Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Soil Analysis
2.2. Phytochemical Assay
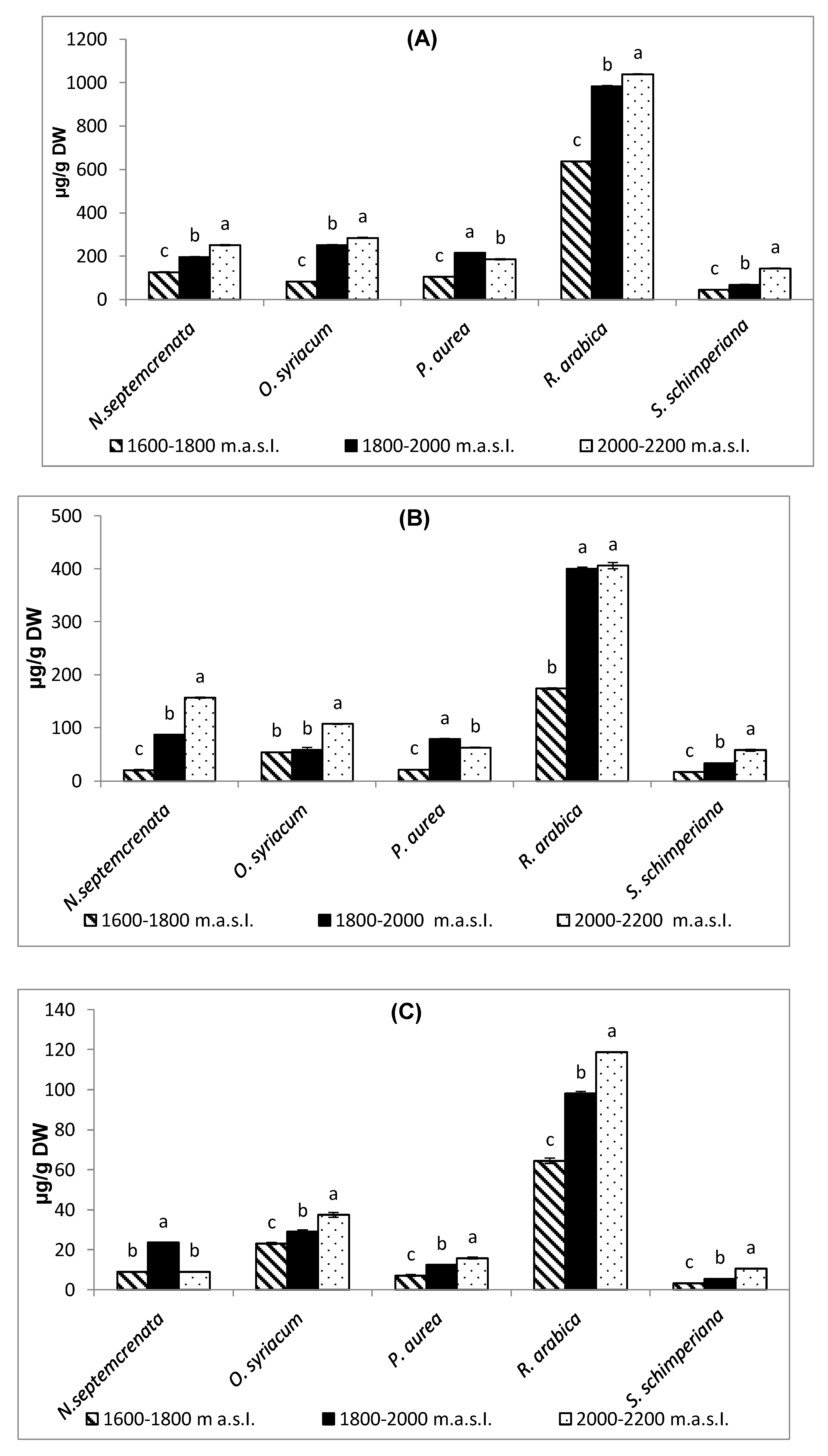
2.2.1. Change in the Photosynthetic Pigments
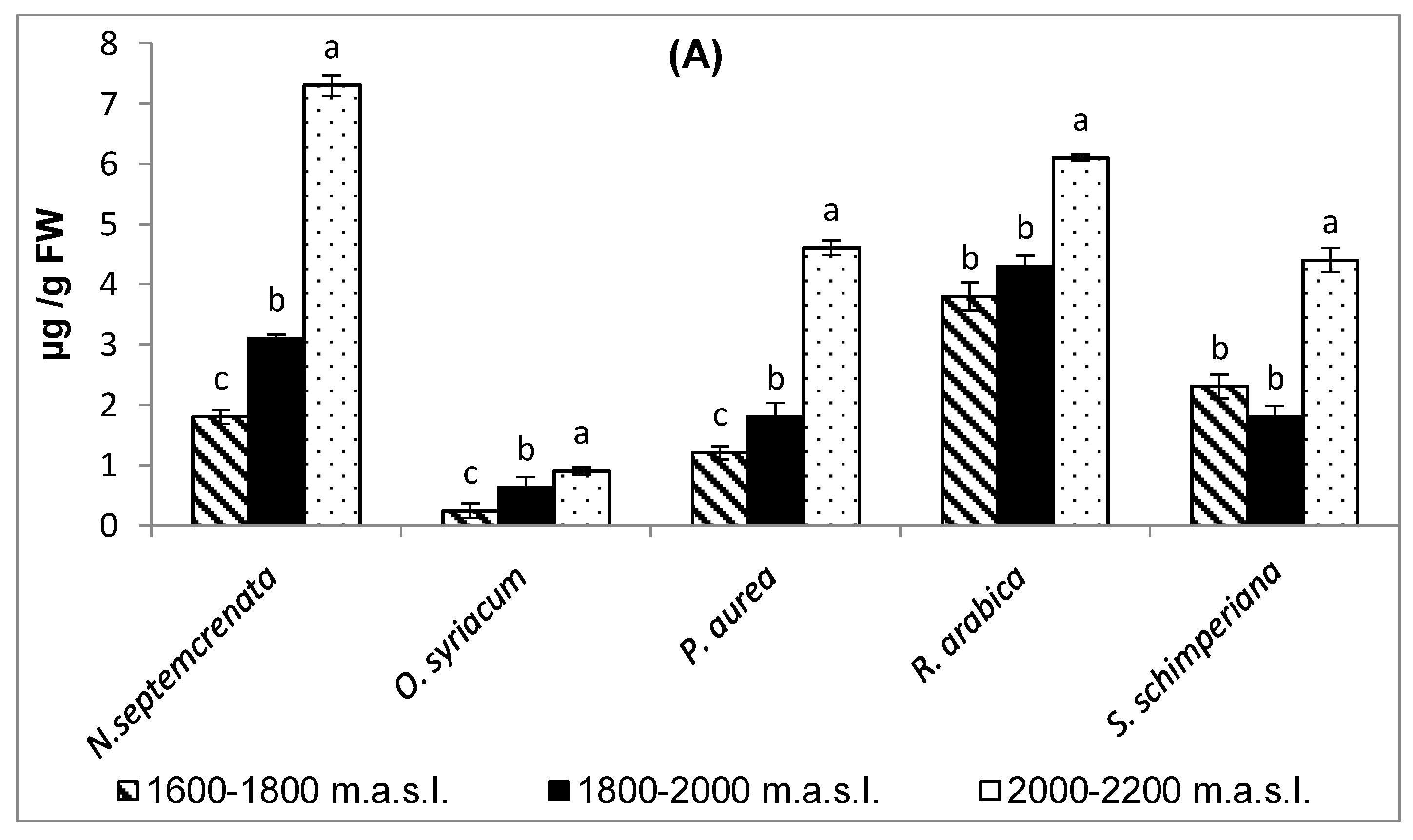
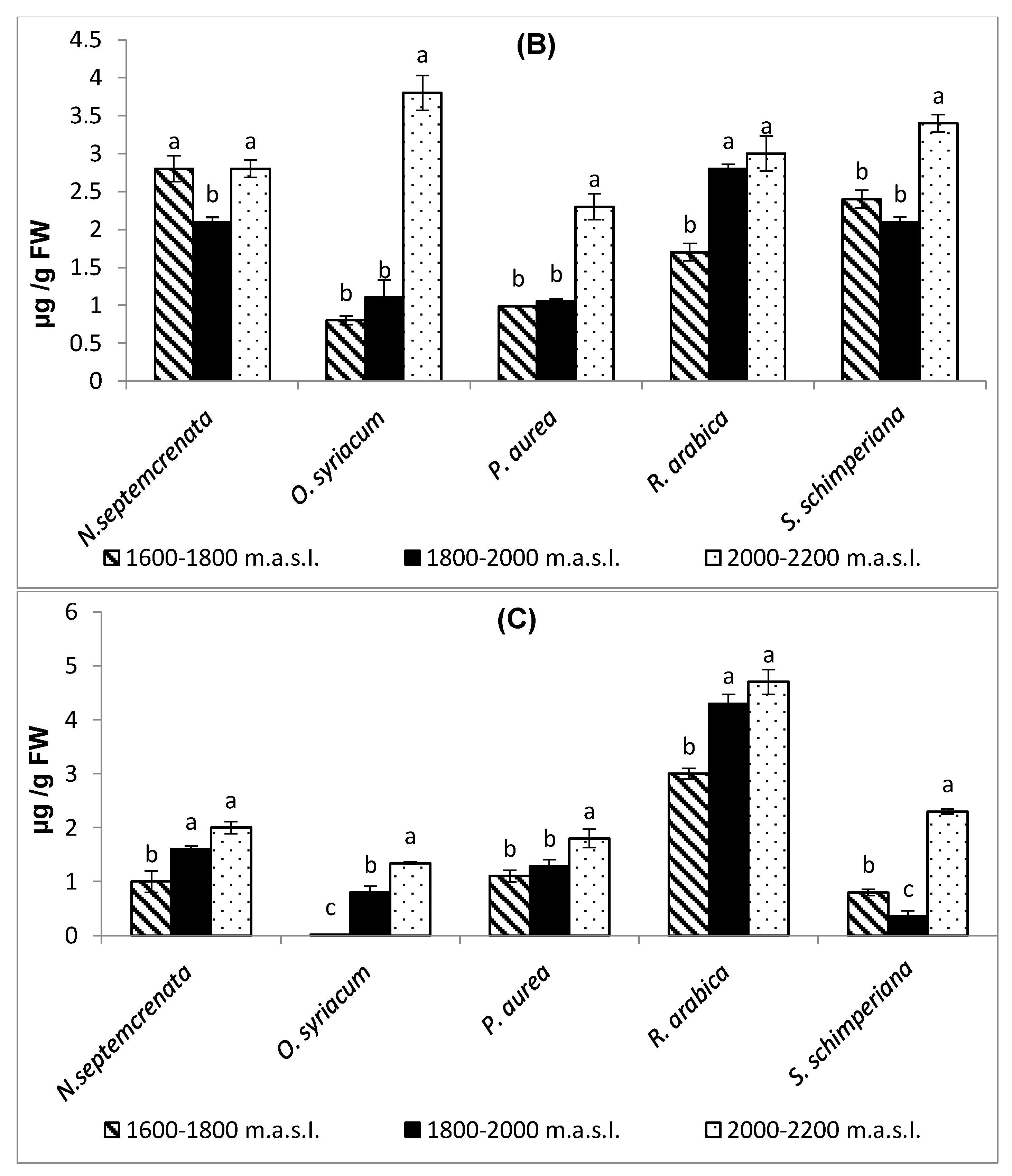
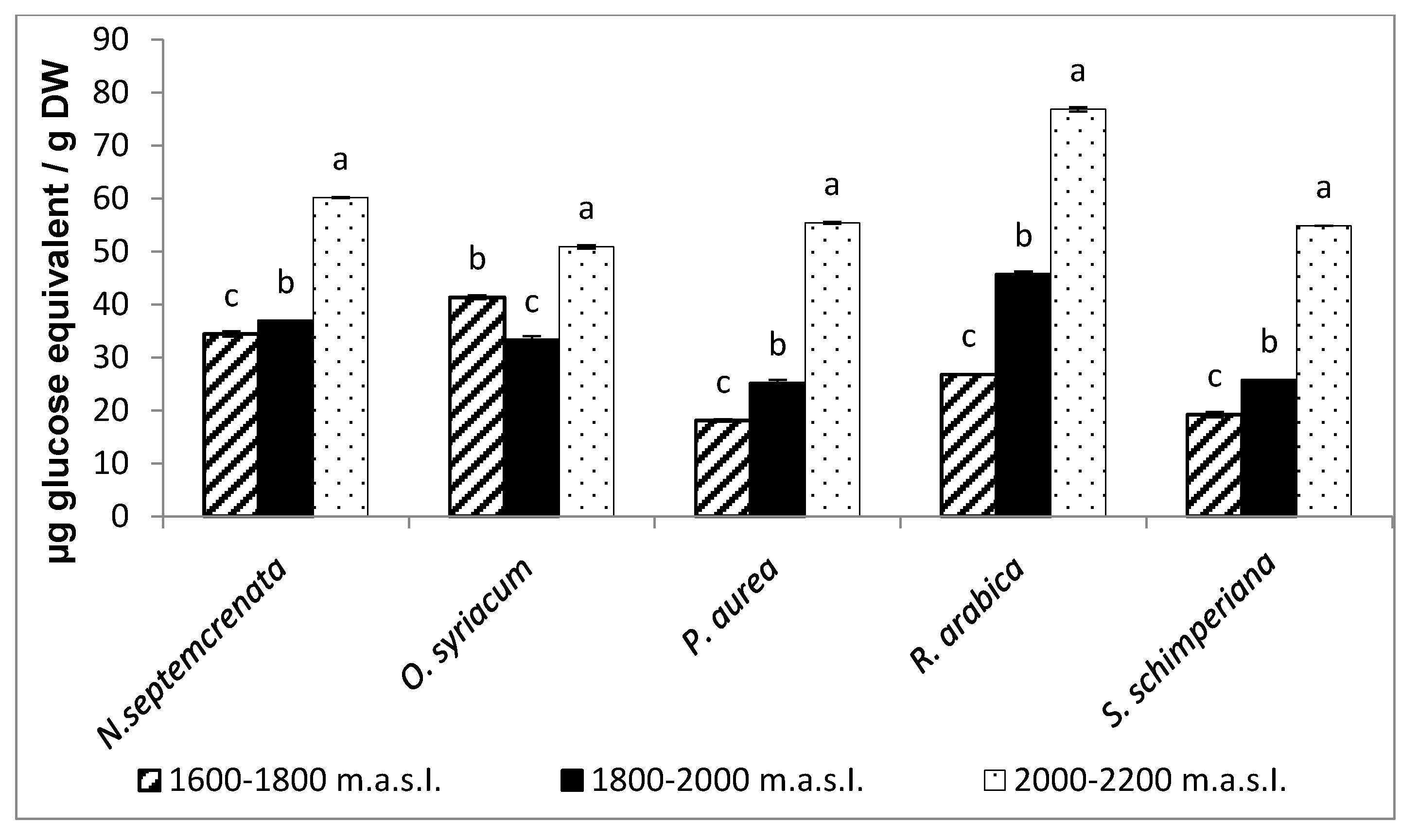
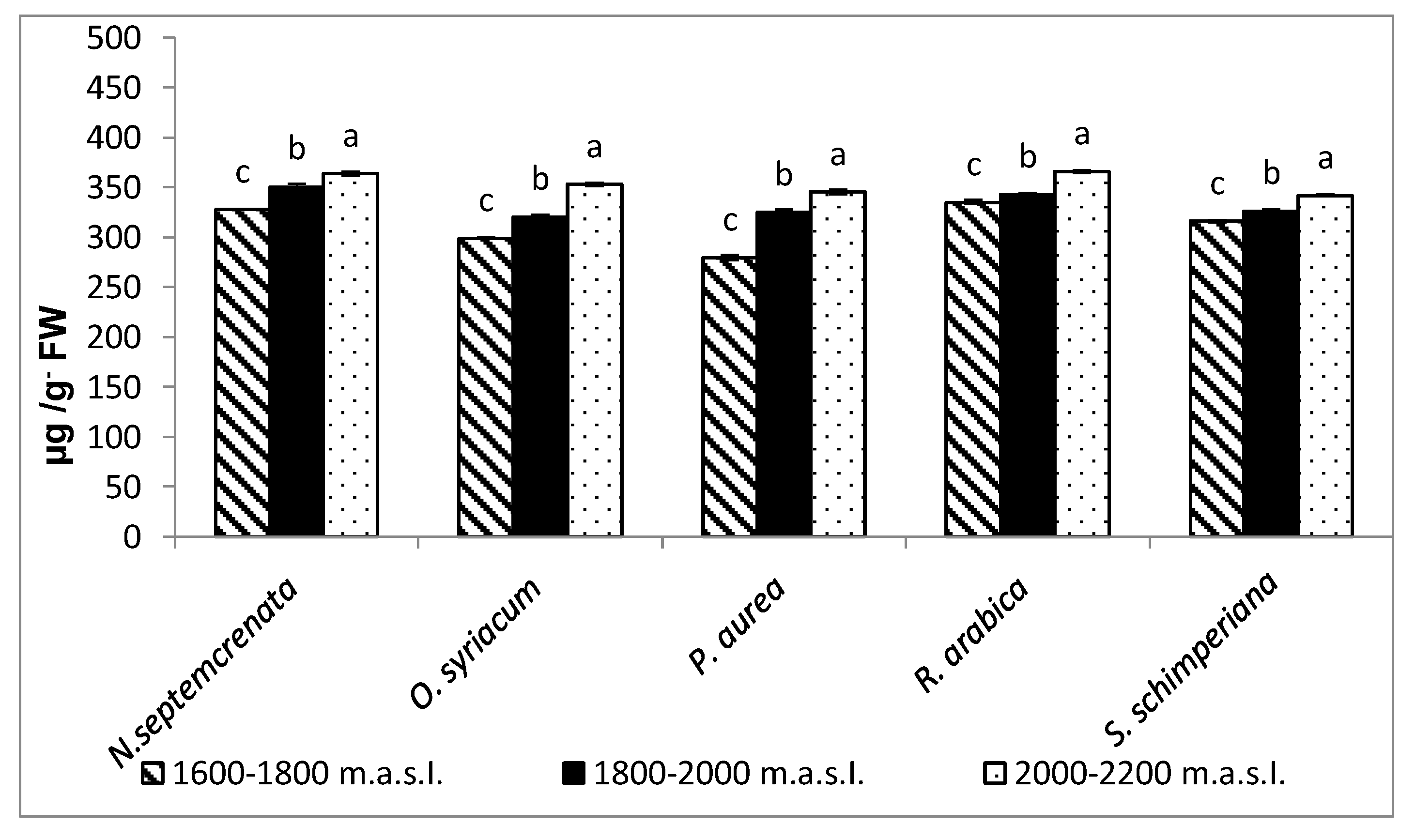
2.2.2. Changes in the Content of Major Secondary Metabolites
2.2.3. Changes in the Content of Total Soluble Sugars
2.3. Biochemical Analysis
2.3.1. Changes in the Content of Total Soluble Proteins
2.3.2. Determination of Protein Banding Pattern
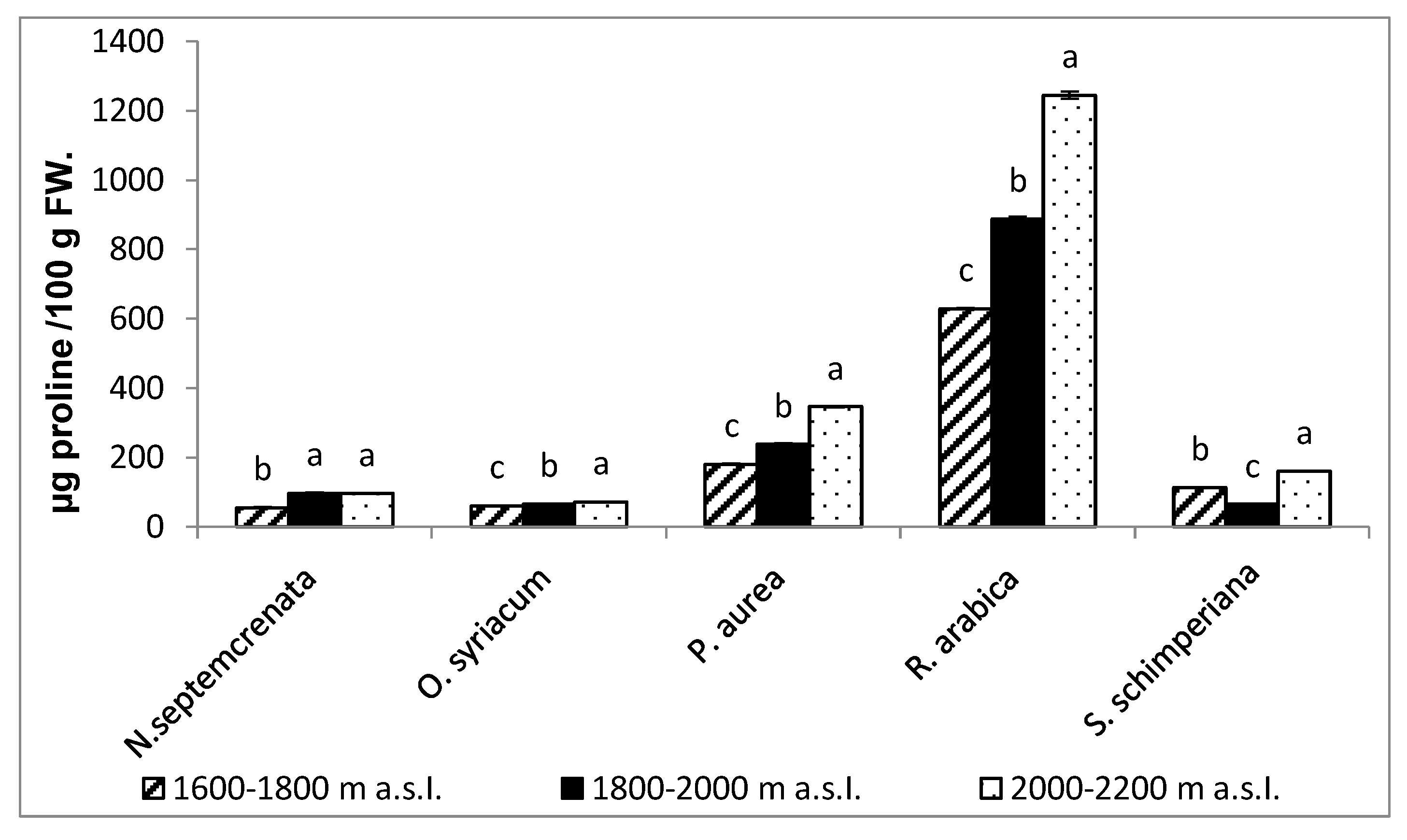
2.3.3. Changes in the Proline Content
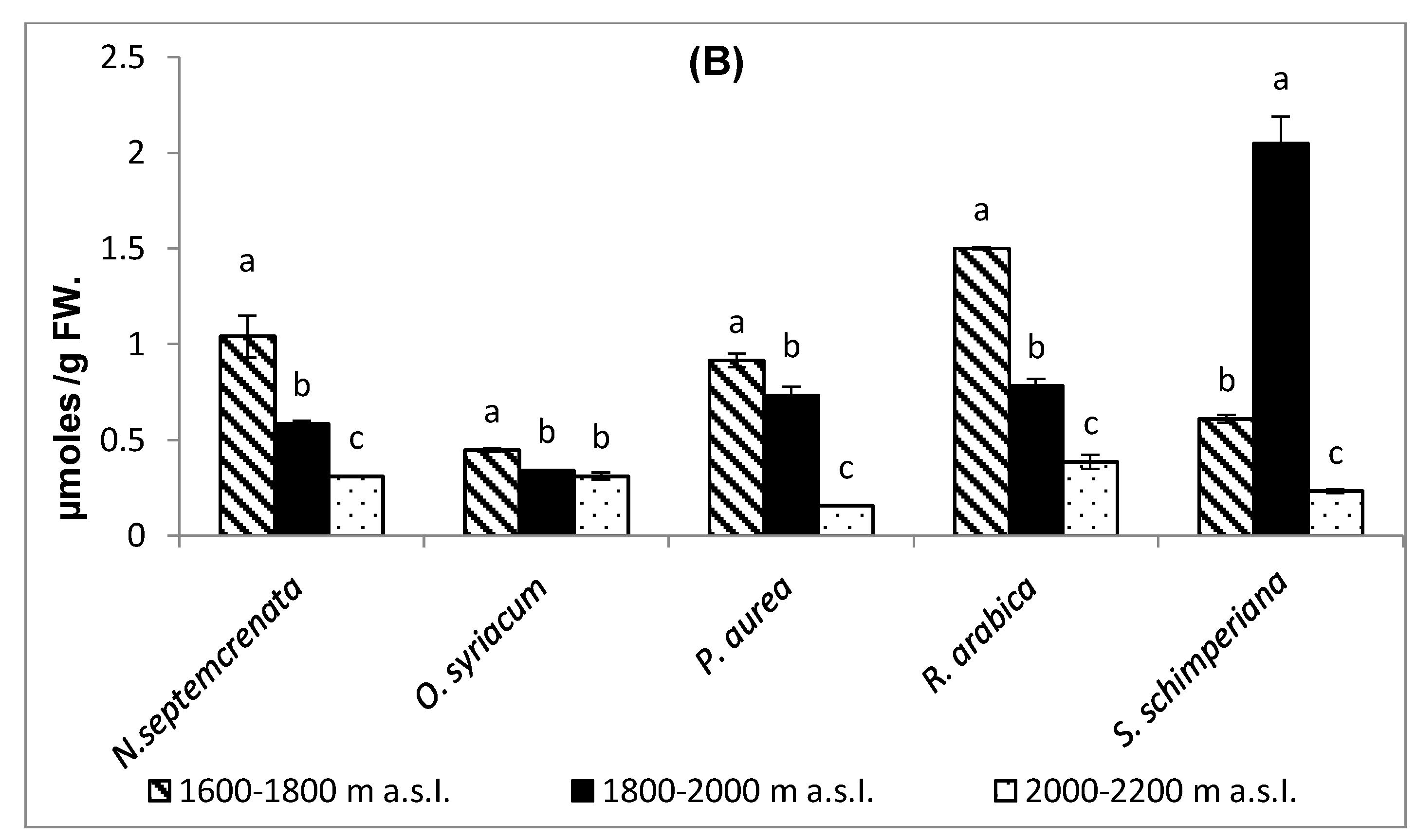
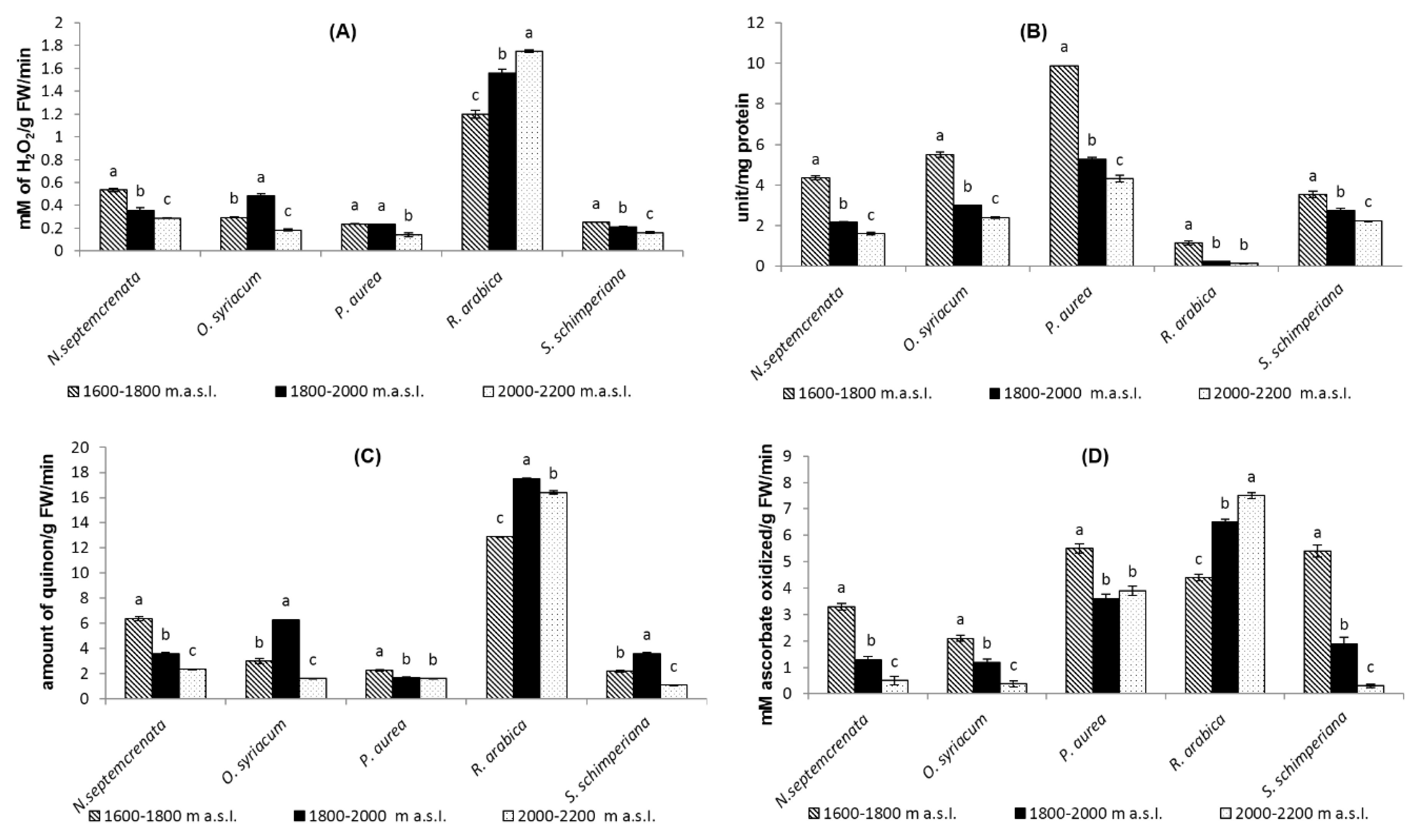
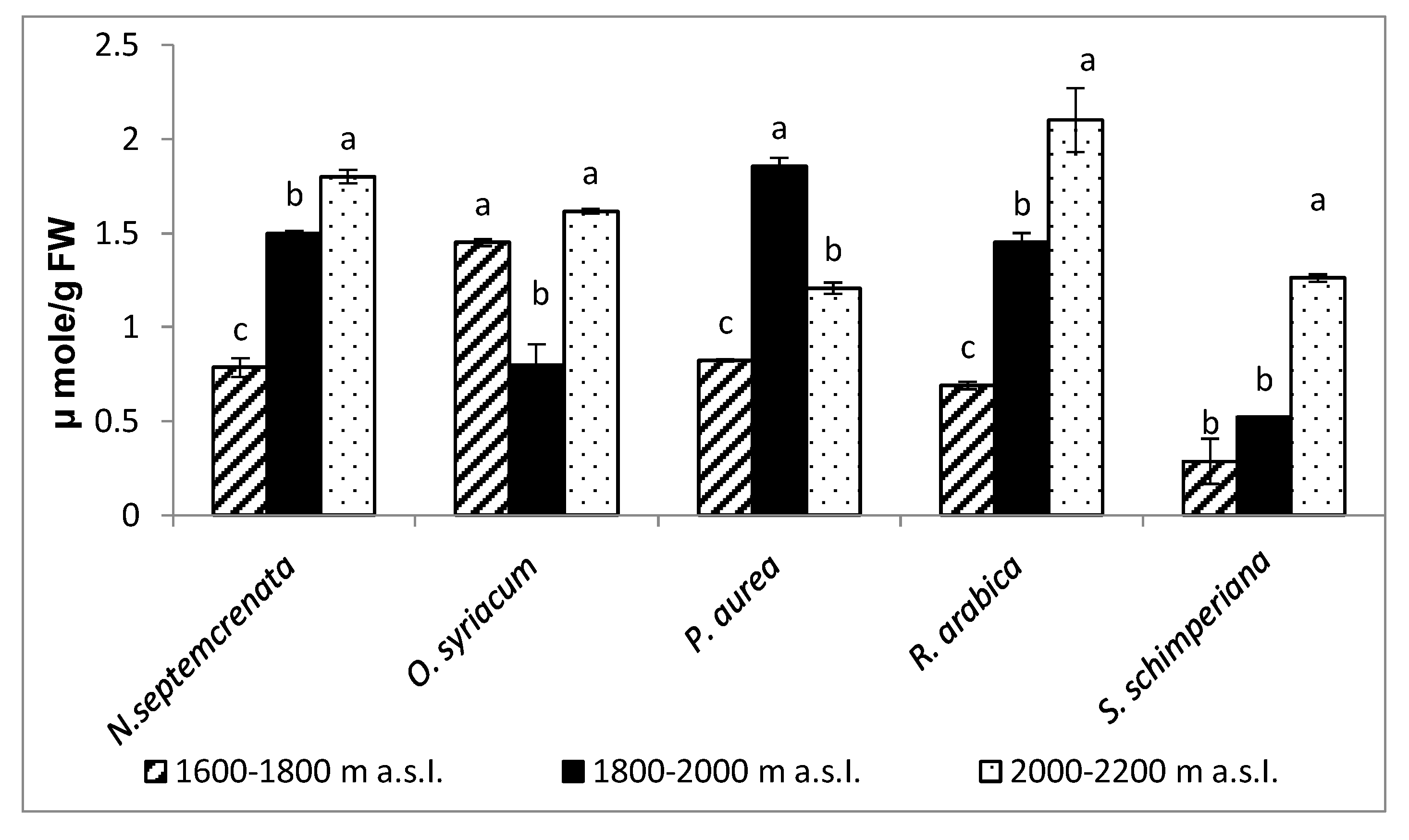
2.3.4. Changes in Total Antioxidant Capacity and Malondialdehyde (MDA) Contents
2.3.5. Change in Activity Level of Some Antioxidant Enzymes
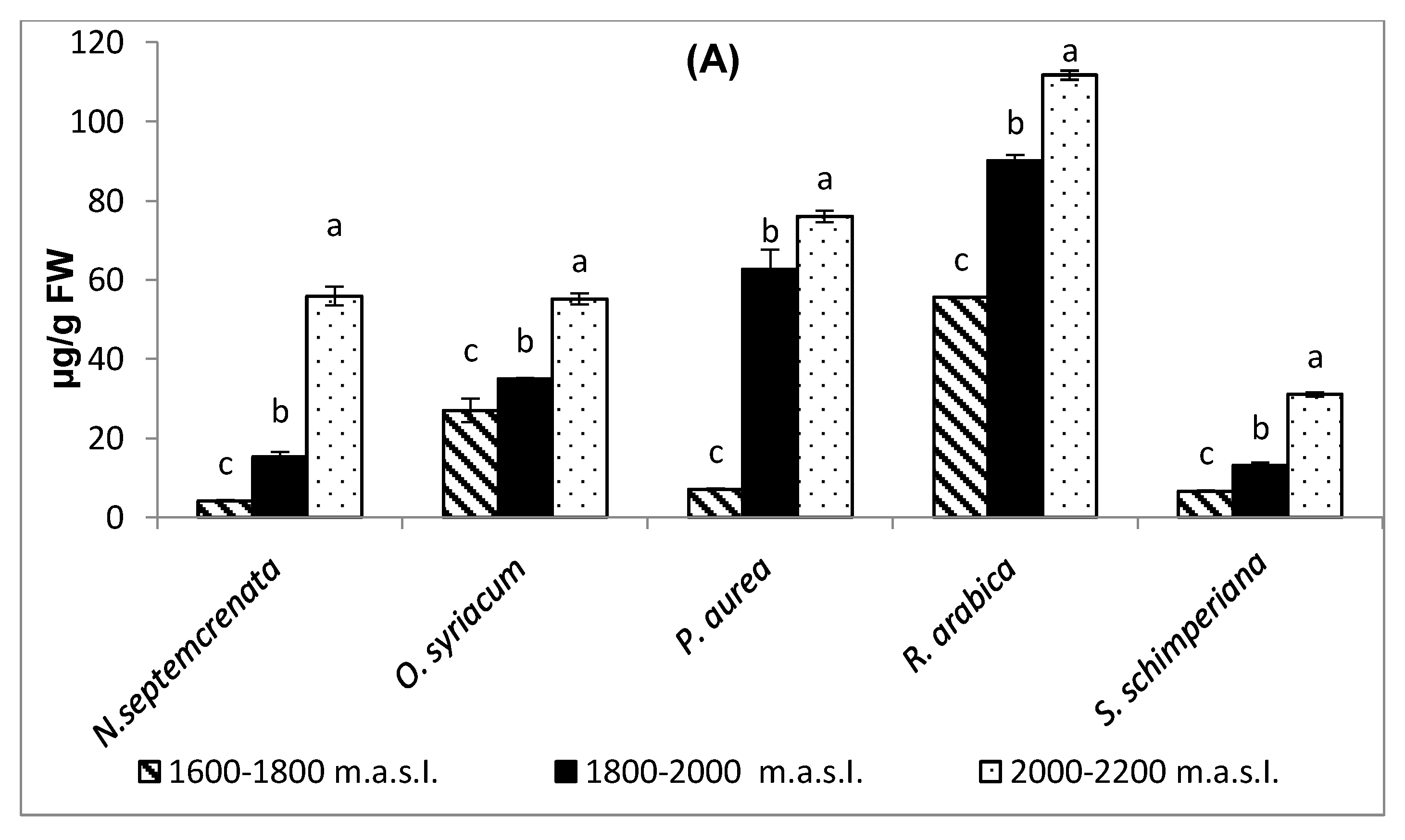
2.3.6. Change in Ascorbic Acid Contents
3. Discussion
4. Materials and Methods
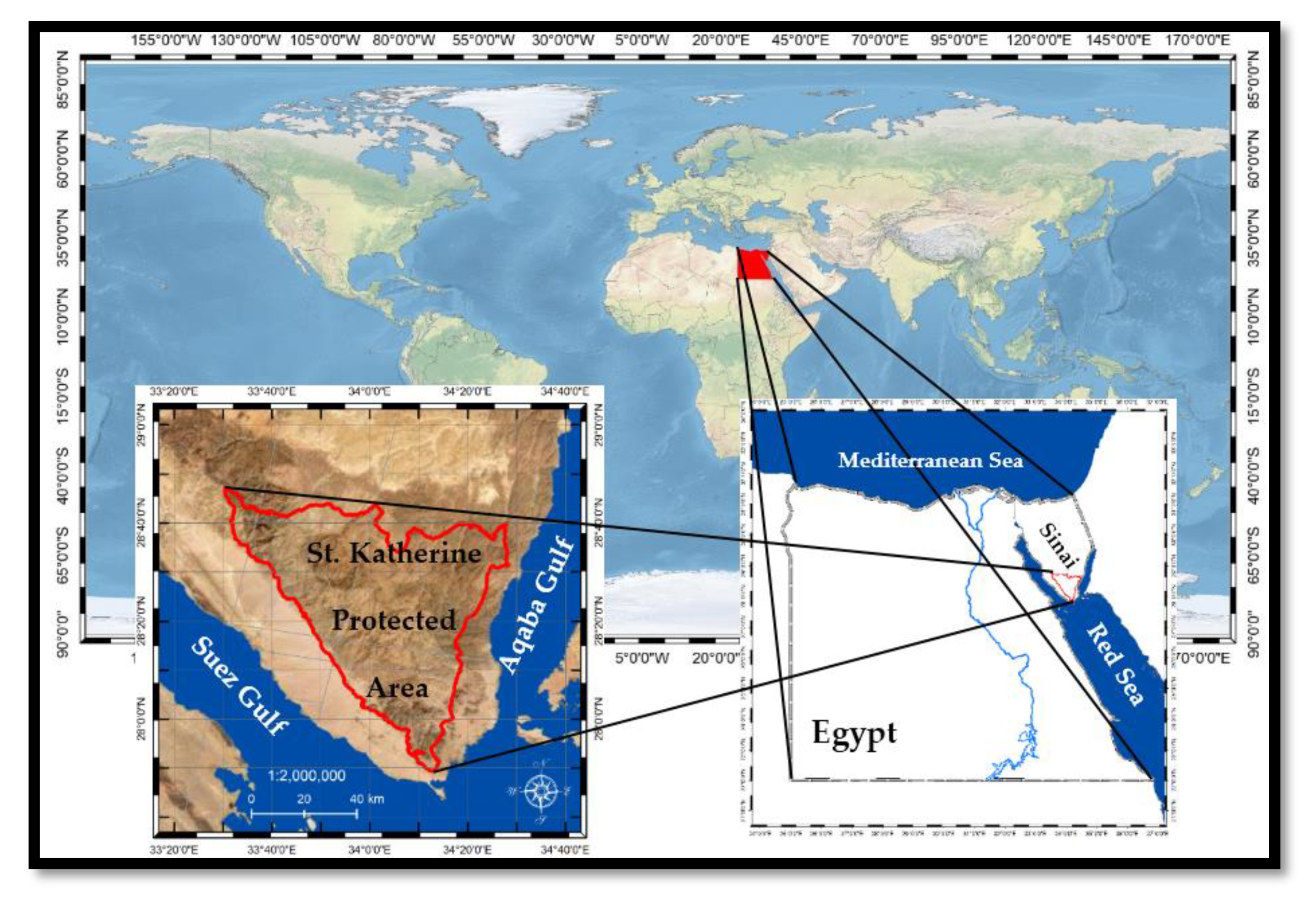
4.1. Study Area
4.2. Target Species
4.3. Soil Analysis
4.4. Phytochemical Assay
4.4.1. Extraction and Estimation of Photosynthetic Pigments
4.4.2. Extraction and Estimation of Total Soluble Sugars
4.4.3. Extraction and Estimation of Total Phenolic Content
4.4.4. Extraction and Estimation of Total Flavonoids Content
4.4.5. Extraction and Estimation of Tannins Content
4.5. Biochemical Assay
4.5.1. Extraction and Estimation of Total Soluble Proteins
4.5.2. Determination of Protein Banding Pattern
4.5.3. Estimation of Proline
4.5.4. Extraction and Estimation of Malondialdehyde
4.6. Enzyme Extraction and Assays
Extraction and Assaying Activity of Certain Enzymes
4.7. Determination of Ascorbic Acid
4.8. Determination of Total Antioxidant Capacity
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williams, S.E.; Williams, Y.M.; VanDerWal, J.; Isaac, J.L.; Shoo, L.P.; Johnson, C.N. Ecological specialization and population size in a biodiversity hotspot: How rare species avoid extinction. Proc. Natl. Acad. Sci. USA 2009, 106, 19737–19741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhaithloul, H.A.S. Impact of Combined Heat and Drought Stress on the Potential Growth Responses of the Desert Grass Artemisia sieberi alba: Relation to Biochemical and Molecular Adaptation. Plants 2019, 8, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, N.; Ali, Q.; Ali, S.; Javed, M.T.; Zulqurnain Haider, M.; Perveen, R.; Shahid, M.R.; Rizwan, M.; Abdel-Daim, M.M.; Elkelish, A.; et al. Use of Nitric Oxide and Hydrogen Peroxide for Better Yield of Wheat (Triticum aestivum L.) under Water Deficit Conditions: Growth, Osmoregulation, and Antioxidative Defense Mechanism. Plants 2020, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, G.; Gottfried, M.; Pauli, H. Climate effects on mountain plants. Nature 1994, 369, 448. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Almoneafy, A.; Mahmoud, A.; Elkelish, A.; Arnao, M.B.; Li, L.; Ai, S. Melatonin and Its Protective Role against Biotic Stress Impacts on Plants. Biomolecules 2020, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- El-Keblawy, A.A.; Khedr, A.-H.A. Population structure and ecological role of Moringa peregrina (Forssk.) Fiori. at its northwestern range edge in the Hajar Mountains. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2017, 151, 29–38. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Alhaithloul, H.A.S.; Qari, S.H.; Soliman, M.H.; Hasanuzzaman, M. Pretreatment with Trichoderma harzianum alleviates waterlogging-induced growth alterations in tomato seedlings by modulating physiological, biochemical, and molecular mechanisms. Environ. Exp. Bot. 2019, 103946. [Google Scholar] [CrossRef]
- Moustafa, A.R.A.; Klopatek, J.M. Vegetation and landforms of the Saint Catherine area, southern Sinai, Egypt. J. Arid Environ. 1995, 30, 385–395. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.; Nada, A.A.; O’Donovan, A.; Kumar Thakur, V.; Elkelish, A. Mycogenic Silver Nanoparticles from Endophytic Trichoderma atroviride with Antimicrobial Activity. J. Renew. Mater. 2019, 7, 171–185. [Google Scholar] [CrossRef]
- Ayyad, M.A.; Fakhry, A.M.; Moustafa, A.-R.A. Plant biodiversity in the Saint Catherine area of the Sinai peninsula, Egypt. Biodivers. Conserv. 2000, 9, 265–281. [Google Scholar] [CrossRef]
- Tackholm, V. Students’ flora of Egypt 1974, 2nd ed.; Cairo University Publishing: Beirut, Lebanon, 1974; 888p. [Google Scholar]
- IUCN. The 100 Most Threatened Species. Are They Priceless or Worthless? Available online: https://www.iucn.org/content/100-most-threatened-species-are-they-priceless-or-worthless (accessed on 21 March 2020).
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [Green Version]
- Shaltout, K.H.; El-Hamdi, K.H.; El-Masry, S.A.; Eid, E.M. Bedouin farms in the Saint Katherine mountainous area (South Sinai, Egypt). J. Mt. Sci. 2019, 16, 2232–2242. [Google Scholar] [CrossRef]
- Boulos, L. Flora of Egypt. Checklist; Al-Hadara Publishing: Cairo, Egypt, 1995. [Google Scholar]
- El-Demerdash, M. The ex Situ Conservation Technical Report on Propagation of Medicinal Plants; MPCP, The Egyptian Environmental Affairs Agency (EEAA): Cairo, Egypt. Available online: http://www.eeaa.gov.eg/en-us/topics/nature/biodiversity/plantgeneticresources/floraconservation.aspx (accessed on 1 April 2020).
- Shaltout, K.H.; Ahmed, D.A.; Shabana, H.A. Population structure and dynamics of the endemic species Phlomis aurea Decne in different habitats in southern Sinai Peninsula, Egypt. Glob. Ecol. Conserv. 2015, 4, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Roupioz, L.; Jia, L.; Nerry, F.; Menenti, M. Estimation of Daily Solar Radiation Budget at Kilometer Resolution over the Tibetan Plateau by Integrating MODIS Data Products and a DEM. Remote Sens. 2016, 8, 504. [Google Scholar] [CrossRef] [Green Version]
- Wonsick, M.M.; Pinker, R.T. The radiative environment of the Tibetan Plateau. Int. J. Climatol. 2014, 34, 2153–2162. [Google Scholar] [CrossRef]
- Elkeilsh, A.; Awad, Y.M.; Soliman, M.H.; Abu-Elsaoud, A.; Abdelhamid, M.T.; El-Metwally, I.M. Exogenous application of β-sitosterol mediated growth and yield improvement in water-stressed wheat (Triticum aestivum) involves up-regulated antioxidant system. J. Plant Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium protects wheat seedlings against salt stress-mediated oxidative damage by up-regulating antioxidants and osmolytes metabolism. Plant Physiol. Biochem. 2019. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2019, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Soliman, M.H.; Abdulmajeed, A.M.; Alhaithloul, H.; Alharbi, B.M.; El-Esawi, M.A.; Hasanuzzaman, M.; Elkelish, A. Saponin biopriming positively stimulates antioxidants defense, osmolytes metabolism and ionic status to confer salt stress tolerance in soybean. Acta Physiol. Plant 2020, 42, 114. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Elkelish, A.; Soliman, M.; Elansary, H.O.; Zaid, A.; Wani, S.H. Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants 2020, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Soliman, M.H.; Alayafi, A.A.M.; El Kelish, A.A.; Abu-Elsaoud, A.M. Acetylsalicylic acid enhance tolerance of Phaseolus vulgaris L. to chilling stress, improving photosynthesis, antioxidants and expression of cold stress responsive genes. Bot. Stud. 2018, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, M.; Riaz, M.; Soliman, M.H.; Elkelish, A.; et al. Influence of phosphorus on copper phytoextraction via modulating cellular organelles in two jute (Corchorus capsularis L.) varieties grown in a copper mining soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Magdy Beshbishy, A.; Adeyemi, O.S.; Nadwa, E.H.; Rashwan, E.; Kadry, M.; Alkazmi, L.M.; Elkelish, A.A.; Igarashi, I. Phytochemical Screening and Antiprotozoal Effects of the Methanolic Berberis vulgaris and Acetonic Rhus coriaria Extracts. Molecules 2020, 25, 550. [Google Scholar] [CrossRef] [Green Version]
- Rashad, Y.; Aseel, D.; Hammad, S.; Elkelish, A. Rhizophagus irregularis and Rhizoctonia solani Differentially Elicit Systemic Transcriptional Expression of Polyphenol Biosynthetic Pathways Genes in Sunflower. Biomolecules 2020, 10, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julkunen-Tiitto, R.; Rousi, M.; Bryant, J.; Sorsa, S.; Keinänen, M.; Sikanen, H. Chemical diversity of several Betulaceae species: Comparison of phenolics and terpenoids in northern birch stems. Trees 1996, 11, 16–22. [Google Scholar] [CrossRef]
- Gong, J.; Zhang, Z.; Zhang, C.; Zhang, J.; Ran, A. Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau. Forests 2018, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Gea-Izquierdo, G.; Fonti, P.; Cherubini, P.; Martín-Benito, D.; Chaar, H.; Cañellas, I. Xylem hydraulic adjustment and growth response of Quercus canariensis Willd. to climatic variability. Tree Physiol. 2012, 32, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Polle, A.; Baumbusch, L.O.; Oschinski, C.; Eiblmeier, M.; Kuhlenkamp, V.; Vollrath, B.; Scholz, F.; Rennenberg, H. Growth and protection against oxidative stress in young clones and mature spruce trees (Picea abies L.) at high altitudes. Oecologia 1999, 121, 149–156. [Google Scholar] [CrossRef]
- Elkelish, A.; Qari, S.H.; Mazrou, Y.S.A.; Abdelaal, K.A.A.; Hafez, Y.M.; Abu-Elsaoud, A.M.; Batiha, G.E.-S.; El-Esawi, M.A.; El Nahhas, N. Exogenous Ascorbic Acid Induced Chilling Tolerance in Tomato Plants Through Modulating Metabolism, Osmolytes, Antioxidants, and Transcriptional Regulation of Catalase and Heat Shock Proteins. Plants 2020, 9, 431. [Google Scholar] [CrossRef] [Green Version]
- Soliman, M.; Elkelish, A.; Souad, T.; Alhaithloul, H.; Farooq, M. Brassinosteroid seed priming with nitrogen supplementation improves salt tolerance in soybean. Physiol. Mol. Biol. Plants 2020. [Google Scholar] [CrossRef]
- Conklin, P.L. Recent advances in the role and biosynthesis of ascorbic acid in plants. Plant Cell Environ. 2001, 24, 383–394. [Google Scholar] [CrossRef]
- Soliman, M.; Alhaithloul, H.A.; Hakeem, K.R.; Alharbi, B.M.; El-Esawi, M.; Elkelish, A. Exogenous Nitric Oxide Mitigates Nickel-Induced Oxidative Damage in Eggplant by Upregulating Antioxidants, Osmolyte Metabolism, and Glyoxalase Systems. Plants 2019, 8, 562. [Google Scholar] [CrossRef] [Green Version]
- Zamin, M.; Fahad, S.; Khattak, A.M.; Adnan, M.; Wahid, F.; Raza, A.; Wang, D.; Saud, S.; Noor, M.; Bakhat, H.F.; et al. Developing the first halophytic turfgrasses for the urban landscape from native Arabian desert grass. Env. Sci. Pollut. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Streb, P.; Aubert, S.; Gout, E.; Bligny, R. Reversibility of cold- and light-stress tolerance and accompanying changes of metabolite and antioxidant levels in the two high mountain plant species Soldanella alpina and Ranunculus glacialis. J. Exp. Bot. 2003, 54, 405–418. [Google Scholar] [CrossRef]
- Öncel, I.; Yurdakulol, E.; Keleş, Y.; Kurt, L.; Yıldız, A. Role of antioxidant defense system and biochemical adaptation on stress tolerance of high mountain and steppe plants. Acta Oecologica 2004, 26, 211–218. [Google Scholar] [CrossRef]
- Hegazy, A.; Doust, J.L. Plant Ecology in the Middle East; Oxford University Press: Oxford, UK, 2016; ISBN 978-0-19-966081-0. [Google Scholar]
- Helal, N.M.; Alharby, H.F.; Alharbi, B.M.; Bamagoos, A.A.; Hashim, A.M. Thymelaea hirsuta and Echinops spinosus: Xerophytic Plants with High Potential for First-Generation Biodiesel Production. Sustainability 2020, 12, 1137. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Wahab, R.; Zayed, A.E.-M.; Moustafa, A.E.-R.; Klopatek, J.; Helmy, M. Landforms, Vegetation, and Soil Quality in South Sinai, Egypt. Catrina Int. J. Environ. Sci. 2018, 1, 127–138. [Google Scholar]
- Moustafa, A.E.-R.A.; Zaghloul, M.S. Environment and vegetation in the montane Saint Catherine area, south Sinai, Egypt. J. Arid Environ. 1996, 34, 331–349. [Google Scholar] [CrossRef]
- Neina, D. The Role of Soil pH in Plant Nutrition and Soil Remediation. Available online: https://www.hindawi.com/journals/aess/2019/5794869/ (accessed on 12 April 2020).
- Santiago, L.S. Use of Coarse Woody Debris by the Plant Community of a Hawaiian Montane Cloud Forest1. Biotropica 2000, 32, 633–641. [Google Scholar] [CrossRef]
- Soethe, N.; Wilcke, W.; Homeier, J.; Lehmann, J.; Engels, C. Plant Growth along the Altitudinal —Role of Plant Nutritional Status, Fine Root Activity, and Soil Properties. In Gradients in a Tropical Mountain Ecosystem of Ecuador; Ecological Studies; Beck, E., Bendix, J., Kottke, I., Makeschin, F., Mosandl, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 259–266. ISBN 978-3-540-73526-7. [Google Scholar]
- Sharma, P.; Rana, J.C.; Devi, U.; Randhawa, S.S.; Kumar, R. Floristic Diversity and Distribution Pattern of Plant Communities along Altitudinal Gradient in Sangla Valley, Northwest Himalaya. Available online: https://www.hindawi.com/journals/tswj/2014/264878/ (accessed on 12 April 2020).
- Chen, Y.; Zhang, X.; Guo, Q.; Cao, L.; Qin, Q.; Li, C.; Zhao, M.; Wang, W. Plant morphology, physiological characteristics, accumulation of secondary metabolites and antioxidant activities of Prunella vulgaris L. under UV solar exclusion. Biol. Res. 2019, 52. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambers, H.; Oliveira, R.S. Plant Physiological Ecology; Springer Nature: Berlin/Heidelberg, Germany, 2019; ISBN 978-3-030-29639-1. [Google Scholar]
- Guidi, L.; Tattini, M.; Landi, M. How Does Chloroplast Protect Chlorophyll Against Excessive Light? In Chlorophyll; Jacob-Lopes, E., Zepka, L.Q., Queiroz, M.I., Eds.; InTech: Vienna, Austria, 2017; ISBN 978-953-51-3107-6. [Google Scholar]
- Castrillo, M. Photosynthesis in three altitudinal populations of the Andean plant Espeletia schultzii (Compositae). Rev. Biol. Trop. 2006, 54, 1143–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Elkelish, A.; Alnusaire, T.S.; Soliman, M.H.; Gowayed, S.; Senousy, H.H.; Fahad, S. Calcium availability regulates antioxidant system, physio-biochemical activities and alleviates salinity stress mediated oxidative damage in soybean seedlings. J. Appl. Bot. Food Qual. 2019, 258–266. [Google Scholar] [CrossRef]
- D’angiolillo, F.; Mammano, M.M.; Fascella, G. Pigments, Polyphenols and Antioxidant Activity of Leaf Extracts from Four Wild Rose Species Grown in Sicily. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 402–409. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Gallardo, M.G.; Boero, C.; Liberman Cruz, M.; Prado, F.E. Altitudinal and seasonal variation of protective and photosynthetic pigments in leaves of the world’s highest elevation trees Polylepis tarapacana (Rosaceae). Acta Oecologica 2007, 32, 36–41. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoid oxidation products as stress signals in plants. Plant J. 2014, 79, 597–606. [Google Scholar] [CrossRef]
- Kusvuran, S. Influence of drought stress on growth, ion accumulation and antioxidative enzymes in okra genotypes. Int. J. Agric. Biol. 2012, 14, 401–406. [Google Scholar]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S. Antioxidant Enzyme Activities and Abiotic Stress Tolerance Relationship in Vegetable Crops. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; InTech: Vienna, Austria, 2016; ISBN 978-953-51-2250-0. [Google Scholar]
- El-Ghani, M.M.A.; Huerta-Martínez, F.M.; Hongyan, L.; Qureshi, R. Plant Responses to Hyperarid Desert Environments; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 978-3-319-59135-3. [Google Scholar]
- Logan, B.A. Reactive Oxygen Species and Photosynthesis. In Antioxidants and Reactive Oxygen Species in Plants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; pp. 250–267. ISBN 978-0-470-98856-5. [Google Scholar]
- Sharma, P.; Dubey, R.S. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 2007, 26, 2027–2038. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]
- Li, S.; Yang, Y.; Zhang, Q.; Liu, N.; Xu, Q.; Hu, L. Differential physiological and metabolic response to low temperature in two zoysiagrass genotypes native to high and low latitude. PLoS ONE 2018, 13, e0198885. [Google Scholar] [CrossRef] [PubMed]
- Gidalevitz, T.; Prahlad, V.; Morimoto, R.I. The Stress of Protein Misfolding: From Single Cells to Multicellular Organisms. Cold Spring Harb Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andresen, I.; Becker, W.; Schlüter, K.; Burges, J.; Parthier, B.; Apel, K. The identification of leaf thionin as one of the main jasmonate-induced proteins of barley (Hordeum vulgare). Plant Mol. Biol. 1992, 19, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Hemantaranjan, A. Advances in Plant Physiology; Scientific Publishers: Oxford, UK, 2013; Volume 14, ISBN 978-93-86237-59-0. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- ElKelish, A.E.; Winkler, J.B.; Lang, H.; Holzinger, A.; Behrendt, H.; Durner, J.; Kanter, U.; Ernst, D. Effects of ozone, CO2 and drought stress on the growth and pollen production of common ragweed (Ambrosia artemisiifolia); Julius Kühn Institut, Bundesforschungsinstitut für Kulturpflanzen: Quedlinburg, Germany, 2014; pp. 139–147. [Google Scholar]
- Cirak, C.; Radusiene, J.; Jakstas, V.; Ivanauskas, L.; Seyis, F.; Yayla, F. Altitudinal changes in secondary metabolite contents of Hypericum androsaemum and Hypericum polyphyllum. Biochem. Syst. Ecol. 2017, 70, 108–115. [Google Scholar] [CrossRef]
- Watson, R.R. Polyphenols in Plants: Isolation, Purification and Extract Preparation; Academic Press: New York, NY, USA, 2014; ISBN 978-0-12-398491-3. [Google Scholar]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of Secondary Metabolites and Brassinosteroids in Plant Defense Against Environmental Stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Gourlay, G.; Constabel, C.P. Condensed tannins are inducible antioxidants and protect hybrid poplar against oxidative stress. Tree Physiol. 2019, 39, 345–355. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; da Silva, J.A.T.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense Is a Key Factor. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A.K., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. ISBN 978-94-007-2220-0. [Google Scholar]
- Dai, L.-P.; Xiong, Z.-T.; Huang, Y.; Li, M.-J. Cadmium-induced changes in pigments, total phenolics, and phenylalanine ammonia-lyase activity in fronds of Azolla imbricata. Env. Toxicol. 2006, 21, 505–512. [Google Scholar] [CrossRef]
- Morales, F.; Ancín, M.; Fakhet, D.; González-Torralba, J.; Gámez, A.L.; Seminario, A.; Soba, D.; Ben Mariem, S.; Garriga, M.; Aranjuelo, I. Photosynthetic Metabolism under Stressful Growth Conditions as a Bases for Crop Breeding and Yield Improvement. Plants 2020, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sai, K.; Thapa, R.; Devkota, H.P.; Joshi, K.R. Phytochemical Screening, Free Radical Scavenging and α-Amylase Inhibitory Activities of Selected Medicinal Plants from Western Nepal. Medicines 2019, 6, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. Available online: https://www.hindawi.com/journals/jb/2012/217037/ (accessed on 12 April 2020).
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A. Phytohormones: Signaling Mechanisms and Crosstalk in Plant Development and Stress Responses; BoD—Books on Demand: Norderstedt, Germany, 2017; ISBN 978-953-51-3411-4. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Liu, W.; Yin, D.; Li, N.; Hou, X.; Wang, D.; Li, D.; Liu, J. Influence of Environmental Factors on the Active Substance Production and Antioxidant Activity in Potentilla fruticosa L. and Its Quality Assessment. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Moustafa, A.G. Long Term Monitoring of Rosa arabica Populations as a Threatened Species in South Sinai, Egypt. Available online: https://www.semanticscholar.org/paper/Long-Term-Monitoring-of-Rosa-arabica-Populations-as-Moustafa/a9209f600f5eef6928a0df021c7866cea10018f1 (accessed on 2 April 2020).
- El-Wahab, R.A.; Zaghloul, M.S.; Moustafa, A.A. Conservation of Medicinal plants in St. Catherine Protectorate, South Sinai, Egypt. In Proceedings of the First International Conference on Strategy of Egyptian Herbaria, Dokki, Egypt, 9–11 March 2004; pp. 231–251. [Google Scholar]
- Danin, A. Desert Rocks—A Habitat Which Supports Many Species That Were New to Science in the Last 40 Years. Turk. J. Bot. 2008, 32, 459–464. [Google Scholar]
- Fakhry, A.M.; El-Keblawy, A.; Shabana, H.A.; Gamal, I.E.; Shalouf, A. Microhabitats Affect Population Size and Plant Vigor of Three Critically Endangered Endemic Plants in Southern Sinai Mountains, Egypt. Land 2019, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, A.E.-R.A.; Zayed, A. Effect of environmental factors on the flora of alluvial fans in southern Sinai. J. Arid Environ. 1996, 32, 431–443. [Google Scholar] [CrossRef]
- Omar, K.A. Ecological and Climatic Attribute Analysis for Egyptian Hypericum sinaicum. Am. J. Life Sci. 2014, 2, 369. [Google Scholar] [CrossRef]
- Dewis, J.; Freitas, F. Physical and Chemical Methods of Soil and Water Analysis; Food and Agriculture Organization of the United Nations: Rome, Italy, 1984; ISBN 978-92-5-102010-4. [Google Scholar]
- Blume, H.-P. Page, A.L., R. H. Miller and D.R. Keeney (Ed., 1982): Methods of soil analysis; 2. Chemical and microbiological properties, 2. Aufl. 1184 S., American Soc. of Agronomy (Publ.), Madison, Wisconsin, USA, gebunden 36 Dollar. Zeitschrift für Pflanzenernährung und Bodenkunde 1985, 148, 363–364. [Google Scholar] [CrossRef]
- Johnson, C.M.; Ulrich, A. Analytical Methods for Use in Plant Analysis; University of California: Oakland, CA, USA, 1959. [Google Scholar]
- Shapiro, L.; Brannock, W.W. A Field Method for the Determination of Calcium and Magnesium in Limestone and Dolomite; Open-File Report; U.S. Geological Survey: Reston, VA, USA, 1957; Volume 57–99, p. 9.
- Black, C.A.; American Society of Agronomy; American Society for Testing and Materials. Methods of soil Analysis Part 1; American Society of Agronomy: Crop Science Society of America: Soil Science Society of America: Madison, WI, USA, 1965. [Google Scholar]
- Metzner, H.; Rau, H.; Senger, H. Untersuchungen zur Synchronisierbarkeit einzelner Pigmentmangel-Mutanten von Chlorella. Planta 1965, 65, 186–194. [Google Scholar] [CrossRef]
- Prud’homme, M.-P.; Gonzalez, B.; Billard, J.-P.; Boucaud, J. Carbohydrate Content, Fructan and Sucrose Enzyme Activities in Roots, Stubble and Leaves of Ryegrass (Lolium perenne L.) as Affected by Source/Sink Modification after Cutting. J. Plant Physiol. 1992, 140, 282–291. [Google Scholar] [CrossRef]
- Loewus, F.A. Improvement in Anthrone Method for Determination of Carbohydrates. Anal. Chem. 1952, 24, 219. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histo-Enzymology: A Text Manual; Kalyani Publishers: Delhi, India, 1980. [Google Scholar]
- Harborne, A.J. Phytochemical Methods A Guide to Modern Techniques of Plant Analysis, 3rd ed.; Springer: Amsterdam, The Netherlands, 1998; ISBN 978-0-412-57260-9. [Google Scholar]
- Ejikeme, C.M.; Ezeonu, C.S.; Eboatu, A.N. Determination of Physical and Phytochemical Constituents of Some Tropical Timbers Indigenous to Niger Delta Area of Nigeria. Eur. Sci. J. ESJ 2014, 10, 247–270. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Hermans, C.; Conn, S.J.; Chen, J.; Xiao, Q.; Verbruggen, N. An update on magnesium homeostasis mechanisms in plants. Metallomics 2013, 5, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Kar, M.; Mishra, D. Catalase, Peroxidase, and Polyphenoloxidase Activities during Rice Leaf Senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Koricheva, J.; Roy, S.; Vranjic, J.A.; Haukioja, E.; Hughes, P.R.; Hänninen, O. Antioxidant responses to simulated acid rain and heavy metal deposition in birch seedlings. Env. Pollut. 1997, 95, 249–258. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Montagu, M.V.; Inze, D. Effects of Iron Excess on Nicotiana plumbaginifolia Plants (Implications to Oxidative Stress). Plant Physiol. 1995, 107, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Altitude (m a.s.l.) | pH Value | Anions (ppm) | Cations (ppm) | |||||
|---|---|---|---|---|---|---|---|---|
| HCO3− | SO4− | Cl− | Ca++ | Mg++ | Na+ | K+ | ||
| 1600–1800 | 8.2 ± 0.12 a | 165.6 ± 7.5 b | 95.6 ± 2 a | 34 ± 4.9 a | 79.5 ± 1.5 a | 6.6 ± 0.4 c | 28.0 ± 1.26 a | 7 ± 1.15 b |
| 1800–2000 | 7.8 ± 0.03 ab | 228.7 ± 11.4a | 59.1 ± 1.7 b | 31.2 ± 5.1 a | 42.2 ± 3.5 b | 10.8 ± 0.37 a | 8.6 ± 1.33 b | 14.3 ± 0.88 b |
| 2000–2200 | 7.6 ± 0.22 b | 190.6 ± 5.6 b | 18.6 ± 4.5 c | 27.5 ± 1.3 a | 74.0 ± 0.85 a | 8.9 ± 0.29 b | 8.0 ± 1.15 b | 33.3 ± 3.84 a |
| Family | Plant Species | Altitude (m a.s.l.) | Field Photo | ||
|---|---|---|---|---|---|
| (1600–1800) | (1800–2000) | (2000–2200) | |||
| Lamiaceae | Nepeta septemcrenata Benth. | 1630 | 1945 | 2038 |  |
| Origanum syriacum subsp. Sinaicum (Boiss.) Greater and Burdet. | 1630 | 1825 | 2038 |  | |
| Phlomis aurea Decene. | 1710 | 1825 | 2038 |  | |
| Rosaceae | Rosa arabica Crep. | 1750 | 1940 | 2150 |  |
| Caryophyllaceae | Silene schimperiana Boiss. | 1750 | 1825 | 2110 |  |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. https://doi.org/10.3390/plants9070869
Hashim AM, Alharbi BM, Abdulmajeed AM, Elkelish A, Hozzein WN, Hassan HM. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants. 2020; 9(7):869. https://doi.org/10.3390/plants9070869
Chicago/Turabian StyleHashim, Ahmed M., Basmah M. Alharbi, Awatif M. Abdulmajeed, Amr Elkelish, Wael N. Hozzein, and Heba M. Hassan. 2020. "Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content" Plants 9, no. 7: 869. https://doi.org/10.3390/plants9070869