G.; Silin, A.E.; Padutov, V.E. Allozyme Variation in Natural Populations of Eurasian Pines. 3.Population Structure, Diversity, Differentiation and Gene Flow in Central and Isolated Populations of Pinus Sylvestris L. in Eastern Europe and Siberia. Silvae Genet. 1994, 43, 119–131. [Google Scholar]
- Orav, A.; Kapp, K.; Raal, A. Chemosystematic markers for the essential oils in leaves of Mentha species cultivated or growing naturally in Estonia. Proc. Est. Acad. Sci. 2013, 62, 175–186. [Google Scholar] [CrossRef]
- Wawrzyniak, R.; Wasiak, W.; Ba̧czkiewicz, A.; Buczkowska, K. Volatile Compounds in Cryptic Species of the Aneura Pinguis Complex and Aneura Maxima (Marchantiophyta, Metzgeriidae). Phytochemistry 2014, 105, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Kaundun, S.S.; Lebreton, P. Taxonomy and Systematics of the Genus Pinus Based on Morphological, Biogeographical and Biochemical Characters. Plant Syst. Evol. 2010, 284, 1–15. [Google Scholar] [CrossRef]
- Ioannou, E.; Koutsaviti, A.; Tzakou, O.; Roussis, V. The Genus Pinus: A Comparative Study on the Needle Essential Oil Composition of 46 Pine Species. Phytochem. Rev. 2014, 13, 741–768. [Google Scholar] [CrossRef]
- Biteau, F.; Nisse, E.; Miguel, S.; Hannewald, P.; Bazile, V.; Gaume, L.; Mignard, B.; Hehn, A.; Bourgaud, F. A Simple SDS-PAGE Protein Pattern from Pitcher Secretions as a New Tool to Distinguish Nepenthes Species (Nepenthaceae). Am. J. Bot. 2013, 100, 2478–2484. [Google Scholar] [CrossRef] [PubMed]
- Schlötterer, C. The Evolution of Molecular Markers—Just a Matter of Fashion? Nat. Rev. Genet. 2004, 5, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Hollingsworth, M.L.; Andra Clark, A.; Forrest, L.L.; Richardson, J.; Pennington, R.T.; Long, D.G.; Cowan, R.; Chase, M.W.; Gaudeul, M.; Hollingsworth, P.M. Selecting Barcoding Loci for Plants: Evaluation of Seven Candidate Loci with Species-Level Sampling in Three Divergent Groups of Land Plants. Mol. Ecol. Resour. 2009, 9, 439–457. [Google Scholar] [CrossRef]
- Han, Y.W.; Duan, D.; Ma, X.F.; Jia, Y.; Liu, Z.L.; Zhao, G.F.; Li, Z.H. Efficient Identification of the Forest Tree Species in Aceraceae Using DNA Barcodes. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, D.P.; Knopf, P.; Schulz, C. DNA Barcode Identification of Podocarpaceae-The Second Largest Conifer Family. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- von Cräutlein, M.; Korpelainen, H.; Pietiläinen, M.; Rikkinen, J. DNA Barcoding: A Tool for Improved Taxon Identification and Detection of Species Diversity. Biodivers. Conserv. 2011, 20, 373–389. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA Barcoding: From Gene to Genome. Biol. Rev. Camb. Philos. Soc. 2015, 90, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, D.F.; Guo, X.L.; Zheng, Z.Y.; He, X.J.; Zhou, S.D. Comparative Analysis of the Complete Plastid Genome of Five Bupleurum Species and New Insights into Dna Barcoding and Phylogenetic Relationship. Plants 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Song, M.; Guan, Y.; Ma, X. Species Identification of Dracaena Using the Complete Chloroplast Genome as a Super-Barcode. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Samah, S.; Ventura-Zapata, E.; Valadez-Moctezuma, E. Fractionation and Electrophoretic Patterns of Seed Protein of Opuntia Genus. A Preliminary Survey as a Tool for Accession Differentiation and Taxonomy. Biochem. Syst. Ecol. 2015, 58, 187–194. [Google Scholar] [CrossRef]
- Raj, L.J.M.; Britto, S.J.; Prabhu, S.; Senthilkumar, S.R. Phylogenetic Relationships of Crotalaria Species Based on Seed Protein Polymorphism Revealed by SDS-PAGE. Plant Sci. 2011, 2, 119–128. [Google Scholar]
- Aniel Kumar, O.; Subba Tata, S. SDS-Page Seed Storage Protein Profiles in Chili Peppers (Capsicum L.). Not. Sci. Biol. 2010, 2, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Sinha, K.N. Electrophoretic Study of Seed Storage Protein in Five Species of Bauhinia. IOSR J. Pharm. Biol. Sci. 2012, 4, 8–11. [Google Scholar] [CrossRef]
- Emre, I.; Turgut-Balik, D.; Genç, H.; Şahin, A. Total Seed Storage Protein Patterns of Some Lathyrus Species Growing in Turkey Using SDS-Page. Pak. J. Bot. 2010, 42, 3157–3163. [Google Scholar]
- Ertugrul, K.; Arslan, E.; Tugay, O. Characterization of Conolida S.F. Gray (Ranunculaceae) taxa in Turkey by seed storage protein electrophoresis. Turk. J. Biochem. 2010, 35, 99–104. [Google Scholar]
- El Rabey, H.A.; Al-Malki, A.L.; Abulnaja, K.O.; Ebrahim, M.K.; Kumosani, T.; Khan, J.A. Phylogeny of Ten Species of the Genus Hordeum L. as Revealed by AFLP Markers and Seed Storage Protein Electrophoresis. Mol. Biol. Rep. 2014, 41, 364–372. [Google Scholar] [CrossRef]
- Christensen, K. Taxonomic Revision of the Pinus Mugo Complex and P. Rhaetica (P. Mugo Sylvestris) (Pinaceae). Nord. J. Bot. 1987, 7, 383–408. [Google Scholar] [CrossRef]
- Hamerník, J.; Musil, I. The Pinus Mugo Complex—Its Structuring and General Overview of the Used Nomenclature. J. For. Sci. 2007, 53, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Wachowiak, W.; Prus-Głowacki, W. Hybridisation Processes in Sympatric Populations of Pines Pinus sylvestris L., P. mugo Turra and P. uliginosa Neumann. Plant Syst. Evol. 2008, 271, 29–40. [Google Scholar] [CrossRef]
- Prus-Głowacki, W.; Szweykowski, J.; Nowak, R. Serotaxonomical Investigation of the European Pine Species. Silvae Genet. 1985, 34, 162–170. [Google Scholar]
- Lewandowski, A.; Boratyński, A.; Mejnartowicz, L. Allozyme Investigations on the Genetic Differentiation between Closely Related Pines—Pinus sylvestris, P. mugo, P. uncinata, and P. uliginosa (Pinaceae). Plant Syst. Evol. 2000, 221, 15–24. [Google Scholar] [CrossRef]
- Prus-Głowacki, W.; Bujas, E.; Ratyńska, H. Taxonomic Position of Pinus Uliginosa Neumann as Related to Other Taxa of Pinus Mugo Complex. Acta Soc. Bot. Pol. 1998, 67, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Monteleone, I.; Ferrazzini, D.; Belletti, P. Effectiveness of Neutral RAPD Markers to Detect Genetic Divergence between the Subspecies uncinata and mugo of Pinus mugo Turra. Silva Fenn. 2006, 40, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Bogunić, F.; Siljak-Yakovlev, S.; Muratović, E.; Pustahija, F.; Medjedović, S. Molecular Cytogenetics and Flow Cytometry Reveal Conserved Genome Organization in Pinus Mugo and P. Uncinata. Annals Forest Sci. 2011, 68, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Celiński, K.; Chudzińska, E.; Gmur, A.; Piosik, Ł.; Wojnicka-Półtorak, A. Cytological Characterization of Three Closely Related Pines—Pinus Mugo, P. Uliginosa and P. × Rhaetica from the Pinus Mugo Complex (Pinaceae). Biologia 2019, 74, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Bonikowski, R.; Celiński, K.; Wojnicka-Półtorak, A.; Maliński, T. Composition of Essential Oils Isolated from the Needles of Pinus Uncinata and P. Uliginosa Grown in Poland. Nat. Prod. Commun. 2015, 10, 371–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celiński, K.; Bonikowski, R.; Wojnicka-Półtorak, A.; Chudzińska, E.; Maliński, T. Volatiles as Chemosystematic Markers for Distinguishing Closely Related Species within the Pinus Mugo Complex. Chem. Biodivers. 2015, 12, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Juan, R.; Pastor, J.; Alaiz, M.; Vioque, J. Electrophoretic Characterization of Amaranthus L. Seed Proteins and Its Systematic Implications. Bot. J. Linn. Soc. 2007, 155, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Lewandowski, A.; Wiśniewska, M. Short Note: Crossability between Pinus Uliginosa and Its Putative Parental Species Pinus Sylvestris and Pinus Mugo. Silvae Genet. 2006, 55, 52–54. [Google Scholar] [CrossRef] [Green Version]
- Kormutak, A.; Demankova, B.; Gömöry, D. Spontaneous Hybridization between Pinus Sylvestris L. and P. Mugo Turra in Slovakia. Silvae Genet. 2008, 57, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Wachowiak, W.; Celiński, K.; Prus-Głowacki, W. Evidence of Natural Reciprocal Hybridisation between Pinus Uliginosa and P. Sylvestris in the Sympatric Population of the Species. Flora Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 563–568. [Google Scholar] [CrossRef]
- Bhat, T.M.; Kudesia, R. Evaluation of Genetic Diversity in Five Different Species of Family Solanaceae Using Cytological Characters and Protein Profiling. Genet. Eng. Biotechnol. J. 2011, 1–8. Available online: http://astonjournals.com/gebj (accessed on 10 June 2020).
- Schirone, B.; Piovesan, G.; Bellarosa, R.; Pelosi, C. A Taxonomic Analysis of Seed Proteins in Pinus Spp. (Pinaceae). Plant Syst. Evol. 1991, 178, 43–53. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tomooka, N. Experimental Manual of SDS-PAGE (Slab Method): Progress Report; Tropical Agriculture Research Center: Okinawa, Japan, 1989; p. 13. [Google Scholar]
- Popov, N.; Schmitt, M.; Schulzeck, S.; Matthies, H. Reliable Micromethod for Determination of the Protein Content in Tissue Homogenates. Acta Biol. Med. Ger. 1975, 34, 1441–1446. [Google Scholar]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Hames, B.D.; Rickwood, D. One dimensional polyacrylamide gel electrophoresis. In Gel Electrophoresis of Proteins, 2nd ed.; Hames, B.D., Rickwood, D., Eds.; Oxford University Press: New York, NY, USA, 1990; p. 382. [Google Scholar]
- GelAnalyzer. Available online: http://www.gelanalyzer.com/?i=1 (accessed on 12 June 2020).
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research-an Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Kovach, W.L. MVSP—A MultiVariate Statistical Package for Windows, ver. 3.1. Kovach Computing Services; Pentraeth: Wales, UK, 2007. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Taxon | NB | Rf Range | Bands Relative Mobility Value (Rf) | NUB |
|---|---|---|---|---|---|
| 1 | A. koreana | 13 | 0.09–0.72 | 0.09, 0.11, 0.15, 0.18, 0.22, 0.24, 0.30, 0.41, 0.48, 0.50, 0.60, 0.70, 0.72 | 2 |
| 2 | P. nigra | 21 | 0.05–0.85 | 0.05, 0.07, 0.10, 0.12, 0.15, 0.17, 0.19, 0.20, 0.22, 0.28. 0.32. 0.33, 0.40, 0.45, 0.54, 0.56, 0.58, 0.71, 0.75, 0.81, 0.85 | 5 |
| 3 | P. strobus | 24 | 0.07–0.87 | 0.07, 0.09, 0.12, 0.14, 0.15, 0.22, 0.25, 0.29, 0.32, 0.33, 0.35, 0.39, 0.44, 0.46, 0.50, 0.55, 0.59, 0.61, 0.63, 0.67, 0.76, 0.78, 0.82, 0.87 | 7 |
| 4 | P. menziesii | 19 | 0.09–0.82 | 0.09, 0.12, 0.14, 0.15, 0.16, 0.18, 0.22, 0.23, 0.29, 0.33, 0.39, 0.41, 0.46, 0.57, 0.65, 0.68, 0.72, 0.79, 0.82 | 3 |
| 5 | P. sylvestris | 20 | 0.06–0.81 | 0.06, 0.09, 0.11, 0.14, 0.17, 0.19, 0.20, 0.22, 0.23, 0.27, 0.31, 0.33, 0.40, 0.46, 0.54, 0.59, 0.64, 0.71, 0.79, 0.81 | 2 |
| 6 | P. × rhaetica | 24 | 0.06–0.82 | 0.06, 0.09, 0.11, 0.12, 0.13, 0.14, 0.16, 0.17, 0.19, 0.20, 0.22, 0.23, 0.28, 0.30, 0.32, 0.33, 0.40, 0.46, 0.54, 0.58, 0.60, 0.71, 0.79, 0.82 | 0 |
| 7 | P. uliginosa | 28 | 0.04–0.86 | 0.04, 0.06, 0.09, 0.12, 0.13, 0.15, 0.16, 0.17, 0.19, 0.20, 0.22, 0.24, 0.25, 0.27, 0.32, 0.33, 0.40, 0.46, 0.55, 0.58, 0.60, 0.63, 0.69, 0.72, 0.77, 0.80, 0.82, 0.86 | 4 |
| 8 | P. mugo | 22 | 0.05–0.82 | 0.05, 0.09, 0.11, 0.13, 0.15, 0.16, 0.17, 0.19, 0.22, 0.24, 0.28, 0.30, 0.32, 0.34, 0.40, 0.46, 0.55, 0.60, 0.72, 0.74, 0.80, 0.82 | 2 |
| A. koreana | P. nigra | P. strobus | P. menziesii | P. sylvestris | P. × rhaetica | P. uliginosa | P. mugo | |
|---|---|---|---|---|---|---|---|---|
| A. koreana | - | 0.067 | 0.121 | 0.231 | 0.100 | 0.156 | 0.171 | 0.296 |
| P. nigra | 0.606 | - | 0.103 | 0.086 | 0.258 | 0.265 | 0.205 | 0.171 |
| P. strobus | 0.579 | 0.693 | - | 0.303 | 0.158 | 0.200 | 0.268 | 0.179 |
| P. menziesii | 0.361 | 0.663 | 0.428 | - | 0.219 | 0.303 | 0.237 | 0.206 |
| P. sylvestris | 0.526 | 0.428 | 0.663 | 0.476 | - | 0.517 | 0.263 | 0.200 |
| P. × rhaetica | 0.526 | 0.383 | 0.663 | 0.428 | 0.238 | - | 0.444 | 0.438 |
| P. uliginosa | 0.579 | 0.579 | 0.606 | 0.579 | 0.552 | 0.361 | - | 0.471 |
| P. mugo | 0.340 | 0.526 | 0.663 | 0.526 | 0.552 | 0.318 | 0.318 | - |
| No. | Taxon | Subfamily | Genus | Subgenus/Section | Seeds Origin |
|---|---|---|---|---|---|
| 1 | Abies koreana E.H. Wilson | Abietoideae | Abies | - | BGL |
| 2 | Pinus nigra Arn. | Pinoideae | Pinus | Pinus | BGL |
| 3 | Pinus strobus L. | Pinoideae | Pinus | Strobus | BGL |
| 4 | Pseudotsuga menziesii (Mirbel) Franco | Laricoideae | Pseudotsuga | - | BGL |
| 5 | Pinus sylvestris L. | Pinoideae | Pinus | Pinus | BGL |
| 6 | Pinus × rhaetica Brügger* | Pinoideae | Pinus | Pinus | DGP |
| 7 | Pinus uliginosa Neumann ex Wimmer* | Pinoideae | Pinus | Pinus | DGP |
| 8 | Pinus mugo Turra* | Pinoideae | Pinus | Pinus | DGP |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Celiński, K.; Sokołowska, J.; Zemleduch-Barylska, A.; Kuna, R.; Kijak, H.; Staszak, A.M.; Wojnicka-Półtorak, A.; Chudzińska, E. Seed Total Protein Profiling in Discrimination of Closely Related Pines: Evidence from the Pinus mugo Complex. Plants 2020, 9, 872. https://doi.org/10.3390/plants9070872
Celiński K, Sokołowska J, Zemleduch-Barylska A, Kuna R, Kijak H, Staszak AM, Wojnicka-Półtorak A, Chudzińska E. Seed Total Protein Profiling in Discrimination of Closely Related Pines: Evidence from the Pinus mugo Complex. Plants. 2020; 9(7):872. https://doi.org/10.3390/plants9070872
Chicago/Turabian StyleCeliński, Konrad, Joanna Sokołowska, Agata Zemleduch-Barylska, Roman Kuna, Hanna Kijak, Aleksandra Maria Staszak, Aleksandra Wojnicka-Półtorak, and Ewa Chudzińska. 2020. "Seed Total Protein Profiling in Discrimination of Closely Related Pines: Evidence from the Pinus mugo Complex" Plants 9, no. 7: 872. https://doi.org/10.3390/plants9070872