
Holistic Approaches to Plant Stress Alleviation: A Comprehensive Review of the Role of Organic Compounds and Beneficial Bacteria in Promoting Growth and Health



Abstract
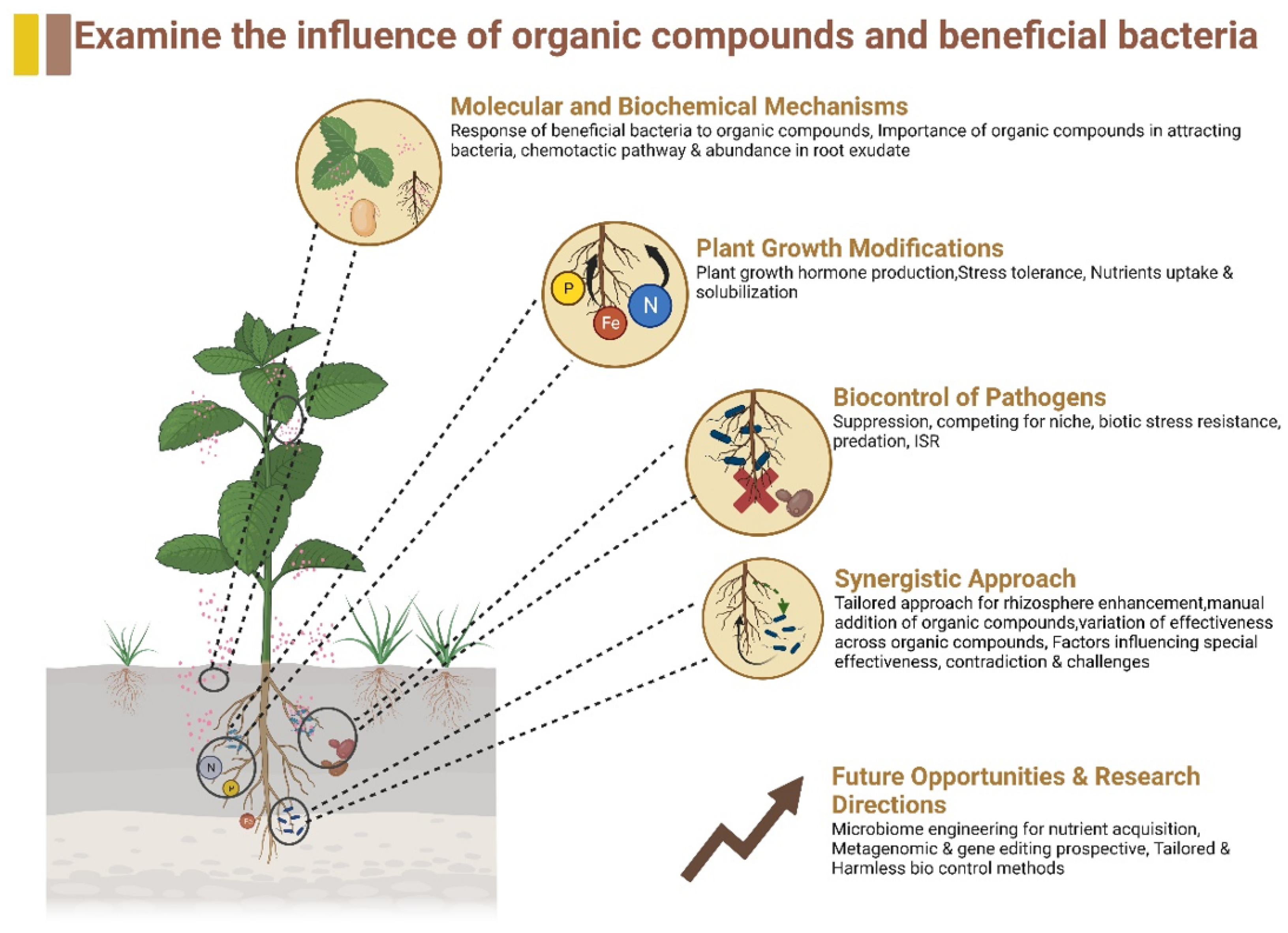
:1. Introduction
1.1. A Brief Overview of the Rhizosphere Microbiome
1.2. The Importance of Organic Compounds and Beneficial Bacteria in Stress Alleviations
2. Molecular and Biochemical Mechanisms
2.1. The Response of Beneficial Bacteria to Organic Compounds
2.2. The Importance of Organic Compounds in Attracting Beneficial Bacteria
2.3. Chemotactic Pathways and Abundance in Root Exudates
3. Plant Growth Modification
Potential of Beneficial Bacteria in Plant Growth Promotion
4. Potential of Beneficial Bacteria in Biocontrol of Pathogens
Strategies Employed by Beneficial Bacteria
5. Synergistic Effects of Organic Compounds and Beneficial Bacteria
5.1. Tailored Approaches for Rhizosphere Enhancement
5.2. Manual Addition of Organic Compounds to Attract Beneficial Bacteria
5.3. Variation in Effectiveness across Organic Compounds
5.4. Factors Influencing Specific Effectiveness
5.5. Contradictions and Challenges
6. Future Opportunities and Research Directions
6.1. Microbiome Engineering for Nutrient Acquisition and Disease Resistance
6.2. Metagenomic and Genome Editing Prospective for Deeper Understanding
6.3. Tailored and Harmless Biocontrol Methods
7. Discussion
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Li, J.; Wang, C.; Liang, W.; Liu, S. Rhizosphere Microbiome: The Emerging Barrier in Plant-Pathogen Interactions. Front. Microbiol. 2021, 12, 772420. [Google Scholar] [CrossRef]
- Yue, H.; Yue, W.; Jiao, S.; Kim, H.; Lee, Y.-H.; Wei, G.; Song, W.; Shu, D. Plant domestication shapes rhizosphere microbiome assembly and metabolic functions. Microbiome 2023, 11, 70. [Google Scholar] [CrossRef]
- Pantigoso, H.A.; Newberger, D.; Vivanco, J.M. The rhizosphere microbiome: Plant-microbial interactions for resource acquisition. J. Appl. Microbiol. 2022, 133, 2864–2876. [Google Scholar] [CrossRef]
- Ling, N.; Wang, T.; Kuzyakov, Y. Rhizosphere bacteriome structure and functions. Nat. Commun. 2022, 13, 836. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef]
- Umar, O.B.; Ranti, L.A.; Abdulbaki, A.S.; Bola, A.; Abdulhamid, A.; Biola, M.; Victor, K. Stresses in plants: Biotic and abiotic. In Current Trends in Wheat Research; IntechOpen: London, UK, 2021. [Google Scholar]
- Nawaz, M.; Sun, J.; Shabbir, S.; Khattak, W.A.; Ren, G.; Nie, X.; Bo, Y.; Javed, Q.; Du, D.; Sonne, C. A review of plants strategies to resist biotic and abiotic environmental stressors. Sci. Total Environ. 2023, 900, 165832. [Google Scholar] [CrossRef]
- Freedman, B. Environmental Science: A Canadian Perspective, 6th ed.; Dalhousie University Libraries: Halifax, NS, Canada, 2018. [Google Scholar]
- Sahu, P.K.; Jayalakshmi, K.; Tilgam, J.; Gupta, A.; Nagaraju, Y.; Kumar, A.; Hamid, S.; Singh, H.V.; Minkina, T.; Rajput, V.D. ROS generated from biotic stress: Effects on plants and alleviation by endophytic microbes. Front. Plant Sci. 2022, 13, 1042936. [Google Scholar] [CrossRef]
- Biswas, S.; Das, R.; Nwe, L.L. Organic Farming to Mitigate Abiotic Stresses under Climate Change Scenario; IntechOpen: London, UK, 2023. [Google Scholar]
- Georgieva, M.; Vassileva, V. Stress Management in Plants: Examining Provisional and Unique Dose-Dependent Responses. Int. J. Mol. Sci. 2023, 24, 5105. [Google Scholar] [CrossRef]
- Norman, C.R.O.; Usselman, M.C.; Noller, C.R. Organic Compound. Available online: https://www.britannica.com/science/organic-compound (accessed on 23 January 2024).
- Kai, M. Diversity and distribution of volatile secondary metabolites throughout Bacillus subtilis isolates. Front. Microbiol. 2020, 11, 559. [Google Scholar] [CrossRef]
- Rowan, D.D. Volatile metabolites. Metabolites 2011, 1, 41–63. [Google Scholar] [CrossRef]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant Volatile Organic Compounds Evolution: Transcriptional Regulation, Epigenetics and Polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7, 11050. [Google Scholar] [CrossRef]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting plant volatile organic compounds (VOCs) in agriculture to improve sustainable defense strategies and productivity of crops. Front. Plant Sci. 2019, 10, 436279. [Google Scholar] [CrossRef]
- Kigathi, R.N.; Weisser, W.W.; Reichelt, M.; Gershenzon, J.; Unsicker, S.B. Plant volatile emission depends on the species composition of the neighboring plant community. BMC Plant Biol. 2019, 19, 58. [Google Scholar] [CrossRef]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef]
- Lyu, D.; Smith, D.L. The root signals in rhizospheric inter-organismal communications. Front. Plant Sci. 2022, 13, 1064058. [Google Scholar] [CrossRef]
- Pantigoso, H.A.; Manter, D.K.; Fonte, S.J.; Vivanco, J.M. Root exudate-derived compounds stimulate the phosphorus solubilizing ability of bacteria. Sci. Rep. 2023, 13, 4050. [Google Scholar] [CrossRef]
- Khan, N.; Humm, E.A.; Jayakarunakaran, A.; Hirsch, A.M. Reviewing and renewing the use of beneficial root and soil bacteria for plant growth and sustainability in nutrient-poor, arid soils. Front. Plant Sci. 2023, 14, 1147535. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Barra Caracciolo, A.; Terenzi, V. Rhizosphere microbial communities and heavy metals. Microorganisms 2021, 9, 1462. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Almeida, O.A.C.; de Araujo, N.O.; Mulato, A.T.N.; Persinoti, G.F.; Sforça, M.L.; Calderan-Rodrigues, M.J.; Oliveira, J.V.d.C. Bacterial volatile organic compounds (VOCs) promote growth and induce metabolic changes in rice. Front. Plant Sci. 2023, 13, 1056082. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Paramasivan, M.; Sahayarayan, J.J. Microbial volatile organic compounds: An alternative for chemical fertilizers in sustainable agriculture development. Microorganisms 2022, 11, 42. [Google Scholar] [CrossRef]
- Smułek, W.; Kaczorek, E. Factors Influencing the Bioavailability of Organic Molecules to Bacterial Cells—A Mini-Review. Molecules 2022, 27, 6579. [Google Scholar] [CrossRef]
- Lohse, M.; Santangeli, M.; Steininger-Mairinger, T.; Oburger, E.; Reemtsma, T.; Lechtenfeld, O.J.; Hann, S. The effect of root hairs on exudate composition: A comparative non-targeted metabolomics approach. Anal. Bioanal. Chem. 2023, 415, 823–840. [Google Scholar] [CrossRef]
- Contreras, F.; Díaz, J.; Rombolà, A.D.; de la Luz Mora, M. Prospecting intercropping between subterranean clover and grapevine as potential strategy for improving grapevine performance. Curr. Plant Biol. 2019, 19, 100110. [Google Scholar] [CrossRef]
- Hazrati, H.; Fomsgaard, I.S.; Kudsk, P. Root-exuded benzoxazinoids: Uptake and translocation in neighboring plants. J. Agric. Food Chem. 2020, 68, 10609–10617. [Google Scholar] [CrossRef]
- Sharma, M.; Saleh, D.; Charron, J.-B.; Jabaji, S. A crosstalk between Brachypodium root exudates, organic acids, and Bacillus velezensis B26, a growth promoting bacterium. Front. Microbiol. 2020, 11, 575578. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, N.; Fu, R.; Liu, Y.; Krell, T.; Du, W.; Shao, J.; Shen, Q.; Zhang, R. Recognition of dominant attractants by key chemoreceptors mediates recruitment of plant growth-promoting rhizobacteria. Environ. Microbiol. 2019, 21, 402–415. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, N.; Huang, Q.; Raza, W.; Li, R.; Vivanco, J.M.; Shen, Q. Organic acids from root exudates of banana help root colonization of PGPR strain Bacillus amyloliquefaciens NJN-6. Sci. Rep. 2015, 5, 13438. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Wang, X.; Qin, Y.; Ren, Z.; Zhao, X. Watermelon Root Exudates Enhance Root Colonization of Bacillus amyloliquefaciens TR2. Curr. Microbiol. 2023, 80, 110. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yang, C.; Mei, X.; Shen, S.; Raza, W.; Shen, Q.; Xu, Y. The effect of organic acids from tomato root exudates on rhizosphere colonization of Bacillus amyloliquefaciens T-5. Appl. Soil Ecol. 2013, 64, 15–22. [Google Scholar] [CrossRef]
- Wen, T.; Yuan, J.; He, X.; Lin, Y.; Huang, Q.; Shen, Q. Enrichment of beneficial cucumber rhizosphere microbes mediated by organic acid secretion. Hortic. Res. 2020, 7. [Google Scholar] [CrossRef]
- Saleh, D.; Sharma, M.; Seguin, P.; Jabaji, S. Organic acids and root exudates of Brachypodium distachyon: Effects on chemotaxis and biofilm formation of endophytic bacteria. Can. J. Microbiol. 2020, 66, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, Q.-F.; Shang, N.; Li, N.; Niu, Q.-H.; Hong, Q.; Huang, X. The enhanced mechanisms of Hansschlegelia zhihuaiae S113 degrading bensulfuron-methyl in maize rhizosphere by three organic acids in root exudates. Ecotoxicol. Environ. Saf. 2021, 223, 112622. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, F.; Zhao, H.-Z.; Lu, J.-S.; Zhao, M.-J.; Hong, Q.; Huang, X. Colonization on cucumber root and enhancement of chlorimuron-ethyl degradation in the rhizosphere by Hansschlegelia zhihuaiae S113 and root exudates. J. Agric. Food Chem. 2018, 66, 4584–4591. [Google Scholar] [CrossRef]
- Tian, J.-H.; Shuang, R.; Yang, G.; Yang, L.; Cai, K.-Z. Wheat straw biochar amendment suppresses tomato bacterial wilt caused by Ralstonia solanacearum: Potential effects of rhizosphere organic acids and amino acids. J. Integr. Agric. 2021, 20, 2450–2462. [Google Scholar] [CrossRef]
- Feng, H.; Fu, R.; Hou, X.; Lv, Y.; Zhang, N.; Liu, Y.; Xu, Z.; Miao, Y.; Krell, T.; Shen, Q.; et al. Chemotaxis of Beneficial Rhizobacteria to Root Exudates: The First Step towards Root-Microbe Rhizosphere Interactions. Int. J. Mol. Sci. 2021, 22, 6655. [Google Scholar] [CrossRef]
- López-Farfán, D.; Reyes-Darias, J.A.; Matilla, M.A.; Krell, T. Concentration Dependent Effect of Plant Root Exudates on the Chemosensory Systems of Pseudomonas putida KT2440. Front. Microbiol. 2019, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Zhang, N.; Du, W.; Zhang, H.; Liu, Y.; Fu, R.; Shao, J.; Zhang, G.; Shen, Q.; Zhang, R. Identification of chemotaxis compounds in root exudates and their sensing chemoreceptors in plant-growth-promoting rhizobacteria Bacillus amyloliquefaciens SQR9. Mol. Plant-Microbe Interact. 2018, 31, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Sabagh, A.E.; Mbarki, S.; Hossain, A.; Iqbal, M.A.; Islam, M.S.; Raza, A.; Llanes, A.; Reginato, M.; Rahman, M.A.; Mahboob, W. Potential role of plant growth regulators in administering crucial processes against abiotic stresses. Front. Agron. 2021, 3, 648694. [Google Scholar] [CrossRef]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef] [PubMed]
- Abhyankar, P.; Gunjal, A.; Kapadnis, B.; Ambade, S. Potential of lactic acid bacteria in plant growth promotion. Bhartiya Krishi Anusandhan Patrika 2021, 36, 326–329. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Duan, J.; DiBernardo, M.; Zetter, E.; Campos-García, J.; Glick, B.R.; Santoyo, G. The Production of ACC Deaminase and Trehalose by the Plant Growth Promoting Bacterium Pseudomonas sp. UW4 Synergistically Protect Tomato Plants Against Salt Stress. Front. Plant Sci. 2019, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Rana, K.L.; Sheikh, I.; Kumar, V.; Yadav, A.N.; Dhaliwal, H.S.; Saxena, A.K. Alleviation of drought stress and plant growth promotion by Pseudomonas libanensis EU-LWNA-33, a drought-adaptive phosphorus-solubilizing bacterium. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 785–795. [Google Scholar] [CrossRef]
- Zubair, M.; Hanif, A.; Farzand, A.; Sheikh, T.M.M.; Khan, A.R.; Suleman, M.; Ayaz, M.; Gao, X. Genetic Screening and Expression Analysis of Psychrophilic Bacillus spp. Reveal Their Potential to Alleviate Cold Stress and Modulate Phytohormones in Wheat. Microorganisms 2019, 7, 337. [Google Scholar] [CrossRef]
- Din, M.; Nelofer, R.; Salman, M.; Khan, F.H.; Khan, A.; Ahmad, M.; Jalil, F.; Din, J.U.; Khan, M. Production of nitrogen fixing Azotobacter (SR-4) and phosphorus solubilizing Aspergillus niger and their evaluation on Lagenaria siceraria and Abelmoschus esculentus. Biotechnol. Rep. 2019, 22, e00323. [Google Scholar] [CrossRef]
- Cozzolino, V.; Monda, H.; Savy, D.; Di Meo, V.; Vinci, G.; Smalla, K. Cooperation among phosphate-solubilizing bacteria, humic acids and arbuscular mycorrhizal fungi induces soil microbiome shifts and enhances plant nutrient uptake. Chem. Biol. Technol. Agric. 2021, 8, 1–18. [Google Scholar] [CrossRef]
- Suleman, M.; Yasmin, S.; Rasul, M.; Yahya, M.; Atta, B.M.; Mirza, M.S. Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLoS ONE 2018, 13, e0204408. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, L.; Ma, Z.; Wang, J. Improved iron acquisition of Astragalus sinicus under low iron-availability conditions by soil-borne bacteria Burkholderia cepacia. J. Plant Interact. 2018, 13, 9–20. [Google Scholar] [CrossRef]
- Zaheer, A.; Malik, A.; Sher, A.; Qaisrani, M.M.; Mehmood, A.; Khan, S.U.; Ashraf, M.; Mirza, Z.; Karim, S.; Rasool, M. Isolation, characterization, and effect of phosphate-zinc-solubilizing bacterial strains on chickpea (Cicer arietinum L.) growth. Saudi J. Biol. Sci. 2019, 26, 1061–1067. [Google Scholar] [CrossRef]
- Kumar, A.; Maleva, M.; Bruno, L.B.; Rajkumar, M. Synergistic effect of ACC deaminase producing Pseudomonas sp. TR15a and siderophore producing Bacillus aerophilus TR15c for enhanced growth and copper accumulation in Helianthus annuus L. Chemosphere 2021, 276, 130038. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Pantigoso, H.A.; Wu, Z.; Vivanco, J.M. Co-inoculation of Bacillus sp. and Pseudomonas putida at different development stages acts as a biostimulant to promote growth, yield and nutrient uptake of tomato. J. Appl. Microbiol. 2019, 127, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-L.; Guo, X.-J.; Huang, X.; Guo, R.-J.; Lu, X.-H.; Li, S.-D.; Zhang, H. The Co-Association of Enterobacteriaceae and Pseudomonas with Specific Resistant Cucumber against Fusarium Wilt Disease. Biology 2023, 12, 143. [Google Scholar] [CrossRef]
- Wu, C.; Wang, F.; Ge, A.; Zhang, H.; Chen, G.; Deng, Y.; Yang, J.; Chen, J.; Ge, T. Enrichment of microbial taxa after the onset of wheat yellow mosaic disease. Agric. Ecosyst. Environ. 2021, 322, 107651. [Google Scholar] [CrossRef]
- Lazcano, C.; Boyd, E.; Holmes, G.; Hewavitharana, S.; Pasulka, A.; Ivors, K. The rhizosphere microbiome plays a role in the resistance to soil-borne pathogens and nutrient uptake of strawberry cultivars under field conditions. Sci. Rep. 2021, 11, 3188. [Google Scholar] [CrossRef]
- Bahroun, A.; Jousset, A.; Mrabet, M.; Mhamdi, R.; Mhadhbi, H. Protists modulate Fusarium root rot suppression by beneficial bacteria. Appl. Soil Ecol. 2021, 168, 104158. [Google Scholar] [CrossRef]
- Rahman, M.M.; Flory, E.; Koyro, H.-W.; Abideen, Z.; Schikora, A.; Suarez, C.; Schnell, S.; Cardinale, M. Consistent associations with beneficial bacteria in the seed endosphere of barley (Hordeum vulgare L.). Syst. Appl. Microbiol. 2018, 41, 386–398. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Yao, G.; Zhu, L.; Lin, J.; Wang, C.; Du, B.; Ding, Y.; Mei, X. Synergistic effect of co-culture rhizosphere Streptomyces: A promising strategy to enhance antimicrobial activity and plant growth-promoting function. Front. Microbiol. 2022, 13, 976484. [Google Scholar] [CrossRef]
- Kushwaha, P.; Srivastava, R.; Pandiyan, K.; Singh, A.; Chakdar, H.; Kashyap, P.L.; Bhardwaj, A.K.; Murugan, K.; Karthikeyan, N.; Bagul, S.Y. Enhancement in plant growth and zinc biofortification of chickpea (Cicer arietinum L.) by Bacillus altitudinis. J. Soil Sci. Plant Nutr. 2021, 21, 922–935. [Google Scholar] [CrossRef]
- Asghar, W.; Kataoka, R. Effect of co-application of Trichoderma spp. with organic composts on plant growth enhancement, soil enzymes and fungal community in soil. Arch. Microbiol. 2021, 203, 4281–4291. [Google Scholar] [CrossRef]
- Tian, T.; Sun, B.; Shi, H.; Gao, T.; He, Y.; Li, Y.; Liu, Y.; Li, X.; Zhang, L.; Li, S. Sucrose triggers a novel signaling cascade promoting Bacillus subtilis rhizosphere colonization. ISME J. 2021, 15, 2723–2737. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.; Zhang, J.; Huang, X.; Huang, Q.; Liu, J.; Yue, W. Dynamics of microbial community composition on grass surfaces in response to glucose and sucrose spraying for enhanced silage quality.2023. arXiv 2023. [Google Scholar] [CrossRef]
- Martins, S.J.; Medeiros, F.H.; Lakshmanan, V.; Bais, H.P. Impact of seed exudates on growth and biofilm formation of Bacillus amyloliquefaciens ALB629 in common bean. Front. Microbiol. 2018, 8, 2631. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Mfarrej, M.F.B.; Rizwan, M.; Hussain, A.; Shahid, M.J.; Wang, X.; Nafees, M.; Waseem, M.; Alharby, H.F. Microbe-citric acid assisted phytoremediation of chromium by castor bean (Ricinus communis L.). Chemosphere 2022, 296, 134065. [Google Scholar] [CrossRef] [PubMed]
- Bowya, T.; Balachandar, D. Harnessing PGPR inoculation through exogenous foliar application of salicylic acid and microbial extracts for improving rice growth. J. Basic Microbiol. 2020, 60, 950–961. [Google Scholar] [CrossRef] [PubMed]
- Jha, Y.; Yadav, K.A.; Mohamed, H.I. Plant growth-promoting bacteria and exogenous phytohormones alleviate the adverse effects of drought stress in pigeon pea plants. Plant Soil 2023, 1–21. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, L.; Zhang, S.; Li, B.; Li, J.; Wang, X.; Zhang, J.; Guan, C.; Ji, J. The combined use of a plant growth promoting Bacillus sp. strain and GABA promotes the growth of rice under salt stress by regulating antioxidant enzyme system, enhancing photosynthesis and improving soil enzyme activities. Microbiol. Res. 2023, 266, 127225. [Google Scholar] [CrossRef] [PubMed]
- Zhitnitsky, D.; Rose, J.; Lewinson, O. The highly synergistic, broad spectrum, antibacterial activity of organic acids and transition metals. Sci. Rep. 2017, 7, 44554. [Google Scholar] [CrossRef] [PubMed]
- Macias-Benitez, S.; Garcia-Martinez, A.M.; Caballero Jimenez, P.; Gonzalez, J.M.; Tejada Moral, M.; Parrado Rubio, J. Rhizospheric organic acids as biostimulants: Monitoring feedbacks on soil microorganisms and biochemical properties. Front. Plant Sci. 2020, 11, 633. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, K.; AlKahtani, M.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The role of plant growth-promoting bacteria in alleviating the adverse effects of drought on plants. Biology 2021, 10, 520. [Google Scholar] [CrossRef] [PubMed]
- Majeed, A.; Muhammad, Z.; Ahmad, H. Plant growth promoting bacteria: Role in soil improvement, abiotic and biotic stress management of crops. Plant Cell Rep. 2018, 37, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Zhang, L.; He, S.Y. Plant-microbe interactions facing environmental challenge. Cell Host Microbe 2019, 26, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Zhao, M.; Yuan, J.; Kowalchuk, G.A.; Shen, Q. Root exudates mediate plant defense against foliar pathogens by recruiting beneficial microbes. Soil Ecol. Lett. 2021, 3, 42–51. [Google Scholar] [CrossRef]
- El Hamss, H.; Kajad, N.; Belabess, Z.; Lahlali, R. Enhancing bioefficacy of Bacillus amyloliquefaciens SF14 with salicylic acid for the control of the postharvest citrus green mould. Plant Stress 2023, 7, 100144. [Google Scholar] [CrossRef]
- Li, S.; Xu, C.; Wang, J.; Guo, B.; Yang, L.; Chen, J.; Ding, W. Cinnamic, myristic and fumaric acids in tobacco root exudates induce the infection of plants by Ralstonia solanacearum. Plant Soil 2017, 412, 381–395. [Google Scholar] [CrossRef]
- Wu, K.; Su, L.; Fang, Z.; Yuan, S.; Wang, L.; Shen, B.; Shen, Q. Competitive use of root exudates by Bacillus amyloliquefaciens with Ralstonia solanacearum decreases the pathogenic population density and effectively controls tomato bacterial wilt. Sci. Hortic. 2017, 218, 132–138. [Google Scholar] [CrossRef]
- Arif, I.; Batool, M.; Schenk, P.M. Plant microbiome engineering: Expected benefits for improved crop growth and resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Lau, S.-E.; Teo, W.F.A.; Teoh, E.Y.; Tan, B.C. Microbiome engineering and plant biostimulants for sustainable crop improvement and mitigation of biotic and abiotic stresses. Discov. Food 2022, 2, 9. [Google Scholar] [CrossRef]
- Singh, A.; Mazahar, S.; Chapadgaonkar, S.S.; Giri, P.; Shourie, A. Phyto-microbiome to mitigate abiotic stress in crop plants. Front. Microbiol. 2023, 14, 1210890. [Google Scholar] [CrossRef]
- Ali, S.; Tyagi, A.; Mir, R.A.; Rather, I.A.; Anwar, Y.; Mahmoudi, H. Plant beneficial microbiome a boon for improving multiple stress tolerance in plants. Front. Plant Sci. 2023, 14, 1266182. [Google Scholar] [CrossRef]
- Shah, A.; Nazari, M.; Antar, M.; Msimbira, L.A.; Naamala, J.; Lyu, D.; Rabileh, M.; Zajonc, J.; Smith, D.L. PGPR in agriculture: A sustainable approach to increasing climate change resilience. Front. Sustain. Food Syst. 2021, 5, 667546. [Google Scholar] [CrossRef]
- Lindemann, S.R. A piece of the pie: Engineering microbiomes by exploiting division of labor in complex polysaccharide consumption. Curr. Opin. Chem. Eng. 2020, 30, 96–102. [Google Scholar] [CrossRef]
- Yang, W.; Gong, T.; Wang, J.; Li, G.; Liu, Y.; Zhen, J.; Ning, M.; Yue, D.; Du, Z.; Chen, G. Effects of compound microbial fertilizer on soil characteristics and yield of wheat (Triticum aestivum L.). J. Soil Sci. Plant Nutr. 2020, 20, 2740–2748. [Google Scholar] [CrossRef]
- Hussain, M.; Shakoor, N.; Adeel, M.; Ahmad, M.A.; Zhou, H.; Zhang, Z.; Xu, M.; Rui, Y.; White, J.C. Nano-enabled plant microbiome engineering for disease resistance. Nano Today 2023, 48, 101752. [Google Scholar] [CrossRef]
- Awla, H.K.; Kadir, J.; Othman, R.; Rashid, T.S.; Hamid, S.; Wong, M.-Y. Plant growth-promoting abilities and 952biocontrol efficacy of Streptomyces sp. UPMRS4 against Pyricularia oryzae. Biological Control 2017, 112, 55-95363. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Metagenomics methods for the study of plant-associated microbial communities: A review. J. Microbiol. Methods 2020, 170, 105860. [Google Scholar] [CrossRef] [PubMed]
- Urumbil, S.K.; Anilkumar, M. Metagenomic insights into plant growth promoting genes inherent in bacterial endophytes of Emilia sonchifolia (Linn.) DC. Plant Sci. Today 2021, 8, 6–16. [Google Scholar] [CrossRef]
- Esquerra-Ruvira, B.; Baquedano, I.; Ruiz, R.; Fernandez, A.; Montoliu, L.; Mojica, F.J. Identification of the EH CRISPR-Cas9 system on a metagenome and its application to genome engineering. Microb. Biotechnol. 2023, 16, 1505–1523. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12, 634796. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Ahmed, A.I.; Mahmood, M.; El-Tahan, A.M.; Ebrahim, A.A.; El-Mageed, A.; Taia, A. Plant growth-promoting microorganisms as biocontrol agents of plant diseases: Mechanisms, challenges and future perspectives. Front. Plant Sci. 2022, 13, 923880. [Google Scholar] [CrossRef]
- Pirttilä, A.M.; Mohammad Parast Tabas, H.; Baruah, N.; Koskimäki, J.J. Biofertilizers and biocontrol agents for agriculture: How to identify and develop new potent microbial strains and traits. Microorganisms 2021, 9, 817. [Google Scholar] [CrossRef] [PubMed]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef]
- Lee, J.; Kim, S.; Jung, H.; Koo, B.-K.; Han, J.A.; Lee, H.-S. Exploiting bacterial genera as biocontrol agents: Mechanisms, interactions and applications in sustainable agriculture. J. Plant Biol. 2023, 66, 485–498. [Google Scholar] [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Francés, J.; Roselló, G.; Montesinos, E. Bacteria as biological control agents of plant diseases. Microorganisms 2022, 10, 1759. [Google Scholar] [CrossRef]
- Sare, A.R.; Jijakli, M.H.; Massart, S. Microbial ecology to support integrative efficacy improvement of biocontrol agents for postharvest diseases management. Postharvest Biol. Technol. 2021, 179, 111572. [Google Scholar] [CrossRef]
- Xu, X.; Jeger, M. More ecological research needed for effective biocontrol of plant pathogens. In How Research Can Stimulate the Development of Commercial Biological Control Against Plant Diseases; Springer: Berlin/Heidelberg, Germany, 2020; pp. 15–30. [Google Scholar]
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological Control of Plant Diseases: An Evolutionary and Eco-Economic Consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- James, S.; Collins, F.H.; Welkhoff, P.A.; Emerson, C.; Godfray, H.C.J.; Gottlieb, M.; Greenwood, B.; Lindsay, S.W.; Mbogo, C.M.; Okumu, F.O.; et al. Pathway to Deployment of Gene Drive Mosquitoes as a Potential Biocontrol Tool for Elimination of Malaria in Sub-Saharan Africa: Recommendations of a Scientific Working Group. Am. J. Trop. Med. Hyg. 2018, 98, 1–49. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Bacteria | Organic Compound in Media a | Mechanism | Plant | Reference |
|---|---|---|---|---|
| Bacillus velezensis | Malic, Succinic, Citric, Fumaric | Chemotactic response | Brachypodium distachyon | [33] |
| Bacillus velezensis | Malic, Fumaric, Gluconic, Glyceric | Chemotactic response | Cucumus sativus L. | [34] |
| Bacillus amyloliquefaciens Bacillus subtilis | Fumaric | Chemotactic response Biofilm formation | Cucumus sativus L., Musa acuminata | [35] |
| Bacillus amyloliquefaciens | Malic, Oxalic, Fumaric | Chemotactic response Biofilm formation | Musa acuminata | [36] |
| Bacillus amyloliquefaciens | Malic, Citric, succinic, Fumaric | Chemotactic response Swarming motility Biofilm formation | Citrullus lanatus | [37] |
| Bacillus amyloliquefaciens | Malic, Citric, succinic, Fumaric | Chemotactic response Biofilm formation | Lycopersicon esculentum | [38] |
| Comamonadaceae | Citric, Pyruvate, Succinic, Fumarate | Enhance colonization in roots | Cucumus sativus L. | [39] |
| Microbacterium | Citric | Biofilm formation | Brachypodium distachyon | [40] |
| Hansschlegelia zhihuaiae | L-malic, Tarteric, Fumaric | Degrade herbicides | Zea mays L. Cucumus sativus L. | [41,42] |
| Mechanism | Beneficial Bacteria | Outcome | Plant | Reference |
|---|---|---|---|---|
| Production of hormones | Leuconostoc sp. | Promote plant growth | Punica granatum L. | [49] |
| Stress tolerance | Pseudomonas sp. UW4 | Reduce salt stress | Lycopersicon esculentum | [50] |
| Pseudomonas libanensis | Response to drought stress | Triticum aestivum | [51] | |
| Bacillus sp. CJCL2, RJGP41 | Response to cold stress | Triticum aestivum | [52] | |
| N fixation | Azotobacter | Increase plant height, leaf length, fruit size | Lagenaria siceraria Abelmoschus esculentus | [53] |
| P solubilization | B. amyloliquefaciens Pseudomonas sp. | Increase plant biomass | Zea mays L. Triticum aestivum | [54,55] |
| Iron chelation | Burkholderia cerpia | Improve flavin metabolic pathway | Astragalus sinicus | [56] |
| Zn uptake | Bacillus sp. Pseudomonas sp. | Increase in grain yield | Cicer arietinum L. | [57] |
| Cu uptake | Bacillus aerophilus | Improved plant growth | Helianthus annuus | [58] |
| Mn uptake | Bacillus pumilus | High fruit yield Improved plant growth | Solamum lycopersicum | [59] |
| Mechanism | Beneficial Bacteria | Pathogen | Plant | Reference |
|---|---|---|---|---|
| Suppression | Enterobacteriaceae | Fusarium oxysporum f. sp. cucumerinum(Foc) | Cucumus sativus L. | [60] |
| Compete for niche | Streptomyces, Stenotrophomonas, Bradyrhizobium, Sphingomonas Bacillus | Chinese wheat mosaic virus | Triticum aestivum L. | [61] |
| Resistance to biotic stress | Arthrobacter, Nocardioides Gaiella Burkholderia Pseudomonas | Verticillium dahliae Macrophomina phaseolina | Fragaria × ananassa | [62] |
| Predation (with bacterivorous protists) | Rahnella aquatilis P. yamanorum | Fusarium solani | Vicia faba | [63] |
| ISR | Paenibacillus, Pantoea, Pseudomonas spp. | Blumeria graminis | Hordeum vulgare L. | [64] |
| Type | Organic Compound | Beneficial Bacteria | Plant | Mechanism | Reference |
|---|---|---|---|---|---|
| Sugars | Sucrose | B. subtilis | Solamum lycopersicum | IRS against B. cinera Suppression against F. oxyparum | [68] |
| Glucose Sucrose | Rhizosphere microbiome | Lolium multiflorum | Enhance silage quality | [69] | |
| Organic acids | Malic | B. amyloliquefaciens | Phaseolus vulgaris | Higher growth promotion and drought tolerance | [70] |
| Citric | S. aureus | Ricinus communis L. | Phytoremediation of Cr | [71] | |
| Hormones | Salicylic | C. glutamicum S. cerevisiae | Oryza sativa | Abundant of beneficial bacteria in roots | [72] |
| IAA | P. aeruginosa B. megaterium | Cajanus cajan L. | Promote osmolyte synthesis, increasing photosynthetic activity and mineral uptake | [73] | |
| Amino acids | GABA | B. pumilus | Oryza sativa | Increase photosynthetic efficiency, chlorophyll accumulation, and anti-oxidant levels | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hathurusinghe, S.H.K.; Azizoglu, U.; Shin, J.-H. Holistic Approaches to Plant Stress Alleviation: A Comprehensive Review of the Role of Organic Compounds and Beneficial Bacteria in Promoting Growth and Health. Plants 2024, 13, 695. https://doi.org/10.3390/plants13050695
Hathurusinghe SHK, Azizoglu U, Shin J-H. Holistic Approaches to Plant Stress Alleviation: A Comprehensive Review of the Role of Organic Compounds and Beneficial Bacteria in Promoting Growth and Health. Plants. 2024; 13(5):695. https://doi.org/10.3390/plants13050695
Chicago/Turabian StyleHathurusinghe, Sandamali Harshani Kumari, Ugur Azizoglu, and Jae-Ho Shin. 2024. "Holistic Approaches to Plant Stress Alleviation: A Comprehensive Review of the Role of Organic Compounds and Beneficial Bacteria in Promoting Growth and Health" Plants 13, no. 5: 695. https://doi.org/10.3390/plants13050695