Physiological Integration Increases Sexual Reproductive Performance of the Rhizomatous Grass Hierochloe glabra

Abstract
:1. Introduction
2. Results
2.1. Morphological Characteristics
2.2. Biomass Allocation
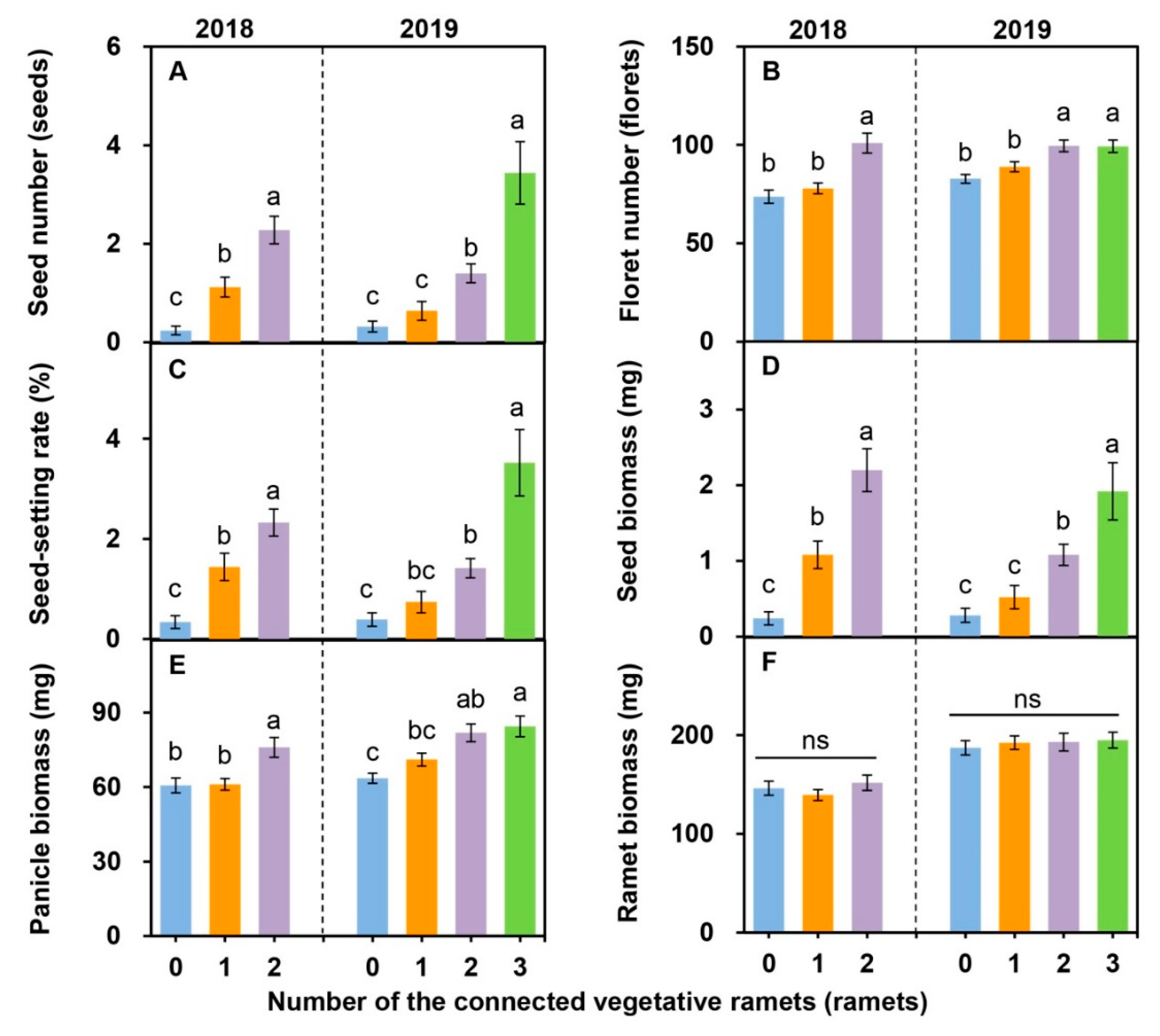
2.3. Sexual Reproductive Characteristics
2.4. Relationships between Soil Physicochemical Properties and Sexual Reproductive Characteristics as Well as Growth Characteristics of Vegetative Ramets
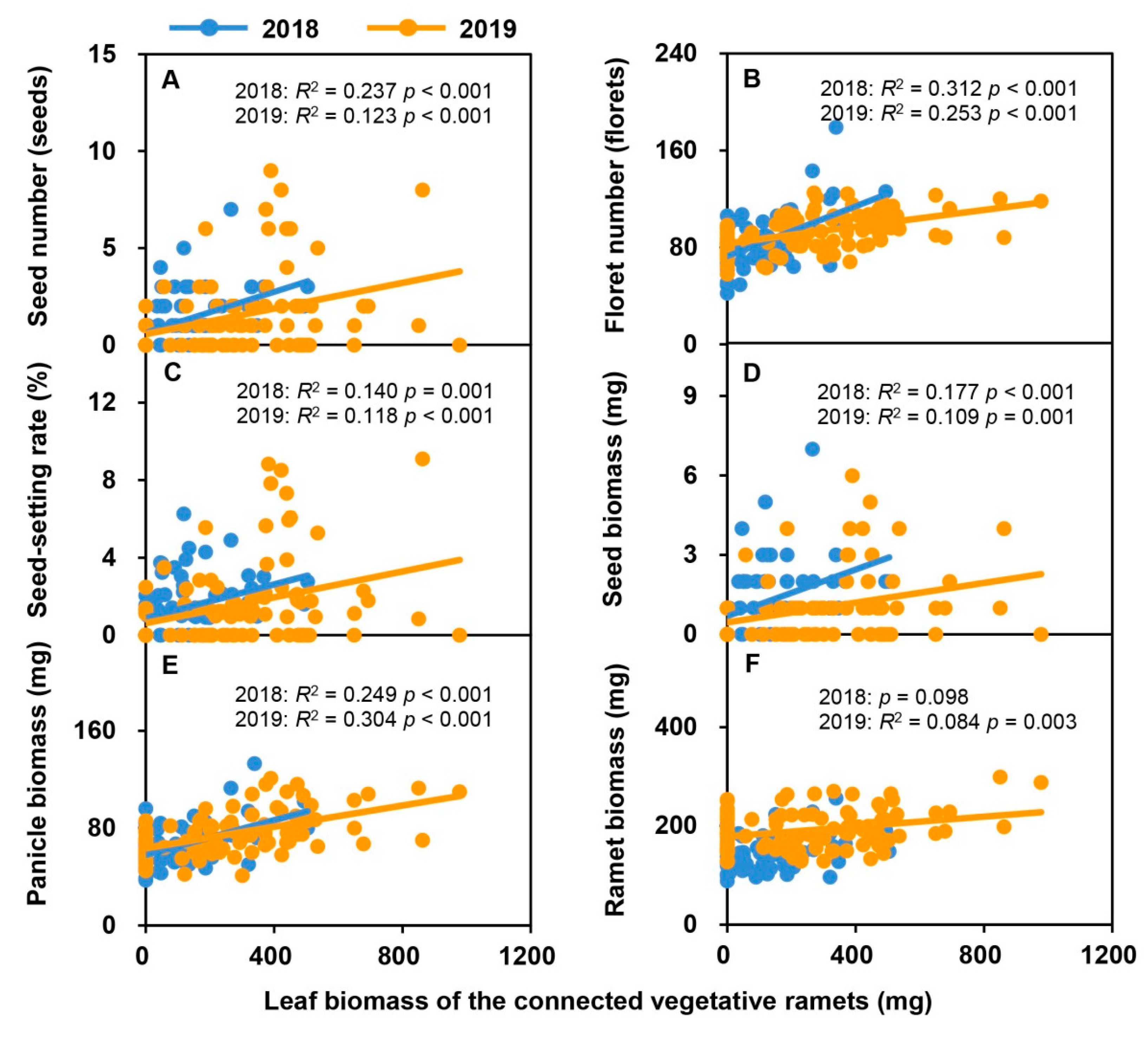
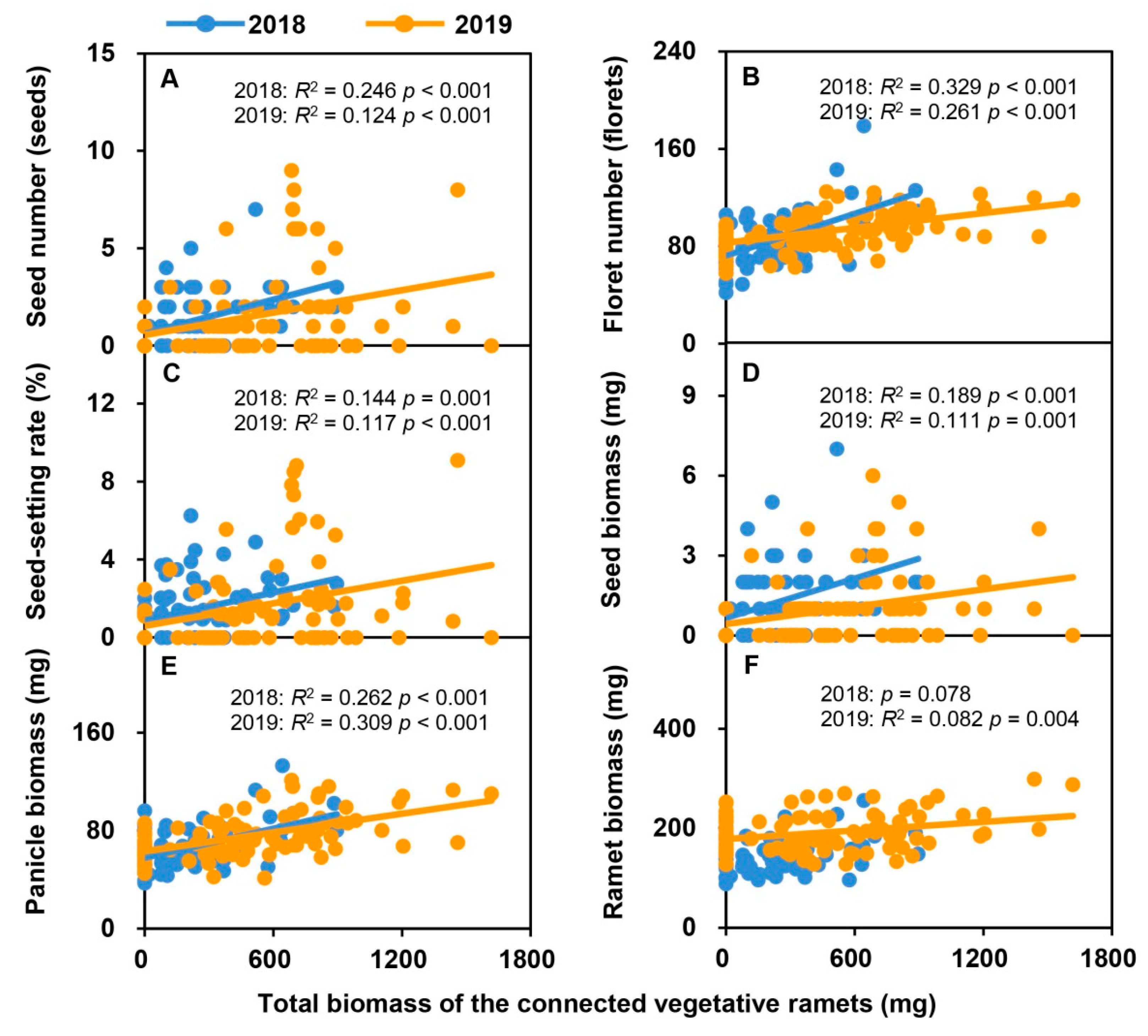
2.5. Relationships between Sexual Reproductive Characteristics and the Leaf Biomass and Total Biomass of Vegetative Ramets Connected to Tillering Nodes
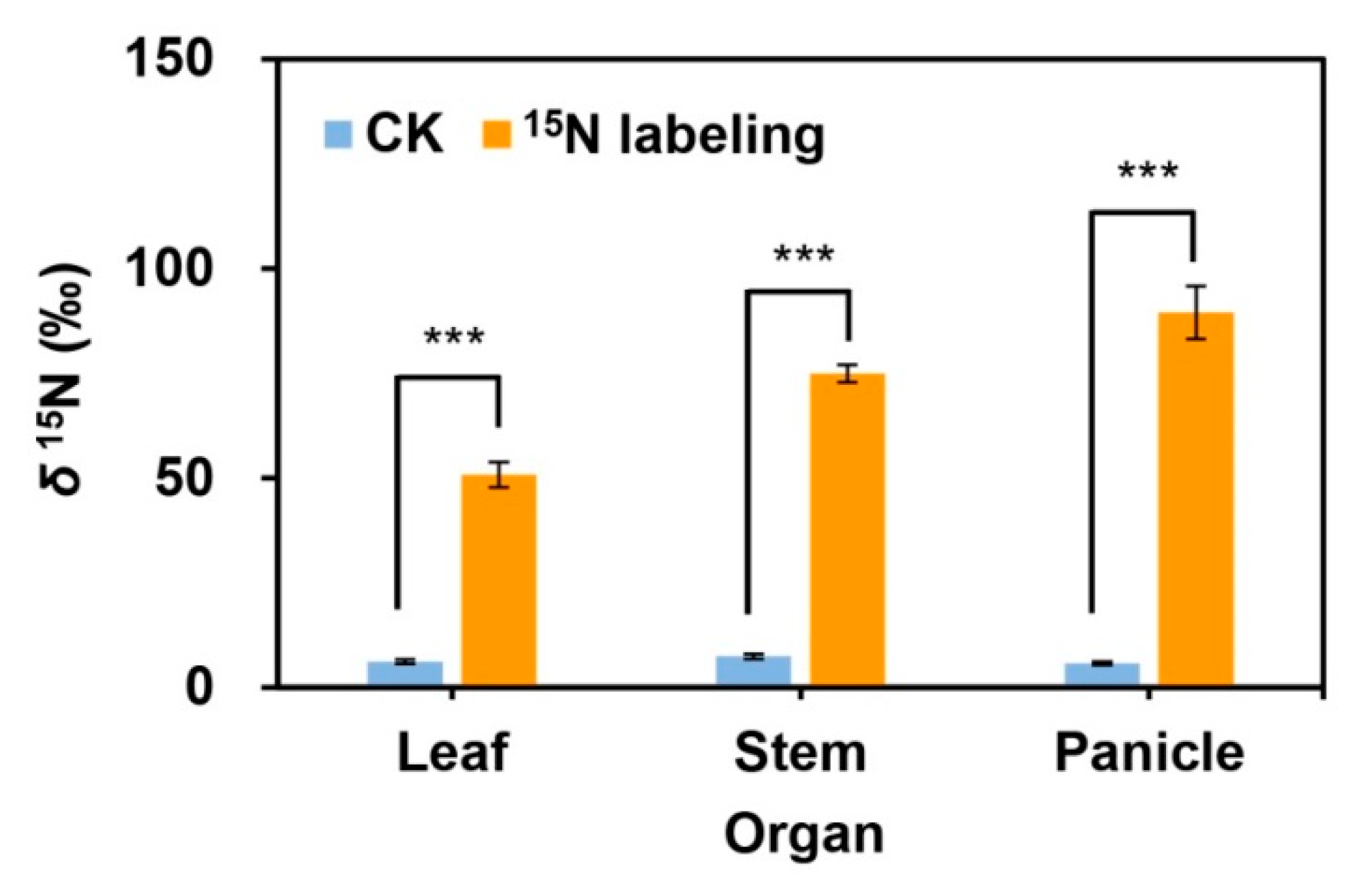
2.6. Transfer of 15N among Ramets
3. Discussion
3.1. Biomass Allocation
3.2. Vegetative Ramets Connected to Tillering Nodes Positively Affect Sexual Reproductive Performance
3.3. The Causes of H. glabra Being a Companion Species Rather than a Dominant Species in the Climax Plant Community
4. Materials and Methods
4.1. Study Area
4.2. Study Species
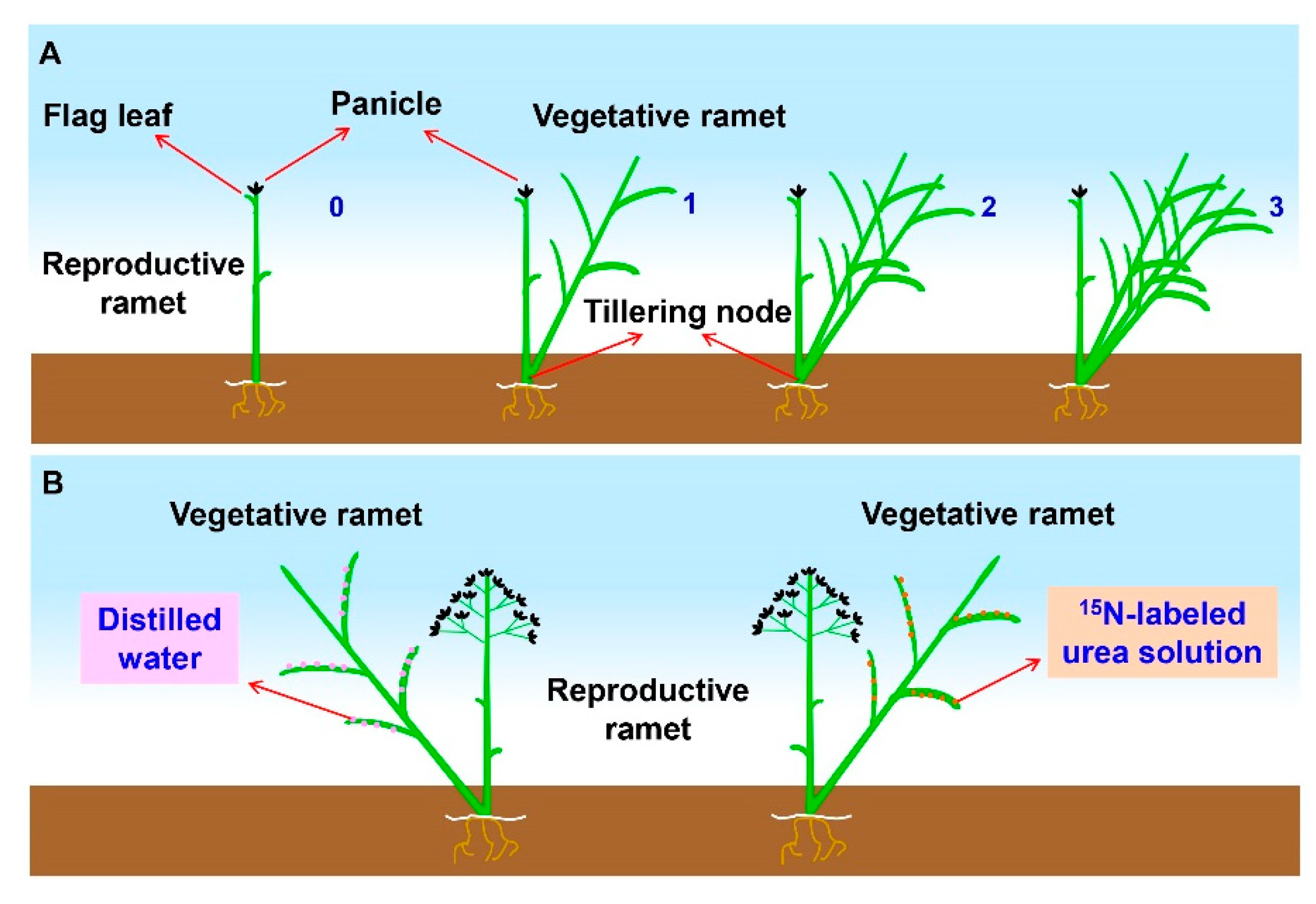
4.3. Experimental Design
4.4. Morphological Characteristics and Biomass Allocation
4.5. Sexual Reproductive Characteristics
4.6. Physicochemical Properties of the Soil
4.7. Stable Isotope Labeling
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
References
- Jackson, R.B.; Caldwell, M.M. Geostatistical patterns of soil heterogeneity around individual perennial plants. J. Ecol. 1993, 81, 683–692. [Google Scholar] [CrossRef]
- Wang, Y.J.; Müller-Schärer, H.; Van Kleunen, M.; Cai, A.M.; Zhang, P.; Yan, R.; Dong, B.C.; Yu, F.H. Invasive alien plants benefit more from physiological integration in heterogeneous environments than natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alpert, P. Physiological integration in Fragaria chiloensis differs between populations: Ramets from grassland are selfish. Oecologia 1999, 120, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, D.; Xing, F.; Liu, J.; Wang, L. Combined effects of resource heterogeneity and simulated herbivory on plasticity of physiological integration in a rhizomatous perennial herb. Plant Biol. 2014, 16, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Ashmun, J.W.; Thomas, R.J.; Pitelka, L.F. Translocation of photoassimilates between sister ramets in two rhizomatous forest herbs. Ann. Bot. 1982, 49, 403–415. [Google Scholar] [CrossRef]
- Alpert, P. Nutrient sharing in natural clonal fragments of Fragaria chiloensis. J. Ecol. 1996, 84, 395–406. [Google Scholar] [CrossRef]
- Evans, J.P.; Cain, M.L. A spatially explicit test of foraging behavior in a clonal plant. Ecology 1995, 76, 1147–1155. [Google Scholar] [CrossRef]
- Stuefer, J.F. Two types of division of labour in clonal plants: Benefits, costs and constraints. Perspect. Plant Ecol. 1998, 1, 47–60. [Google Scholar] [CrossRef]
- Duchoslavová, J.; Jansa, J. The direction of carbon and nitrogen fluxes between ramets in Agrostis stolonifera changes during ontogeny under simulated competition for light. J. Exp. Bot. 2018, 69, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Brewer, J.S.; Bertness, M.D. Disturbance and intraspecific variation in the clonal morphology of salt marsh perennials. Oikos 1996, 77, 107–116. [Google Scholar] [CrossRef]
- Alpert, P.; Holzapfel, C.; Slominski, C. Differences in performance between genotypes of Fragaria chiloensis with different degrees of resource sharing. J. Ecol. 2003, 91, 27–35. [Google Scholar] [CrossRef]
- Yu, F.H.; Chen, Y.F.; Dong, M. Physiological integration enhances survival and performance of Potentilla anserina, suffering from partial sand burial on Ordos Plateau, China. Evol. Ecol. 2002, 15, 303–318. [Google Scholar] [CrossRef]
- Song, Y.B.; Yu, F.H.; Keser, L.H.; Dawson, W.; Fischer, M.; Dong, M.; Van Kleunen, M. United we stand, divided we fall: A meta-analysis of experiments on physiological integration and its relationship to invasiveness. Oecologia 2013, 171, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.H.; Liu, J.; Dong, M. Ecological consequences of physiological integration in plants. Front. Plant Sci. 2016, 7, 770. [Google Scholar] [CrossRef] [Green Version]
- Richards, A.J. Plant Breeding Systems; George Allen and Unwin: London, UK, 1986. [Google Scholar]
- Cook, R.E. Growth and development in clonal plant populations. In Population Biology and Evolution of Clonal Organisms; Jackson, J.B.C., Buss, L.W., Cook, R.E., Eds.; Yale University Press: New Haven, CT, USA, 1985. [Google Scholar]
- Eriksson, O. Seedling dynamics and life histories in clonal plants. Oikos 1989, 55, 231–238. [Google Scholar] [CrossRef]
- Maynard Smith, J. The origin and maintenance of sex. In Group Selection; Williams, G.C., Ed.; Aldine-Atherton: Chicago, IL, USA, 1971; pp. 163–175. [Google Scholar]
- Eckert, C.G. The loss of sex in clonal plants. Evol. Ecol. 2001, 15, 501–520. [Google Scholar] [CrossRef]
- Xiao, Y.; Tang, J.; Qing, H.; Zhou, C.; An, S. Effects of salinity and physiological integration on growth and sexual reproduction of the invasive grass Spartina alterniflora. Flora 2011, 206, 736–741. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Fischer, M.; Schmid, B. Clonal integration in Ranunculus reptans: By-product or adaptation? J. Evol. Biol. 2000, 13, 237–248. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Nuruzzaman, M.; Yamamoto, Y.; Nitta, Y.; Yoshida, T.; Miyazaki, A. Varietal differences in tillering ability of fourteen japonica and indica rice varieties. Soil Sci. Plant Nutr. 2000, 46, 381–391. [Google Scholar] [CrossRef]
- Ao, H.; Peng, S.; Zou, Y.; Tang, Q.; Visperas, R.M. Reduction of unproductive tillers did not increase the grain yield of irrigated rice. Field Crops Res. 2010, 116, 108–115. [Google Scholar] [CrossRef]
- Thorne, G.N.; Wood, D.W. The fate of carbon in dying tillers of winter wheat. J. Agric. Sci. 1987, 108, 515–522. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Gu, Z.Z.; Wang, A.B.; Zhu, X.K.; Guo, W.S. Effects of removing unproductive tillers on grain yield and quality of wheat. Jiangsu Agric. Sci. 2007, 4, 18–20. [Google Scholar] [CrossRef]
- Avolio, M.L.; Forrestel, E.J.; Chang, C.C.; La Pierre, K.J.; Burghardt, K.T.; Smith, M.D. Demystifying dominant species. New Phytol. 2019, 223, 1106–1126. [Google Scholar] [CrossRef]
- Yang, Y.F.; Zhu, T.C. Plant Ecology; Higher Education Press: Beijing, China, 2011. [Google Scholar]
- Wang, J.Y.; Abdullah, I.; Xu, T.T.; Zhu, W.Y.; Gao, Y.; Wang, L. Effects of mowing disturbance and competition on spatial expansion of the clonal plant Leymus chinensis into saline-alkali soil patches. Environ. Exp. Bot. 2019, 168, 103890. [Google Scholar] [CrossRef]
- Guo, L.H.; Yang, Y.F.; Zhang, B.T. Vegetative propagation ability and growth regulation of Hierochloe glabra experimental clones on the Songnen Plain of China. Acta Pratacult. Sin. 2004, 13, 57–61. [Google Scholar] [CrossRef]
- Gao, Y.; Xing, F.; Jin, Y.J.; Nie, D.D.; Wang, Y. Foraging responses of clonal plants to multi-patch environmental heterogeneity: Spatial preference and temporal reversibility. Plant Soil 2012, 359, 137–147. [Google Scholar] [CrossRef]
- Bian, R.J.; Nie, D.D.; Xing, F.; Zhou, X.L.; Gao, Y.; Bai, Z.J.; Liu, B. Adaptational significance of variations in DNA methylation in clonal plant Hierochloe glabra (Poaceae) in heterogeneous habitats. Aust. J. Bot. 2013, 61, 274–282. [Google Scholar] [CrossRef]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Weiner, J.; Campbell, L.G.; Pino, J.; Echarte, L. The allometry of reproduction within plant populations. J. Ecol. 2009, 97, 1220–1233. [Google Scholar] [CrossRef]
- Xie, X.F.; Hu, Y.K.; Pan, X.; Liu, F.H.; Song, Y.B.; Dong, M. Biomass allocation of stoloniferous and rhizomatous plant in response to resource availability: A phylogenetic meta-analysis. Front. Plant Sci. 2016, 7, 603. [Google Scholar] [CrossRef] [Green Version]
- Gifford, R.M.; Thorne, J.H.; Hitz, W.D.; Giaquinta, R.T. Crop productivity and photoassimilate partitioning. Science 1984, 225, 801–808. [Google Scholar] [CrossRef]
- Watson, M.A. Developmental constraints: Effect on population growth and patterns of resource allocation in a clonal plant. Am. Nat. 1984, 123, 411–426. [Google Scholar] [CrossRef]
- Wang, L.Y.; Wang, H.X.; He, C.G.; Sheng, L.X.; Tang, Z.H. An irreversible division of labor through a sexually dependent system in the clonal plant Iris laevigata (Iridaceae). Ecosphere 2017, 8, e01757. [Google Scholar] [CrossRef] [Green Version]
- Klimešová, J.; Nobis, M.P.; Herben, T. Links between shoot and plant longevity and plant economics spectrum: Environmental and demographic implications. Perspect. Plant Ecol. 2016, 22, 55–62. [Google Scholar] [CrossRef]
- Yuan, J.H.; Li, H.Y.; Yang, Y.F. The compensatory tillering in the forage grass Hordeum brevisubulatum after simulated grazing of different severity. Front. Plant Sci. 2020, 11, 792. [Google Scholar] [CrossRef]
- Sheng, J.; Zhu, Y.; Li, H.Y.; Yang, Y.F. Comparison of module structures of Hierochloe glabra populations in heterogeneous habitats on the Songnen Plain of China. Chin. J. Grassl. 2018, 40, 36–42. [Google Scholar] [CrossRef]
- Yang, Y.F.; Zhu, L. Analysis of winter dormancy characters of vegetative forms in fifteen perennial grass species populations on the Songnen Plain of China. Acta Pratacult. Sin. 1994, 3, 26–31. [Google Scholar]
- Yang, Y.F.; Li, J.D. Vegetative propagation characters of clonal populations of Hierochloe glabra on the Songnen Plain of China. Chin. J. Appl. Ecol. 1997, 8, 571–574. [Google Scholar]
- Abrahamson, W.G. Demography and vegetative reproduction. In Demography and Evolution in Plant Populations; Solbrig, O.T., Ed.; Blackwell: Oxford, UK, 1980; pp. 89–106. [Google Scholar]
- Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Thompson, F.L.; Eckert, C.G. Trade-offs between sexual and clonal reproduction in an aquatic plant: Experimental manipulations vs. phenotypic correlations. J. Evol. Biol. 2004, 17, 581–592. [Google Scholar] [CrossRef]
- Van Drunen, W.E.; Dorken, M.E. Trade-offs between clonal and sexual reproduction in Sagittaria latifolia (Alismataceae) scale up to affect the fitness of entire clones. New Phytol. 2012, 196, 606–616. [Google Scholar] [CrossRef]
- Reekie, E.G. Resource allocation, trade-offs, and reproductive effort in plants. In Life History Evolution in Plants; Vuorisalo, T.O., Mutikainen, P.K., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 1999; pp. 173–193. [Google Scholar]
- Delph, L.F.; Lu, Y.; Jayne, L.D. Patterns of resource allocation in a dioecious Carex (Cyperaceae). Am. J. Bot. 1993, 80, 607–615. [Google Scholar] [CrossRef]
- Cline, M.G. Concepts and terminology of apical dominance. Am. J. Bot. 1997, 84, 1064–1069. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, T.; Seiwa, K.; Nishiwaki, A. Effects of resource heterogeneity on nitrogen translocation within clonal fragments of Sasa palmata: An isotopic (15N) assessment. Ann. Bot. 2006, 98, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Pinno, B.D.; Wilson, S.D. Nitrogen translocation between clonal mother and daughter trees at a grassland-forest boundary. Plant Ecol. 2014, 215, 347–354. [Google Scholar] [CrossRef]
- Li, B.; Shibuya, T.; Yogo, Y.; Hara, T.; Yokozawa, M. Interclonal differences, plasticity and trade-offs of life history traits of Cyperus esculentus in relation to water availability. Plant Species Biol. 2001, 16, 193–207. [Google Scholar] [CrossRef]
- Guo, J.; Li, H.Y.; Yang, Y.F. Phenotypic plasticity in sexual reproduction based on nutrients supplied from vegetative ramets in a Leymus chinensis population. Front. Plant Sci. 2020, 10, 1681. [Google Scholar] [CrossRef]
- Kuo, P.C. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1987. [Google Scholar]
- Wang, D.; Ba, L. Ecology of meadow steppe in northeast China. Rangel. J. 2008, 30, 247–254. [Google Scholar] [CrossRef]
- Zhang, C.H.; Yang, Y.F. Analysis of quantitative characters on reproductive tillers in Hierochloe glabra population on the Songnen Plain of China. Acta Pratacult. Sin. 2001, 10, 1–7. [Google Scholar] [CrossRef]
- Jia, S.X. Grassland Science; China Agriculture Press: Beijing, China, 2001.
- Elliott, E.T.; Heil, J.W.; Kelly, E.F.; Monger, H.C. Soil structural and other physical properties. In Standard Soil Methods for Long-term Ecological Research; Robertson, G.P., Ed.; Oxford University Press: New York, NY, USA, 1999; pp. 74–85. [Google Scholar]
- Robertson, G.P.; Sollins, P.; Ellis, B.G.; Lajtha, K. Exchangable ions, pH, and cation exchange capacity. In Standard Soil Methods for Long-term Ecological Research; Robertson, G.P., Coleman, D.C., Bledsoe, C.S., Sollins, P., Eds.; Oxford University Press: Oxford, UK, 1999; pp. 106–114. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. A rapid and accurate method for estimating organic carbon in soil. Proc. Indiana Acad. Sci. 1975, 84, 456–462. [Google Scholar]
- Gallaher, R.N.; Weldon, C.O.; Boswell, F.C. A semiautomated procedure for total nitrogen in plant and soil samples. Soil Sci. Soc. Am. J. 1976, 40, 887–889. [Google Scholar] [CrossRef]
- Sommers, L.E.; Nelson, D.W. Determination of total phosphorus in soils: A rapid perchloric acid digestion procedure 1. Soil Sci. Soc. Am. J. 1972, 36, 902–904. [Google Scholar] [CrossRef]
- Sugiura, N. Further analysis of the data by Akaike’s information criterion and the finite corrections. Commun. Stat.-Theory Methods 1978, 7, 13–26. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Type | Reproductive Ramet | Vegetative Ramet |
|---|---|---|---|
| Ramet height (cm) | 32.66 ± 0.86 | 52.86 ± 0.67 *** | |
| Leaf length (cm) | 1st leaf | 1.32 ± 0.11 b | 35.57 ± 0.57 a |
| 2nd leaf | 1.94 ± 0.12 a | 26.85 ± 0.53 b | |
| 3rd leaf | 22.09 ± 0.47 c | ||
| 4th leaf | 16.41 ± 0.64 d | ||
| Leaf width (cm) | 1st leaf | 0.26 ± 0.02 b | 0.93 ± 0.01 a |
| 2nd leaf | 0.36 ± 0.01 a | 0.96 ± 0.02 a | |
| 3rd leaf | 0.87 ± 0.02 b | ||
| 4th leaf | 0.65 ± 0.02 c | ||
| Leaf area (cm2) | 1st leaf | 0.20 ± 0.03 b | 26.41 ± 0.83 a |
| 2nd leaf | 0.43 ± 0.04 a | 22.06 ± 0.53 b | |
| 3rd leaf | 16.58 ± 0.71 c | ||
| 4th leaf | 9.45 ± 0.55 d | ||
| Total | 0.63 ± 0.06 | 74.50 ± 1.98 *** |
| Organ | Reproductive Ramet | Vegetative Ramet | ||
|---|---|---|---|---|
| Biomass (mg) | Percentage (%) | Biomass (mg) | Percentage (%) | |
| Leaf | 16.12 ± 1.02 c | 7.88 ± 0.48 d | 467.00 ± 13.87 a | 61.32 ± 0.72 a |
| Sheath | 57.16 ± 2.49 b | 27.52 ± 0.63 c | 294.84 ± 9.92 b | 38.68 ± 0.72 b |
| Stem | 62.88 ± 2.75 b | 30.31 ± 0.75 b | ||
| Panicle | 70.72 ± 2.58 a | 34.29 ± 0.69 a | ||
| Total | 206.88 ± 6.81 | 100.00 | 761.84 ± 21.34 *** | 100.00 |
| Characteristic | Moisture | Bulk Density | pH | Electric Conductivity | Total Organic C | Total N | Total P |
|---|---|---|---|---|---|---|---|
| Seed number | −0.052 | −0.055 | −0.092 | −0.040 | 0.014 | −0.083 | 0.036 |
| Floret number | −0.085 | −0.142 | −0.023 | −0.006 | 0.036 | −0.042 | −0.088 |
| Seed-setting rate | −0.013 | −0.025 | −0.023 | −0.018 | −0.003 | −0.074 | 0.067 |
| Seed biomass | −0.047 | −0.019 | −0.075 | −0.083 | 0.066 | 0.003 | −0.008 |
| Panicle biomass | −0.152 | −0.152 | −0.076 | 0.072 | 0.075 | −0.040 | −0.041 |
| Reproductive ramet biomass | −0.247 * | −0.068 | −0.119 | 0.094 | 0.050 | −0.027 | −0.025 |
| Leaf biomass of vegetative ramet | −0.022 | −0.058 | −0.070 | −0.076 | 0.099 | −0.033 | −0.136 |
| Vegetative ramet biomass | −0.006 | −0.061 | −0.061 | −0.060 | 0.081 | −0.026 | −0.132 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Li, H.; Yang, Y. Physiological Integration Increases Sexual Reproductive Performance of the Rhizomatous Grass Hierochloe glabra. Plants 2020, 9, 1608. https://doi.org/10.3390/plants9111608
Guo J, Li H, Yang Y. Physiological Integration Increases Sexual Reproductive Performance of the Rhizomatous Grass Hierochloe glabra. Plants. 2020; 9(11):1608. https://doi.org/10.3390/plants9111608
Chicago/Turabian StyleGuo, Jian, Haiyan Li, and Yunfei Yang. 2020. "Physiological Integration Increases Sexual Reproductive Performance of the Rhizomatous Grass Hierochloe glabra" Plants 9, no. 11: 1608. https://doi.org/10.3390/plants9111608