
The Assessment and the Within-Plant Variation of the Morpho-Physiological Traits and VOCs Profile in Endemic and Rare Salvia ceratophylloides Ard. (Lamiaceae)

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
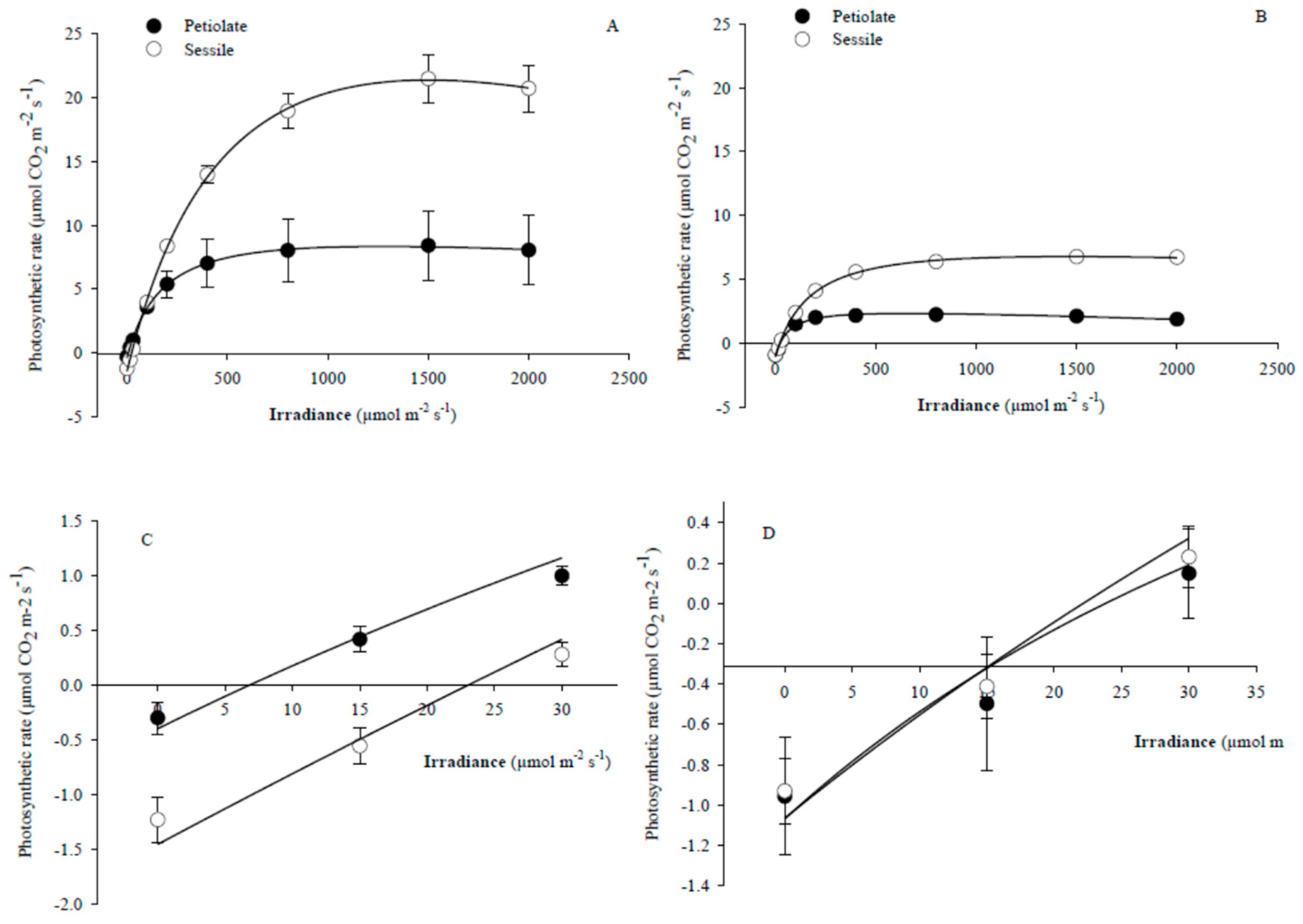
2.1. Physiological Performances and Morphological Traits of S. ceratophylloides
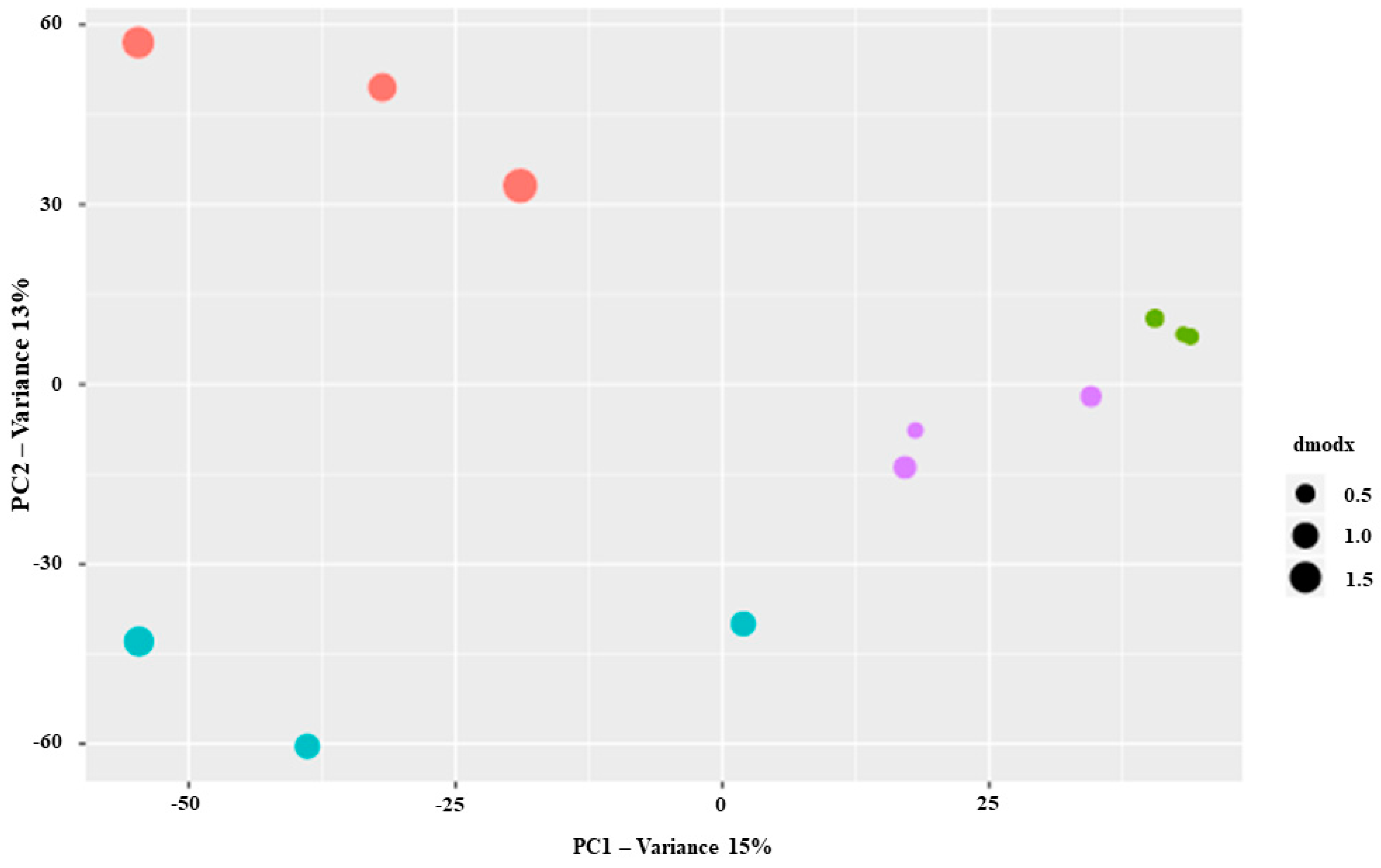
2.2. VOCs Analysis of Salvia ceratophylloides in Its Habitat
3. Discussion
3.1. The Assessment of the Morpho-Physiological Traits of the Rare Salvia ceratophylloides Ard.
3.2. Does the Within-Plant Variation of the Photosynthetic Performance, Morphological Traits and Metabolic Profiles Occured?
4. Materials and Methods
4.1. Species and Sites
4.2. Measurements and Samplings
4.3. Physiological Analysis
4.4. Morphological Analysis
4.5. VOC Analysis: HeadSpace/Solid-Phase Micro-Extraction (HS/SPME) GC-MS Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conti, F.; Manzi, A.; Pedrotti, F. Liste Rosse Regionali delle Piante d’Italia, WWF; Società Botanica Italiana: Camerino, Italy, 1997. [Google Scholar]
- Crisafulli, A.; Cannavò, S.; Maiorca, G.; Musarella, C.M.; Signorino, G.; Spampinato, G. Aggiornamenti floristici per la Calabria. Informatore Botanico Italiano 2010, 42, 431–442. [Google Scholar]
- Bonsignore, C.P.; Laface, V.L.A.; Vono, G.; Marullo, R.; Musarella, C.M.; Spampinato, G. Threats Posed to the Rediscovered and Rare Salvia ceratophylloides Ard. (Lamiaceae) by Borer and Seed Feeder Insect Species. Diversity 2021, 13, 33. [Google Scholar] [CrossRef]
- Laface, V.L.A.; Musarella, C.M.; Ortiz, A.C.; Canas, R.Q.; Cannavò, S.; Spampinato, G. Three New Alien Taxa for Europe and a Chorological Update on the Alien Vascular Flora of Calabria (Southern Italy). Plants 2020, 9, 1181. [Google Scholar] [CrossRef]
- Musarella, C.M.; Stinca, A. New data on the alien vascular flora of Calabria (Southern Italy). Annali di Botanica 2020, 10, 55–66. [Google Scholar] [CrossRef]
- Franks, S.J.; Weber, J.J.; Aitken, S.N. Evolutionary and plastic responses to climate change in terrestrial plant populations. Evol. Appl. 2014, 7, 123–139. [Google Scholar] [CrossRef]
- Noel, F.; Machon, N.; Porcher, E. No genetic diversity at molecular markers and strong phenotypic plasticity in populations of Ranunculus nodiflorus, an endangered plant species in France. Ann. Bot. 2007, 99, 1203–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerband, A.C.; Bialic-Murphy, L. Intraspecific variation in seedling drought tolerance and associated traits in a critically endangered, endemic Hawaiian shrub. Plant Ecol. Div. 2020. [Google Scholar] [CrossRef]
- Winn, A.A. Adaptation to fine-grained environmental variation: An analysis of within-individual leaf variation in an annual plant. Evolution 1996, 50, 1111–1118. [Google Scholar] [CrossRef]
- Winn, A.A. The contributions of programmed developmental change and phenotypic plasticity to within-individual variation in leaf traits in Dicerandra linearifolia. J. Evol. Biol. 1996, 9, 737–752. [Google Scholar] [CrossRef]
- de Kroon, H.; Huber, H.; Stuefer, J.F. A modular concept of phenotypic plasticity in plants. New Phytol. 2005, 166, 73–82. [Google Scholar] [CrossRef]
- Herrera, C.M. Multiplicity in Unity: Plant Subindividual Variation and Interactions with Animals; University of Chicago Press: Chicago, IL, USA, 2009. [Google Scholar]
- Herrera, C.M. The ecology of subindividual variability in plants: Patterns, processes, and prospects. Web Ecol. 2017, 17, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Osada, N.; Yasumura, Y.; Ishida, A. Leaf nitrogen distribution in relation to crown architecture in the tall canopy species, Fagus crenata. Oecologia 2014, 175, 1093–1106. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Bautista, A.; Valverde, P.L. Photosynthetically active radiation and carbon gain drives the southern orientation of Myrtillocactus geometrizans fruits. Plant Biol. 2017, 19, 279–285. [Google Scholar] [CrossRef]
- Hidalgo, J.; Rubio de Casas, R.; Muñoz, M.A. Environmental unpredictability and inbreeding depression select for mixed dispersal syndromes. BMC Evol. Biol. 2016, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.M.; Medrano, M.; Bazaga, P. Continuous within plant variation as a source of intraspecific functional diversity: Patterns, magnitude, and genetic correlates of leaf variability in Helleborus foetidus (Ranunculaceae). Am. J. Bot. 2015, 102, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C.; Liang, X.J. The mean and variability of a floral trait have opposing effects on fitness traits. Ann. Bot. 2016, 117, 421–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobral, M.; Guitián, J. Seed predators exert selection on the subindividual variation of seed size. Plant Biol. 2014, 16, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Takahashi, A. Effects of within plant variability in seed weight and tannin content on foraging behavior of seed consumers. Funct. Ecol. 2015, 29, 1513–1521. [Google Scholar] [CrossRef]
- Wetzel, W.C.; Kharouba, H.M. Variability in plant nutrients reduces insect herbivore performance. Nature 2016, 539, 425–427. [Google Scholar] [CrossRef]
- Wetzel, W.C.; Meek, M.H. Physical defenses and herbivory vary more within plants than among plants in the tropical understory shrub Piper polytrichum. Botany 2019, 97, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Di Iorio, A.; Vescio, R. The rediscovery of an endemic sage species in Calabria (Italy): An assessment of genetic diversity and structure in Salvia ceratophylloides Ard. (Lamiaceae). In Proceedings of the Congresso della Società Botanica Italiana, V International Plant Science Conference (IPSC), Fisciano, SA, Italy, 12–15 September 2018; p. 61, ISBN 978-88-85915-22-0. [Google Scholar]
- Aleric, K.M.; Kirkman, L.K. Growth and photosynthetic responses of the federally endangered shrub, Lindera melissifolia (Lauraceae), to varied light environments. Am. J. Bot. 2005, 92, 682–689. [Google Scholar] [CrossRef]
- Tkemaladze, G.S.; Makhashvili, K.A. Climate changes and photosynthesis. Ann. Agrar. Sci. 2016, 14, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Kattge, J. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa. Photosynthetica 2007, 45, 637–640. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- de la Riva, E.G.; Olmo, M. Leaf Mass per Area (LMA) and Its Relationship with Leaf Structure and Anatomy in 34 Mediterranean Woody Species along a Water Availability Gradient. PLoS ONE 2016, 11, e0148788. [Google Scholar] [CrossRef] [Green Version]
- Knight, C.A.; Ackerly, D.D. An ecological and evolutionary analysis of photosynthetic thermotolerance using the temperature dependent increase in fluorescence. Oecologia 2002, 130, 505–514. [Google Scholar] [CrossRef]
- Castrillo, M.; Vizcaino, D.; Moreno, E. Specific leaf mass, fresh: Dry weight ratio, sugar and protein contents in species of Lamiaceae from different light environments. Rev. Biol. Trop. 2005, 53, 23–28. [Google Scholar] [CrossRef]
- Goergen, P.C.H.; Lago, I. Performance of Chia on Different Sowing Dates: Characteristics of Growth Rate, Leaf Area Index, Shoot Dry Matter Partitioning and Grain Yield. J. Agric. Sci. 2019, 11, 252–263. [Google Scholar] [CrossRef]
- Mommer, L.; Wolters-Arts, M. Submergence-induced leaf acclimation in terrestrial species varying in flooding tolerance. New Phytol. 2007, 176, 337–345. [Google Scholar] [CrossRef]
- Paź-Dyderska, S.; Dyderski, M.K. Leaf Traits and Aboveground Biomass Variability of Forest Understory Herbaceous Plant Species. Ecosystems 2020, 23, 555–569. [Google Scholar] [CrossRef] [Green Version]
- Hedge, I.C. Notes on Salvia. Flora Europaea; Tutin, T.G., Ed.; Cambridge University Press: Cambridge, UK, 1972; Volume 3, pp. 188–192. [Google Scholar]
- Govaerts, R. World Checklist of Selected Plant Families Database in Access: 1-216203; The Board of Trustees of the Royal Botanic Gardens, Kew: London, UK, 2003. [Google Scholar]
- Nikolova, M.; Aneva, I. European Species of Genus Salvia: Distribution, Chemodiversity and Biological Activity. In Salvia Biotechnology; Georgiev, V., Pavlov, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–30. [Google Scholar]
- McKey, D. Adaptive patterns in alkaloid physiology. Am. Nat. 1974, 108, 305–320. [Google Scholar] [CrossRef]
- Meldau, S.; Erb, M.; Baldwin, I.T. Defence on demand: Mechanisms behind optimal defence patterns. Ann. Bot. 2012, 110, 1503–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medrano, H.; Tomás, M. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Schurr, U.; Walter, A.; Rascher, U. Functional dynamics of plant growth and photosynthesis–from steady-state to dynamics–from homogeneity to heterogeneity. Plant Cell Environ. 2006, 29, 340–352. [Google Scholar] [CrossRef]
- Lin, H.; Chen, Y. Stronger cooling effects of transpiration and leaf physical traits of plants from a hot dry habitat than from a hot wet habitat. Funct. Ecol. 2017, 31, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Leuzinger, S.; Korner, C. Tree species diversity affects canopy leaf temperatures in a mature temperate forest. Agr. For. Meteorol. 2007, 146, 29–37. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2018, 220, 692–702. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Frost, C.J.; Appel, H.M. Within-plant signalling via volatiles overcomes vascular constraints on systemic signalling and primes responses against herbivores. Ecol. Lett. 2007, 10, 490–498. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.R.; Rodriguez-Saona, L.E.; Frost, C. Herbivore-Induced Volatiles in the Perennial Shrub, Vaccinium corymbosum, and Their Role in Inter-branch Signaling. J. Chem. Ecol. 2009, 35, 163–175. [Google Scholar] [CrossRef]
- Slot, M.; Krause, G.H. Photosynthetic heat tolerance of shade and sun leaves of three tropical tree species. Photosynth. Res. 2019, 141, 119–130. [Google Scholar] [CrossRef]
- de Casas, R.R.; Vargas, P. Sun and shade leaves of Olea europaea respond differently to plant size, light availability and genetic variation. Funct. Ecol. 2011, 25, 802–812. [Google Scholar] [CrossRef]
- Girón-Calva, P.S.; Li, T. A Role for Volatiles in Intra- and Inter-Plant Interactions in Birch. J. Chem. Ecol. 2014, 40, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Alberto, F.J.; Aitken, S.N. Potential for evolutionary responses to climate change–evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [Green Version]
- Aitken, S.N.; Whitlock, M.C. Assisted gene flow to facilitate local adaptation to climate change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 367–388. [Google Scholar] [CrossRef]
- Sobral, M.; Guitiàn, J.; Guitiàn, P. Selective larchure along a Latitudinal Gradient Affects Subindividual Variation in Plants. PLoS ONE 2013, 8, 74356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruschi, P.; Grossoni, P.; Bussotti, F. Within- and among-tree variation in leaf morphology of Quercus petraea (Matt.) Liebl. natural populations. Trees 2003, 17, 164–172. [Google Scholar] [CrossRef]
- Opedal, Ø.H.; Armbruster, W.S.; Graae, B.J. Linking small-scale topography with microclimate, plant species diversity and intra-specific trait variation in an alpine landscape. Plant Ecol. Div. 2015, 8, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, U.; Valladares, F. Photosynthetic acclimation to simultaneous and interacting environmental stresses along natural light gradients: Optimality and constraints. Plant Biol. 2004, 6, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Meier, I.C.; Leuschner, C. Genotypic variation and phenotypic plasticity in the drought response of fine roots of European beech. Tree Physiol. 2008, 28, 297–309. [Google Scholar] [CrossRef]
- Marchiori, P.E.R.; Machado, E.C. Physiological Plasticity Is Important for Maintaining Sugarcane Growth under Water Deficit. Front. Plant Sci. 2017, 8, 2148. [Google Scholar] [CrossRef] [Green Version]
- Puglielli, G.; Catoni, R.L. Short-term physiological plasticity: Trade-off between drought and recovery responses in three Mediterranean Cistus species. Ecol. Evol. 2017, 7, 10880–10889. [Google Scholar] [CrossRef]
- Valladares, F.; Martinez-Ferri, E. Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: A conservative resource-use strategy? New Phytol. 2000, 148, 79–91. [Google Scholar] [CrossRef] [Green Version]
- ARSSA. I Suoli Della Calabria—Carta dei Suoli in Scala 1:250.000 Della Regione Calabria; Rubettino Editore: Soveria Mannelli, Italy, 2003; p. 387. [Google Scholar]
- Pignatti, S. Flora d’Italia. In 4 Volumi. Volume 3: Flora d’Italia & Flora Digitale, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2018; ISBN 978-88-506-5244-0. [Google Scholar]
- Rivas-Martínez, S. Global Bioclimatics (Clasificación Bioclimática de la Tierra). Available online: http://www.globalbioclimatics.org/book/publications.htm (accessed on 26 September 2019).
- Doussan, C.; Pagès, L.; Pierret, A. Soil exploration and resource acquisition by plant roots: An architectural and modelling point of view. Agronomie 2003, 23, 419–431. [Google Scholar] [CrossRef]
- Jonckheere, I.; Nackaerts, K. A fractal dimension-based modelling approach for studying the effect of leaf distribution on LAI retrieval in forest canopies. Ecol. Model 2006, 97, 179–195. [Google Scholar] [CrossRef]
- Quero, J.L.; Villar, R.; Marañon, T.; Zamora, R. Interactions of drought and shade effects on seedlings of four Quercus species: Physiological and structural leaf responses. New Phytol. 2006, 170, 819–834. [Google Scholar] [CrossRef] [Green Version]
- Ruíz-Robleto, J.; Villar, R. Relative growth rate and biomass allocation in ten woody species with different leaf longevity using phylogenetic independent contrasts (PICs). Plant Biol. 2005, 7, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, J.H.C.; Pérez-Hargundeguy, N. Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents. New Phytol. 1999, 143, 191–200. [Google Scholar] [CrossRef]
- Ryser, P.; Aeschlimann, U. Proportional dry-mass content as an underlying trait for the variation in relative growth rate among 22 Eurasian populations of Dactylis glomerata s.l. Funct. Ecol. 1999, 13, 473–482. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Diaz, S. New handbook for standardized measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Saura-Mas, S.; Shipley, B.; Lloret, F. Relationship between post-fire regeneration and leaf economics spectrum in Mediterranean woody species. Funct. Ecol. 2009, 23, 103–110. [Google Scholar] [CrossRef]
- Cruz, A.F.S.; Adiel, F. Stress index, water potentials and leaf succulence in cauliflower cultivated hydroponically with brackish water. Rev. Bras. Eng. Agrícola Amb. 2018, 22, 622–627. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Patti, G.J. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopka, J.; Schauer, N. GMD@ CSB. DB: The Golm metabolome database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Horai, H.; Arita, M. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Lobo, F.; de Barros, M.P. Fitting net photosynthetic light-response curves with Microsoft Excel—A critical look at the models. Photosynthetica 2013, 51, 445–456. [Google Scholar] [CrossRef]
- Press, W.H.; Teukosky, S.A. Numerical Recipes in C. The Art of Scientific Computing, 2nd ed.; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Larcher, W. Physiological Plant Ecology. Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Heidelberg, Germany, 2003. [Google Scholar]
- Duursma, R.A.; Falster, D.S. Leaf mass per area, not total leaf area, drives differences in above-ground biomass distribution among woody plant functional types. New Phytol. 2016, 212, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Martins, F.; Oliveira, İ. Leaf morpho-physiological dynamics in Salvia officinalis L. var. purpurascens. Turk. J. Biol. 2017, 41, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Perrino, E.V.; Wagensommer, R.P. Crop wild relatives (CWR) priority in Italy: Distribution, ecology, in situ and ex situ conservation and expected actions. Sustainability 2021, 13, 1682. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Site (Sit) | ||||
|---|---|---|---|---|
| Leaf Type (LT) | Mo | Pu | Leaf Type Average | |
| Icomp [µmol(photon) m−2 s−1] | P | 8b | 26a | 18x |
| S | 22a | 22a | 22x | |
| Site average | 16A | 23A | ||
| Imax [µmol(photon) m−2 s−1] | P | 312b | 310b | 311y |
| S | 839a | 725a | 766x | |
| Site average | 655A | 577A | ||
| Isat [µmol(photon) m−2 s−1] | P | 1027b | 818b | 911y |
| S | 1559a | 1588a | 1578 x | |
| Site average | 1323A | 1313A | ||
| PN(Imax) [µmol(CO2) m−2 s−1] | P | 6.85b | 2.17b | 4.25y |
| S | 19.70a | 6.51b | 11.22x | |
| Site average | 14.00A | 4.96B | ||
| RD [µmol(CO2) m−2 s−1] | P | 0.53b | 1.07ab | 0.69 y |
| S | 1.40a | 1.09ab | 1.19 x | |
| Site average | 1.01A | 1.08A | ||
| ϕ(I0-comp) [µmol(CO2) µmol(photon) −1] | P | 0.025b | 0.0095c | 0.016y |
| S | 0.046a | 0.021b | 0.030x | |
| Site average | 0.037A | 0.016B | ||
| Sites (Sit) | ||||
|---|---|---|---|---|
| Leaf Type (LT) | Mo | Pu | Leaf Type Average | |
| Stomatal conductance (mol H2O m−2 s−1) | P | 0.032b | 0.016b | 0.023y |
| S | 0.113a | 0.029b | 0.059x | |
| Site average | 0.077A | 0.025B | ||
| Transpiration rate (mol H2O m−2 s−1) | P | 0.87b | 0.55b | 0.69y |
| S | 2.59a | 0.92b | 1.52x | |
| Site average | 1.82A | 0.79B | ||
| Sites (Sit) | ||||
|---|---|---|---|---|
| Leaf Type (LT) | Mo | Pu | Leaf Type Average | |
| Stomatal conductance (mol H2O m−2 s−1) | P | 0.032b | 0.016b | 0.023y |
| S | 0.107a | 0.032b | 0.059x | |
| Site average | 0.074A | 0.026B | ||
| Transpiration rate (mol H2O m−2 s−1) | P | 0.83b | 0.55b | 0.67y |
| S | 2.44a | 1.03b | 1.53x | |
| Site average | 1.72A | 0.86B | ||
| Sites (Sit) | ||||
|---|---|---|---|---|
| Leaf Type (LT) | Mo | Pu | Leaf Type Average | |
| Leaf fresh weight [g leaf−1] | P | 1.33a | 1.38a | 1.36x |
| S | 1.40a | 0.43b | 0.78y | |
| Site average | 1.37A | 0.77B | ||
| Leaf dry weight [g leaf−] | P | 0.23a | 0.22b | 0.22x |
| S | 0.27a | 0.12b | 0.17x | |
| Site average | 0.25A | 0.16B | ||
| Leaf area [cm2] | P | 41.4ab | 54.4a | 48.6x |
| S | 43.6ab | 19.9b | 28.3y | |
| Site average | 32.2A | 42.6A | ||
| Leaf mass x area [g m−2] | P | 55.4a | 44.1a | 49.1x |
| S | 62.3a | 61.6a | 61.8x | |
| Site average | 59.2A | 55.3A | ||
| Leaf dry content [g dry weight g−1 fresh weight] | P | 0.17a | 0.18a | 0.18x |
| S | 0.19a | 0.28a | 0.25x | |
| Site average | 0.18A | 0.24A | ||
| Leaf water content [g H2O cm−2 leaf area] | P | 0.027a | 0.020a | 0.023x |
| S | 0.025b | 0.016b | 0.019y | |
| Site average | 0.026A | 0.017B | ||
| Fractal dimension | P | 1.67a | 1.73a | 1.70x |
| S | 1.51b | 1.65b | 1.56y | |
| Site average | 1.69A | 1.59A | ||
| Compound | # Statistics | Sessile | Petiolate | |||
|---|---|---|---|---|---|---|
| Pu | Mo | Pu | Mo | |||
| 1 | p-Cymene | LT 7.78 * Sit 14.16 ** LT × Sit 0.21 NS | 195,813 | 82,392 | 108,569 | 19,607 |
| 2 | Pinocarvone | LT 0.04 NS Sit 6.57 * LT × Sit 0.07 NS | 2406 | 987 | 2444 | 696 |
| 3 | Sabinene | LT 11.80 ** Sit 0.34 NS LT × Sit 1.70 NS | 554,775 | 873,306 | 195,242 | 73,210 |
| 4 | Terpinolene | LT 12.40 ** Sit 0.40 NS LT × Sit 3.09 NS | 128,554 | 218,320 | 62,198 | 19,784 |
| 5 | β-Pinene | LT 7.30 * Sit 0.47 NS LT × Sit 1.73 NS | 92,968 | 150,052 | 53,391 | 35,502 |
| 6 | γ-Terpinene | LT 5.40 * Sit 0.19 NS LT × Sit 0.04 NS | 16,341 | 13,366 | 4610 | 3508 |
| 7 | α-Terpineol |
LT 8.13 * Sit 12.91 ** LT × Sit 9.04 * | 10,220b | 80,003a | 11,854b | 18,054b |
| 8 | D-Germacrene | LT 0.11 NS Sit 3.47 NS LT × Sit 4.22 * | 2554a | 169b | 1102a | 1218a |
| 9 | α-Copaene | LT 0.62 NS Sit 11.05 * LT × Sit 0.67 NS | 3517 | 601 | 2385 | 625 |
| 10 | α-Cubebene | LT 8.21 * Sit 19.35 ** LT × Sit 1.02 | 4,460,201 | 1,371,705 | 2,247,264 | 312,465 |
| 11 | α-Muurolene | LT 9.49 * Sit 0.56 NS LT × Sit 0.99 NS | 14,382 | 15,236 | 7105 | 1038 |
| 12 | Isovaleraldehyde | LT 6.10 * Sit 0.52 NS LT × Sit 0.52 NS | 81,876,770 | 46,391,341 | 3,426,789 | 3,466,464 |
| 13 | 5-Methylheptan-3-one | LT 5.70 * Sit 0.21 NS LT × Sit 0.08 NS | 7776 | 8204 | 1578 | 3291 |
| 14 | Pentan-3-one | LT 7.73 * Sit 2.44 NS LT × Sit 1.20 NS | 321,989 | 649,080 | 114,170 | 171,753 |
| 15 | β-tujone |
LT 17.37 ** Sit 6.21 * LT × Sit 12.54 ** | 65,370b | 168,599a | 54,660b | 36,692b |
| 16 | (3z)-3-Hexenyl acetate | LT 3.58 NS Sit 5.09 NS LT × Sit 5.46 * | 1253a | 0b | 99ab | 122ab |
| 17 | Dimethyl Sulfide | LT 23.77 ** Sit 5.34 * LT × Sit 1.40 NS | 29,866,633 | 54,386,837 | 3,926,181 | 11,857,751 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vescio, R.; Abenavoli, M.R.; Araniti, F.; Musarella, C.M.; Sofo, A.; Laface, V.L.A.; Spampinato, G.; Sorgonà, A. The Assessment and the Within-Plant Variation of the Morpho-Physiological Traits and VOCs Profile in Endemic and Rare Salvia ceratophylloides Ard. (Lamiaceae). Plants 2021, 10, 474. https://doi.org/10.3390/plants10030474
Vescio R, Abenavoli MR, Araniti F, Musarella CM, Sofo A, Laface VLA, Spampinato G, Sorgonà A. The Assessment and the Within-Plant Variation of the Morpho-Physiological Traits and VOCs Profile in Endemic and Rare Salvia ceratophylloides Ard. (Lamiaceae). Plants. 2021; 10(3):474. https://doi.org/10.3390/plants10030474
Chicago/Turabian StyleVescio, Rosa, Maria Rosa Abenavoli, Fabrizio Araniti, Carmelo Maria Musarella, Adriano Sofo, Valentina Lucia Astrid Laface, Giovanni Spampinato, and Agostino Sorgonà. 2021. "The Assessment and the Within-Plant Variation of the Morpho-Physiological Traits and VOCs Profile in Endemic and Rare Salvia ceratophylloides Ard. (Lamiaceae)" Plants 10, no. 3: 474. https://doi.org/10.3390/plants10030474