Biomolecule from Trigonella stellata from Saudi Flora to Suppress Osteoporosis via Osteostromal Regulations


 ,
,
Abstract
:1. Introduction
2. Results
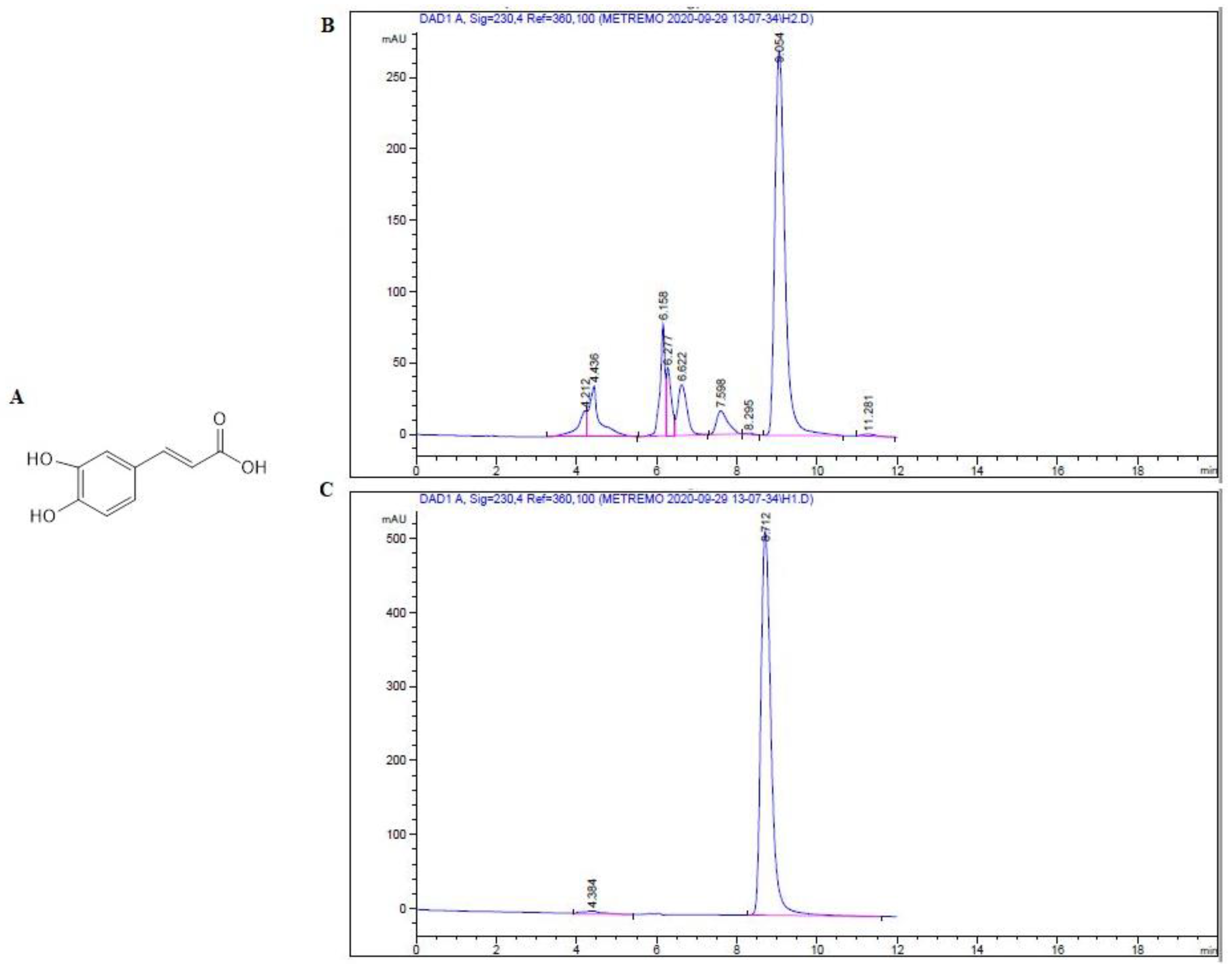
2.1. Isolation and Identification of Major Compound
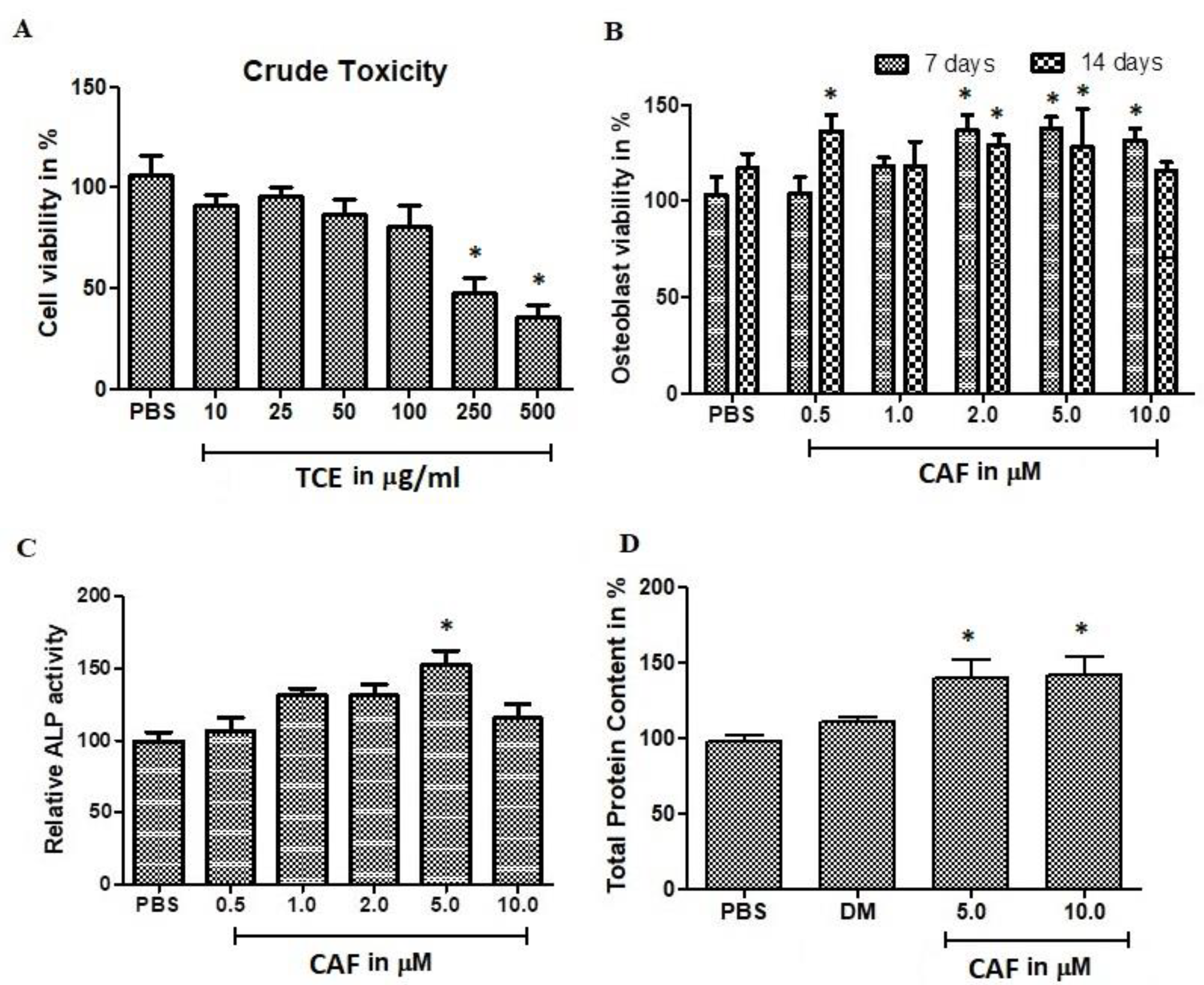
2.2. Effect of T. Stellata Extract Against Mesenchymal Cells
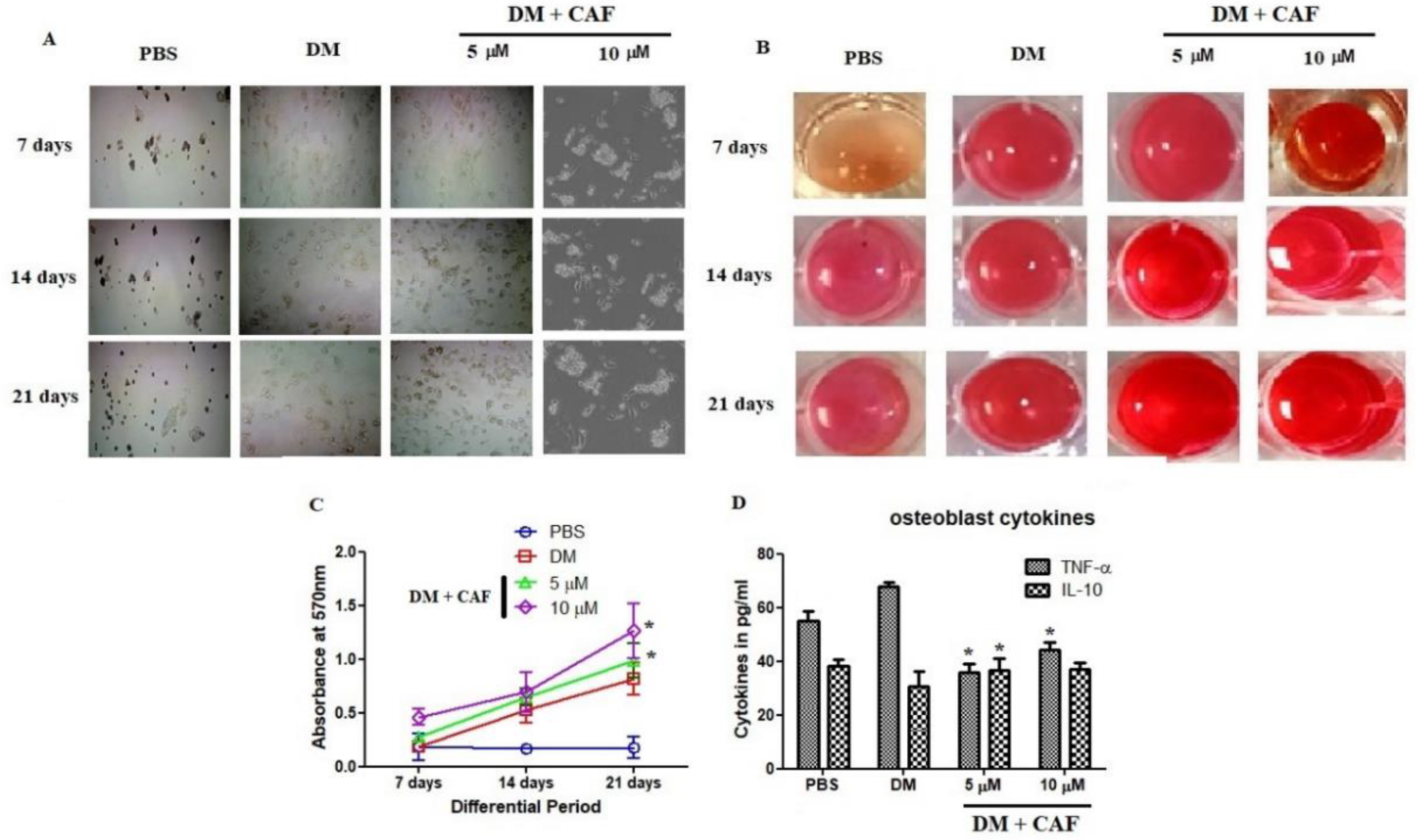
2.3. CAF Regulated In Vitro Mineralization of Osteoblastic Cells
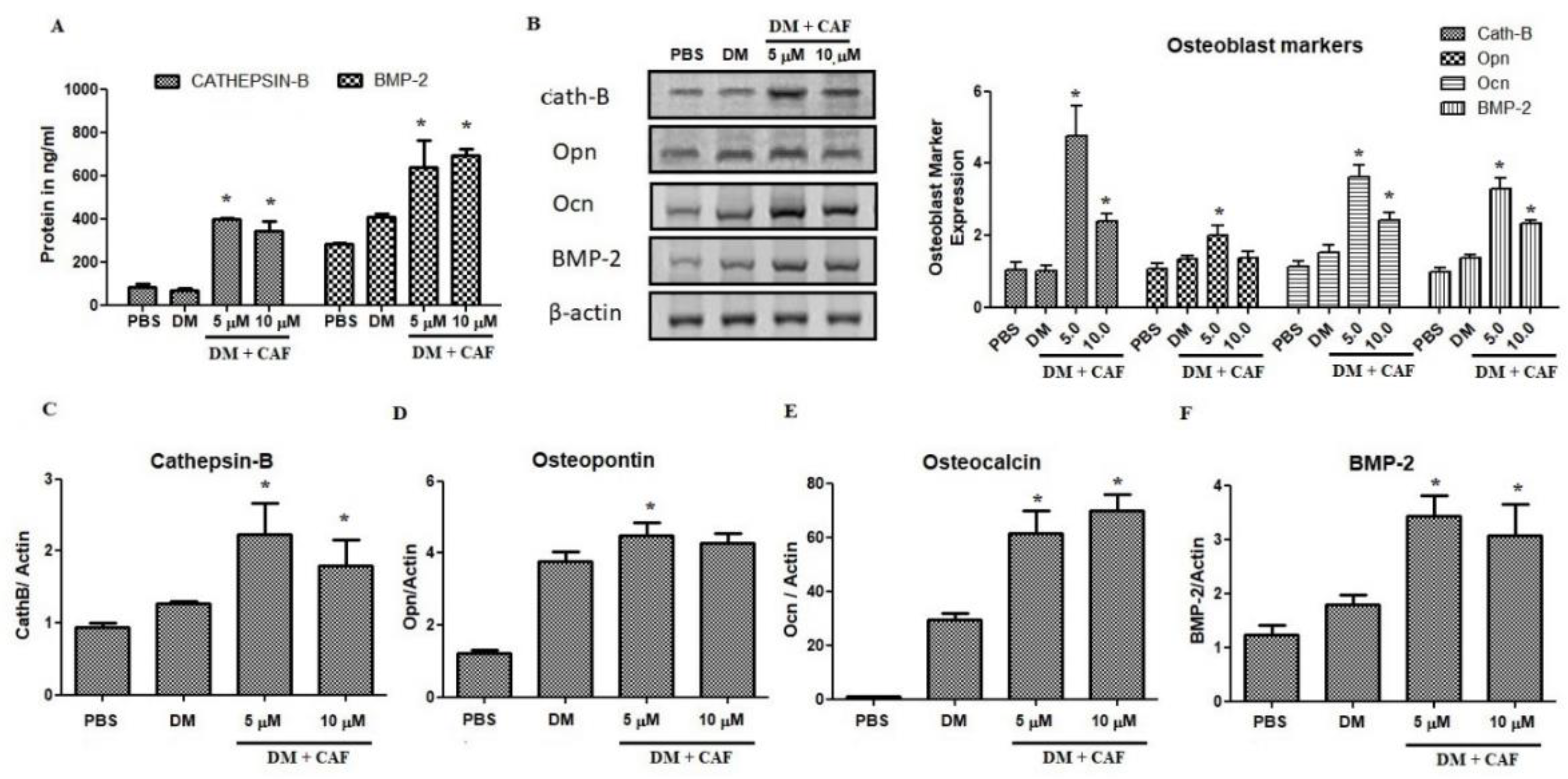
2.4. Regulation of CAF on Osteoblastic Markers
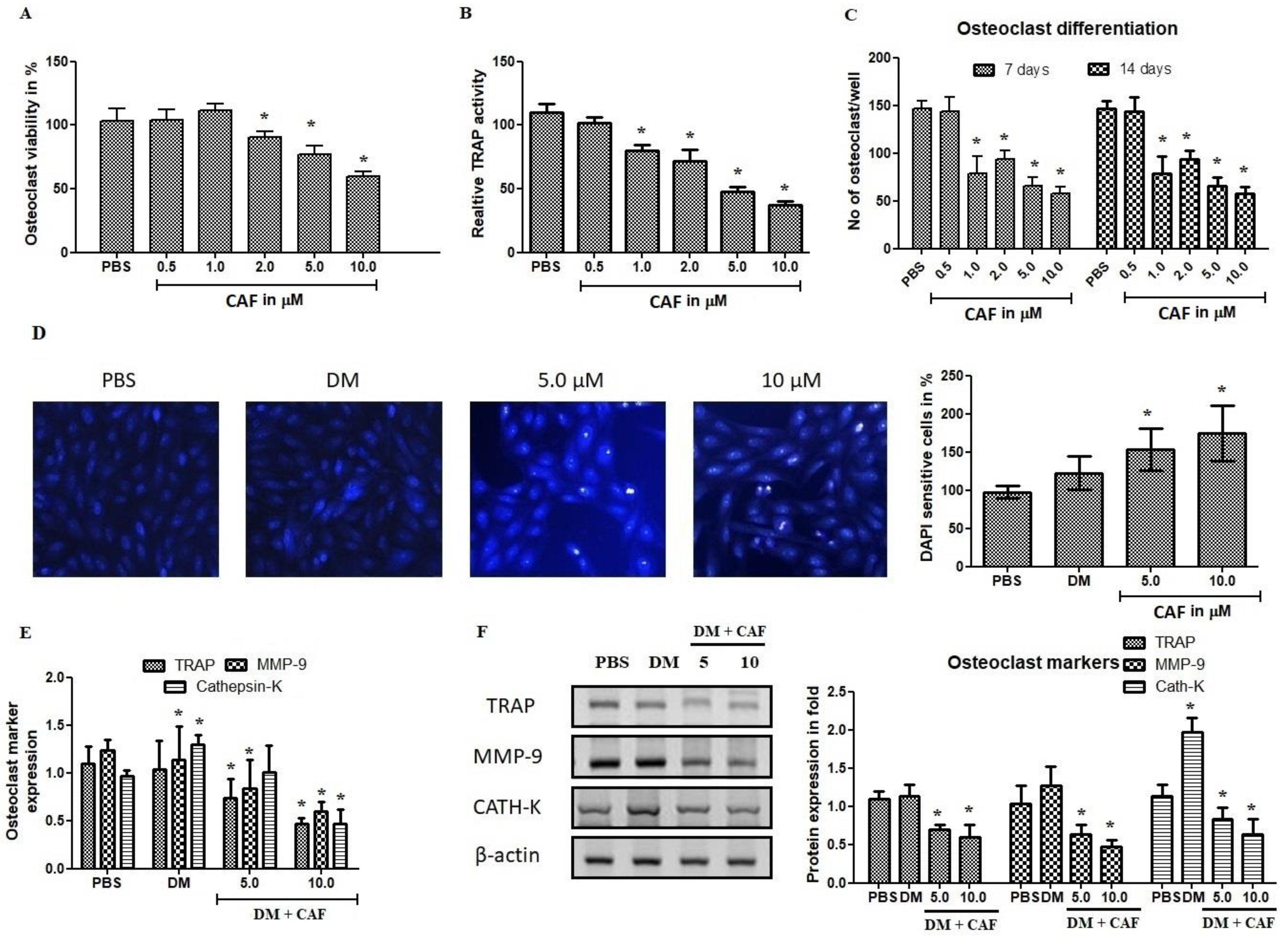
2.5. Effect of CAF on Osteoclastogenic Regulations
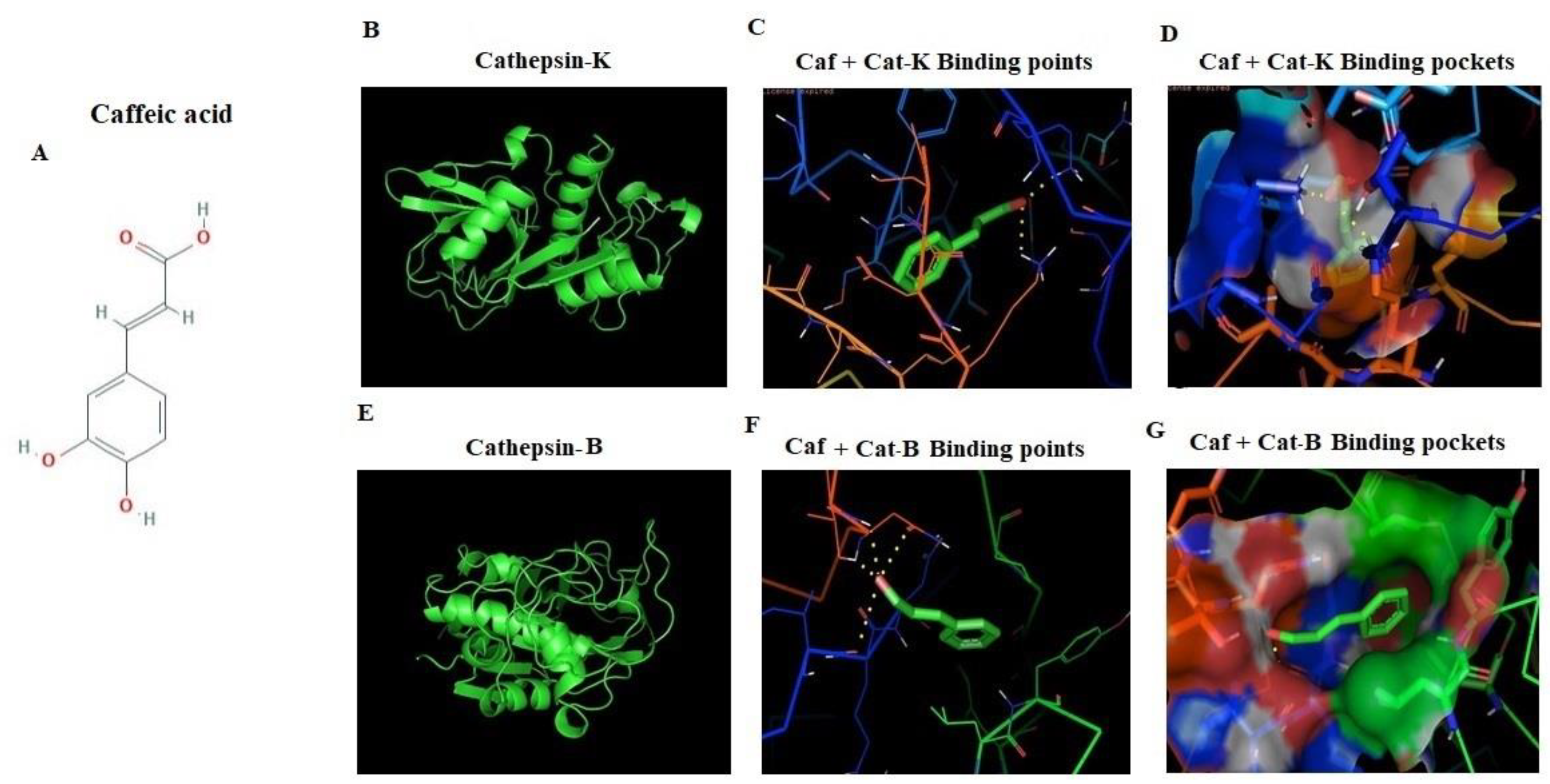
2.6. In Silico Docking of CAF against Cathepsin-B and Cathepsin-K Markers
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures and Chemicals
4.2. Plant Material
4.3. Extraction and Isolation
4.4. Animals and Ethical Aspects
4.5. Culture of BM-MSCs
4.6. In Vitro Cytotoxicity Analyses
4.6.1. MTT Assay
4.6.2. Neutral Red Assay
4.7. Alkaline Phosphatase Activity
4.8. Assays of Osteoblast Differentiation
4.9. Osteoclast Tartrate-Resistant Acid Phosphatase (TRAP) Activity Estimation
Osteoclast Apoptosis
4.10. Gene Expression Analysis by Real-Time Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR)
4.11. Western Blot for Protein Marker Quantification
4.12. Cytokine Estimation by ELISA
4.13. Computational Docking Analysis
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Florencio-Silva, R.; Sasso, G.R.d.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maniatopoulos, C.; Sodek, J.; Melcher, A. Bone formation in vitro by stromal cells obtained from bone marrow of young adult rats. Cell Tissue Res. 1988, 254, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Komori, M.; Rasmussen, S.W.; Kiel, J.A.; Baerends, R.J.; Cregg, J.M.; van der Klei, I.J.; Veenhuis, M. The Hansenula polymorpha PEX14 gene encodes a novel peroxisomal membrane protein essential for peroxisome biogenesis. EMBO J. 1997, 16, 44–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suda, T.; Takahashi, N.; Udagawa, N.; Jimi, E.; Gillespie, M.T.; Martin, T.J. Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr. Rev. 1999, 20, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Jiang, D.; Ge, C.; Zhao, Z.; Lai, Y.; Boules, H.; Phimphilai, M.; Yang, X.; Karsenty, G.; Franceschi, R.T. Cooperative interactions between activating transcription factor 4 and Runx2/Cbfa1 stimulate osteoblast-specific osteocalcin gene expression. J. Biol. Chem. 2005, 280, 30689–30696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razzaq, A.; Ahmad Malik, S.; Saeed, F.; Imran, A.; Rasul, A.; Qasim, M.; Zafar, S.; Kamran, S.K.S.; Maqbool, J.; Imran, M. Moringa oleifera Lam. ameliorates the muscles function recovery following an induced insult to the Sciatic nerve in a mouse model. Food Sci. Nutr. 2020, 8, 4009–4016. [Google Scholar] [CrossRef] [PubMed]
- Vlachogianni, T.; Loridas, S.; Fiotakis, K.; Valavanidis, A. From the traditional medicine to the modern era of synthetic pharmaceuticals natural products and reverse pharmacology approaches in new drug discovery expedition. Pharmakeftiki 2014, 26, 16–30. [Google Scholar]
- Nielsen, O.; Ainsworth, M.; Csillag, C.; Rask-Madsen, J. Systematic review: Coxibs, non-steroidal anti-inflammatory drugs or no cyclooxygenase inhibitors in gastroenterological high-risk patients? Aliment. Pharmacol. Ther. 2006, 23, 27–33. [Google Scholar] [CrossRef]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-inflammatory activity of natural products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef]
- Owona, B.A.; Abia, W.A.; Moundipa, P.F. Natural compounds flavonoids as modulators of inflammasomes in chronic diseases. Int. Immunopharmacol. 2020, 84, 106498. [Google Scholar] [CrossRef]
- Nunes, C.d.R.; Barreto Arantes, M.; Menezes de Faria Pereira, S.; Leandro da Cruz, L.; de Souza Passos, M.; Pereira de Moraes, L.; Vieira, I.J.C.; Barros de Oliveira, D. Plants as Sources of Anti-Inflammatory Agents. Molecules 2020, 25, 3726. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.A.; Díaz, M.A.; Gupta, M.P.; López-Pérez, J.L.; Mondolis, E.; Morán-Pinzón, J.; Guerrero, E. Assessment of the antinociceptive and anti-inflammatory activities of the stem methanol extract of Diplotropis purpurea. Pharm. Biol. 2019, 57, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Nagulapalli Venkata, K.C.; Swaroop, A.; Bagchi, D.; Bishayee, A. A small plant with big benefits: Fenugreek (Trigonella foenum-graecum Linn.) for disease prevention and health promotion. Mol. Nutr. Food Res. 2017, 61, 1600950. [Google Scholar] [CrossRef]
- Sindhu, G.; Ratheesh, M.; Shyni, G.; Nambisan, B.; Helen, A. Anti-inflammatory and antioxidative effects of mucilage of Trigonella foenum graecum (Fenugreek) on adjuvant induced arthritic rats. Int. Immunopharmacol. 2012, 12, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Norton, J.; Majid, S.A.; Allan, D.; Al Safran, M.; Böer, B.; Richer, R. An Illustrated Checklist of The Flora of Qatar; Browndown Publications: Gosport, UK, 2009. [Google Scholar]
- Al-Tabini, R.; Al-Khalidi, K.; Al-Shudiefat, M. Livestock, medicinal plants and rangeland viability in Jordan’s Badia: Through the lens of traditional and local knowledge. Pastor. Res. Policy Pract. 2012, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Alhabri, N. Survey of plant species of medical importance to treat digestive tract diseases in Tabuk Region, Saudi Arabia. J. King Abdulaziz Univ. 2017, 29, 51–56. [Google Scholar]
- Basu, S.K.; Zandi, P.; Cetzal-Ix, W. Fenugreek (Trigonella foenum-graecum L.): Distribution, Genetic Diversity, and Potential to Serve as an Industrial Crop for the Global Pharmaceutical, Nutraceutical, and Functional Food Industries. In The Role of Functional Food Security in Global Health; Elsevier: Amsterdam, The Netherlands, 2019; pp. 471–497. [Google Scholar]
- Shams Eldin, S.M.; Radwan, M.M.; Wanas, A.S.; Habib, A.-A.M.; Kassem, F.F.; Hammoda, H.M.; Khan, S.I.; Klein, M.L.; Elokely, K.M.; ElSohly, M.A. Bioactivity-guided isolation of potential antidiabetic and antihyperlipidemic compounds from Trigonella stellata. J. Nat. Prod. 2018, 81, 1154–1161. [Google Scholar] [CrossRef] [PubMed]
- Sheweita, S.A.; ElHady, S.A.; Hammoda, H.M. Trigonella stellata reduced the deleterious effects of diabetes mellitus through alleviation of oxidative stress, antioxidant-and drug-metabolizing enzymes activities. J. Ethnopharmacol. 2020, 112821. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, J.; Liao, X.; Zhang, P.; Chen, X. One-step separation of antioxidant compounds from Erythrina variegata by high speed counter-current chromatography. J. Chromatogr. Sci. 2015, 53, 730–735. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, T.; Kawaguchi, H.; Jinno, S.; Hoshi, K.; Itaka, K.; Takato, T.; Nakamura, K.; Okayama, H. Bone morphogenetic protein 2-induced osteoblast differentiation requires Smad-mediated down-regulation of Cdk6. Mol Cell Biol 2004, 24, 6560–6568. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-W.; Kim, S.-H.; Lee, K.-S.; Kang, H.J.; Lee, M.J.; Park, K.-I.; Lee, J.H.; Park, K.D.; Seo, W.D. Barley Seedling Extracts Inhibit RANKL-Induced Differentiation, Fusion, and Maturation of Osteoclasts in the Early-to-Late Stages of Osteoclastogenesis. Evid. Based Complementary Altern. Med. 2017, 2017, 6072573. [Google Scholar] [CrossRef]
- Choi, S.-W.; Lee, K.-S.; Lee, J.H.; Kang, H.J.; Lee, M.J.; Kim, H.Y.; Park, K.-I.; Kim, S.-L.; Shin, H.K.; Seo, W.D. Suppression of Akt-HIF-1α signaling axis by diacetyl atractylodiol inhibits hypoxia-induced angiogenesis. BMB Rep. 2016, 49, 508–513. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.-W.; Choi, S.-W.; Kim, H.-J.; Lee, K.-S.; Kim, S.-H.; Kim, S.-L.; Do, S.H.; Seo, W.-D. Germinated soy germ with increased soyasaponin Ab improves BMP-2-induced bone formation and protects against in vivo bone loss in osteoporosis. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Kim, N.; Takami, M.; Rho, J.; Josien, R.; Choi, Y. A novel member of the leukocyte receptor complex regulates osteoclast differentiation. J. Exp. Med. 2002, 195, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2. [Google Scholar] [CrossRef] [Green Version]
- Prynne, C.J.; Mishra, G.D.; O’Connell, M.A.; Muniz, G.; Laskey, M.A.; Yan, L.; Prentice, A.; Ginty, F. Fruit and vegetable intakes and bone mineral status: A cross-sectional study in 5 age and sex cohorts. Am. J. Clin. Nutr. 2006, 83, 1420–1428. [Google Scholar] [CrossRef]
- Chin, K.-Y.; Ima-Nirwana, S. Olives and bone: A green osteoporosis prevention option. Int. J. Environ. Res. Public Health 2016, 13, 755. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef]
- Jao, H.-Y.; Hsu, J.-D.; Lee, Y.-R.; Lo, C.-S.; Lee, H.-J. Mulberry water extract regulates the osteoblast/osteoclast balance in an ovariectomic rat model. Food Funct. 2016, 7, 4753–4763. [Google Scholar] [CrossRef]
- Shim, K.-S.; Lee, C.-J.; Yim, N.-H.; Gu, M.J.; Ma, J.Y. Alpinia officinarum stimulates osteoblast mineralization and inhibits osteoclast differentiation. Am. J. Chin. Med. 2016, 44, 1255–1271. [Google Scholar] [CrossRef]
- Mukudai, Y.; Kondo, S.; Koyama, T.; Li, C.; Banka, S.; Kogure, A.; Yazawa, K.; Shintani, S. Potential anti-osteoporotic effects of herbal extracts on osteoclasts, osteoblasts and chondrocytes in vitro. BMC Complementary Altern. Med. 2014, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devalaraja, S.; Jain, S.; Yadav, H. Exotic fruits as therapeutic complements for diabetes, obesity and metabolic syndrome. Food Res. Int. 2011, 44, 1856–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanham-New, S.A. Fruit and Vegetables: The Unexpected Natural Answer to the Question of Osteoporosis Prevention? Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Sharan, K.; Siddiqui, J.A.; Swarnkar, G.; Maurya, R.; Chattopadhyay, N. Role of phytochemicals in the prevention of menopausal bone loss: Evidence from in vitro and in vivo, human interventional and pharmacokinetic studies. Curr. Med. Chem. 2009, 16, 1138–1157. [Google Scholar] [CrossRef] [PubMed]
- Shim, K.S.; Lee, B.; Ma, J.Y. Water extract of Rumex crispus prevents bone loss by inhibiting osteoclastogenesis and inducing osteoblast mineralization. BMC Complementary Altern. Med. 2017, 17, 1–9. [Google Scholar] [CrossRef]
- Ahn, S.H.; Chen, Z.; Lee, J.; Lee, S.-W.; Min, S.H.; Kim, N.D.; Lee, T.H. Inhibitory Effects of 2N1HIA (2-(3-(2-Fluoro-4-Methoxyphenyl)-6-Oxo-1(6H)-Pyridazinyl)-N-1H-Indol-5-Ylacetamide) on Osteoclast Differentiation via Suppressing Cathepsin K Expression. Molecules 2018, 23, 3139. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-J.; Jeong, M.-H.; Lee, Y.; Hwang, S.-J.; Shin, H.-B.; Hur, J.-S.; Son, Y.-J. Effect of Usnic Acid on Osteoclastogenic Activity. J. Clin. Med. 2018, 7, 345. [Google Scholar] [CrossRef] [Green Version]
- Bikle, D.D. Vitamin D and bone. Curr. Osteoporos. Rep. 2012, 10, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Deng, C.; Li, Y.-P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Maridas, D.E.; Rendina-Ruedy, E.; Le, P.T.; Rosen, C.J. Isolation, culture, and differentiation of bone marrow stromal cells and osteoclast progenitors from mice. JoVE 2018, 131, e56750. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Jakob, C.A.; Aebi, M.; Tuor, U. The MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide] assay is a fast and reliable method for colorimetric determination of fungal cell densities. Appl. Environ. Microbiol. 1999, 65, 3727–3729. [Google Scholar] [CrossRef] [Green Version]
- Borenfreund, E.; Puerner, J.A. A simple quantitative procedure using monolayer cultures for cytotoxicity assays (HTD/NR-90). J. Tissue Cult. Methods 1985, 9, 7–9. [Google Scholar] [CrossRef]
- Roy, A. Rapid method for determining alkaline phosphatase activity in serum with thymolphthalein monophosphate. Clin. Chem. 1970, 16, 431–436. [Google Scholar] [CrossRef]
- Kameda, T.; Mano, H.; Yamada, Y.; Takai, H.; Amizuka, N.; Kobori, M.; Izumi, N.; Kawashima, H.; Ozawa, H.; Ikeda, K. Calcium-sensing receptor in mature osteoclasts, which are bone resorbing cells. Biochem. Biophys. Res. Commun. 1998, 245, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, D.; Choi, J.; Choi, H.; Ryu, J.-H.; Jeong, J.; Park, E.-J.; Kim, S.-H.; Kim, S. Dehydrodiconiferyl alcohol isolated from Cucurbita moschata shows anti-adipogenic and anti-lipogenic effects in 3T3-L1 cells and primary mouse embryonic fibroblasts. J. Biol. Chem. 2012, 287, 8839–8851. [Google Scholar] [CrossRef] [Green Version]
- Laurie, A.T.; Jackson, R.M. Q-SiteFinder: An energy-based method for the prediction of protein–ligand binding sites. Bioinformatics 2005, 21, 1908–1916. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC | δH, mult. (J in Hz) |
|---|---|---|
| 1 | 127.82 | - |
| 2 | 115.10 | 7.06, d, (2.04) |
| 3 | 146.82 | - |
| 4 | 149.47 | - |
| 5 | 116.51 | 6.80, d, (8.16) |
| 6 | 122.88 | 6.95, dd, (2.04, 8.16) |
| 7 | 147.06 | 7.55, d, (15.88) |
| 8 | 115.55 | 6.23, d, (15.88) |
| 9 | 171.08 | - |
| SL. No | Target Proteins | Binding Energy | Ligand Efficiency | Intermole Energy | Ligand Atoms (Ring) | Docked Amino Acid Residue (Bond Length) |
|---|---|---|---|---|---|---|
| 1. | Cat-K | −6.01 | −0.6 | −6.55 | C1-OH C1-OH | LYS`17/HZ2 (1.7 Å) LYS`181/HZ2 (2.3 Å) |
| 2 | Cat-B | −4.43 | −0.44 | −4.97 | C1-OH C1-OH C1-OH C1-OH | ASN`222/OD1 (2.5 Å) THR`223/HN (2.2 Å) THR`223/HG1 (1.7 Å) ILE`20/O (3.2 Å) |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| MMP-9 | GTGCTGGGCTGCTGCTTTGCTG | GTCGCCCTCAAAGGTTTGGAAT |
| TRAP | GGTCAGCAGCTCCCTAGAAG | GGAGTGGGAGCCATATGATTT |
| OCN | AGCAAAGGTGCAGCCTTTGT | GCGCCTGGGTCTCTTCACT |
| OPN | ACATCCAGTACCCTGATGCTACAG | TGGCCTTGTATGCACCATTC |
| Cath-K | GCCAGGATGAAAGTTGTATG | CAGGCGTTGTTCTTATTCC |
| Cath-B | GGTTGCAGACCGTACTCCAT | GGAACTGCATCCAAAATGCT |
| BMP-2 | TGCACCAAGATGAACAGC | GTGCCACGATCCAGTCATTC |
| GAPDH | GTATTGGGCGCCTGGTCACC | CGCTCCTGGAGATGGTGATGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, H.-I.M.; Darrag, H.M.; Alhajhoj, M.R.; Khalil, H.E. Biomolecule from Trigonella stellata from Saudi Flora to Suppress Osteoporosis via Osteostromal Regulations. Plants 2020, 9, 1610. https://doi.org/10.3390/plants9111610
Ibrahim H-IM, Darrag HM, Alhajhoj MR, Khalil HE. Biomolecule from Trigonella stellata from Saudi Flora to Suppress Osteoporosis via Osteostromal Regulations. Plants. 2020; 9(11):1610. https://doi.org/10.3390/plants9111610
Chicago/Turabian StyleIbrahim, Hairul-Islam Mohamed, Hossam M. Darrag, Mohammed Refdan Alhajhoj, and Hany Ezzat Khalil. 2020. "Biomolecule from Trigonella stellata from Saudi Flora to Suppress Osteoporosis via Osteostromal Regulations" Plants 9, no. 11: 1610. https://doi.org/10.3390/plants9111610