
Evaluation of Tomato Germplasm against Tomato Brown Rugose Fruit Virus and Identification of Resistance in Solanum pimpinellifolium
 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Primary Screening of USDA and TGRC Tomato Core Germplasm Collections for Resistance to ToBRFV
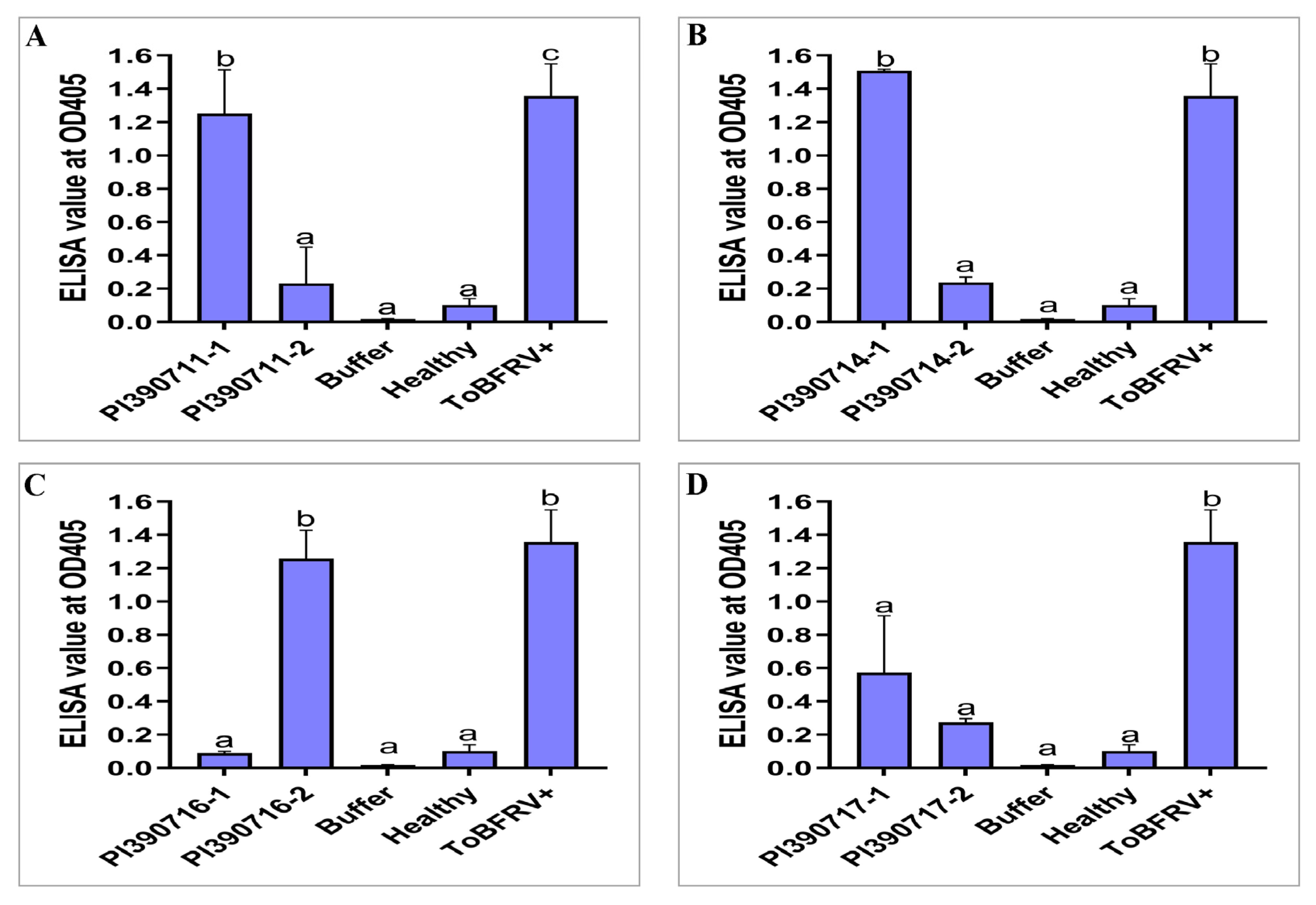
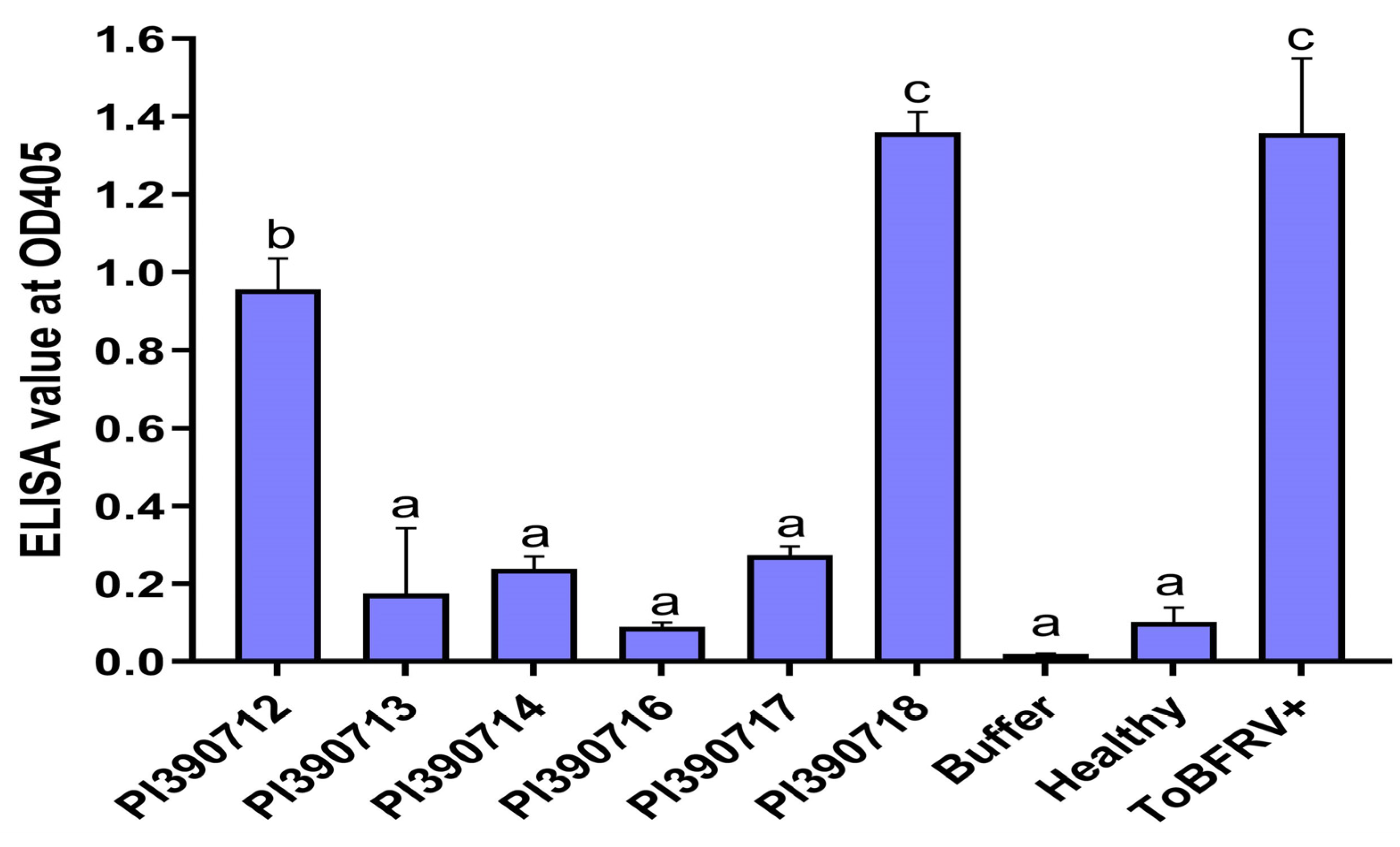
2.2. Rescreening of Selected Lines to Verify Their Resistant Properties to ToBRFV
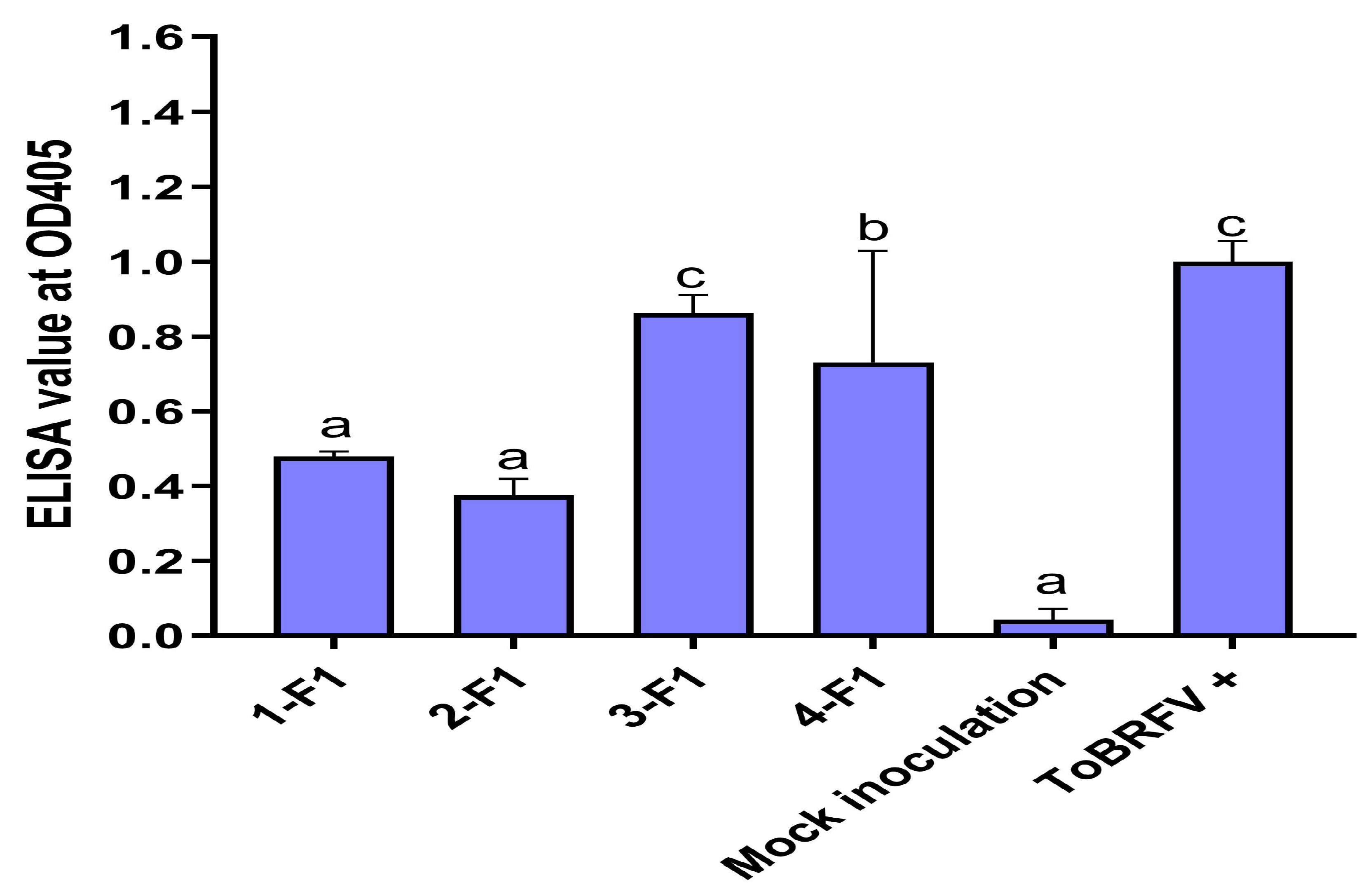
2.3. Testing F1 Progenies for Their Resistance to ToBRFV
3. Discussion
4. Materials and Methods
4.1. Plant Germplasm Materials
4.2. Virus Culture and Mechanical Inoculation
4.3. Virus Detection through a Serological Test Using Enzyme-Linked Immunosorbent Assay
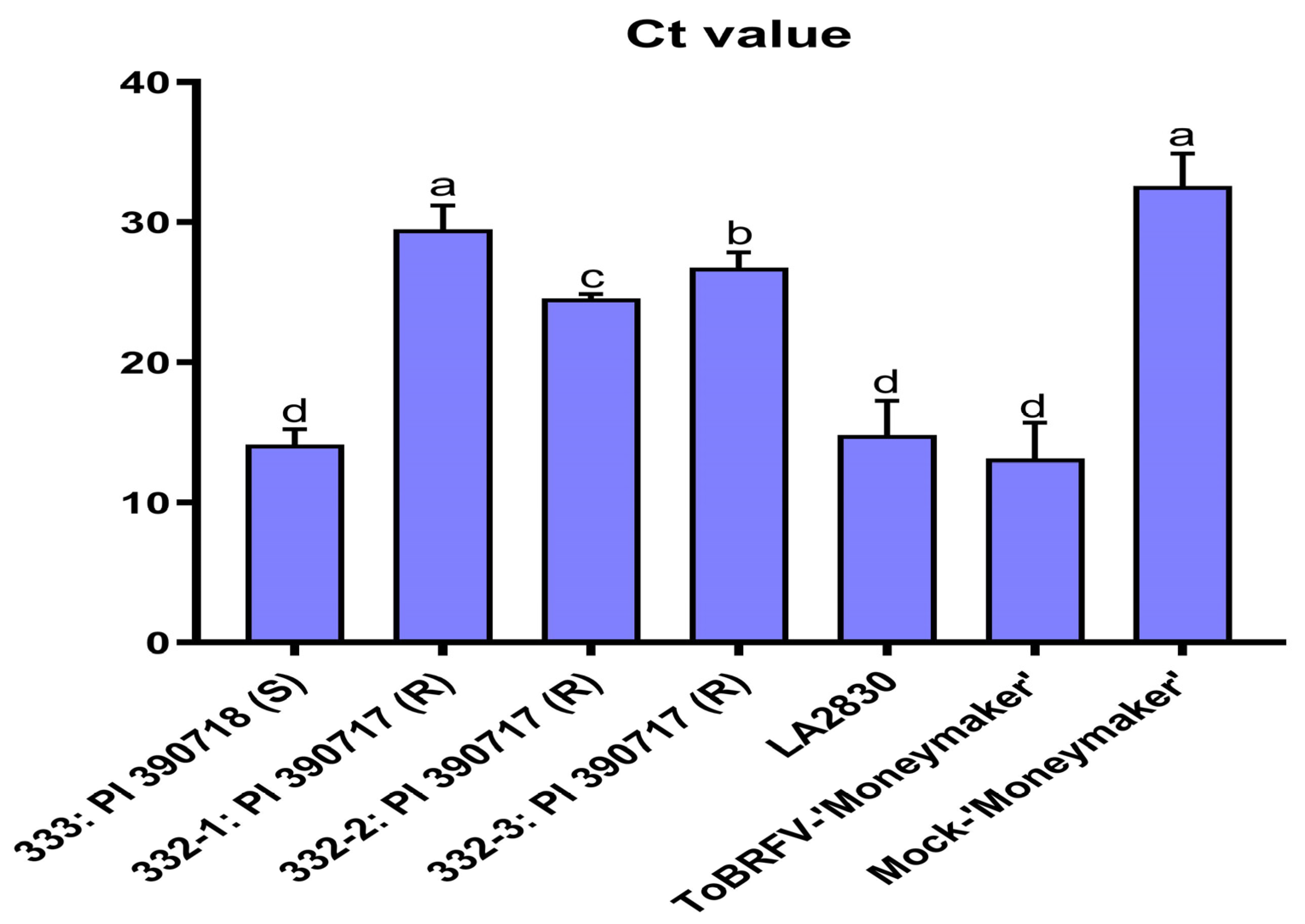
4.4. Virus Detection Using Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
4.5. Disease Scoring and Data Analysis
4.6. Advancing Selected Resistant Lines through Self-Pollination or Cross-Pollination to Generate F1 Plants for Evaluation of The Inheritability of Resistance to ToBRFV
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salem, N.M.; Jewehan, A.; Aranda, M.A.; Fox, A. Tomato brown rugose fruit virus pandemic. Annu. Rev. Phytopathol. 2023, 61, 1. [Google Scholar] [CrossRef]
- Zhang, S.; Griffiths, J.S.; Marchand, G.; Bernards, M.A.; Wang, A. Tomato brown rugose fruit virus: An emerging and rapidly spreading plant RNA virus that threatens tomato production worldwide. Mol. Plant Pathol. 2022, 23, 1262–1277. [Google Scholar] [CrossRef]
- Salem, N.M.; Mansour, A.N.; Abdeen, A.O.; Araj, S.; Khrfan, W.I. First report of tomato chlorosis virus infecting tomato crops in Jordan. Plant Dis. 2015, 99, 1286. [Google Scholar] [CrossRef]
- Luria, N.; Smith, E.; Reingold, V.; Bekelman, I.; Lapidot, M.; Levin, I.; Elad, N.; Tam, Y.; Sela, N.; Abu-Ras, A.; et al. A new israeli tobamovirus isolate infects tomato plants harboring Tm-22 resistance genes. PLoS ONE 2017, 12, e0170429. [Google Scholar] [CrossRef] [PubMed]
- Abou Kubaa, R.; Choueiri, E.; Heinoun, K.; Cillo, F.; Saponari, M. First report of tomato brown rugose fruit virus infecting sweet pepper in Syria and Lebanon. J. Plant Path. 2022, 104, 425. Available online: https://link.springer.com/article/10.1007/s42161-021-00987-y (accessed on 2 January 2024). [CrossRef]
- Alkowni, R.; Alabdallah, O.; Fadda, Z. Molecular identification of tomato brown rugose fruit virus in tomato in Palestine. J. Plant Pathol. 2019, 101, 719–723. [Google Scholar] [CrossRef]
- Ghorbani, A.; Rostami, M.; Seifi, S.; Izadpanah, K. First report of tomato brown rugose fruit virus in greenhouse tomato in Iran. New Dis. Rep. 2021, 44, e12040. [Google Scholar] [CrossRef]
- Hasan, Z.M.; Salem, N.M.; Ismail, I.D.; Akel, E.H.; Ahmad, A.Y. First report of Tomato brown rugose fruit virus on greenhouse tomato in Syria. Plant Dis. 2022, 106, 772. [Google Scholar] [CrossRef] [PubMed]
- Sabra, A.; Saleh, M.A.A.; Alshahwan, I.M.; Amer, M.A. First report of Tomato brown rugose fruit virus infecting tomato crop in Saudi Arabia. Plant Dis. 2021, 106, 1310. [Google Scholar] [CrossRef]
- Yan, Z.-Y.; Ma, H.-Y.; Han, S.-L.; Geng, C.; Tian, Y.-P.; Li, X.-D. First report of tomato brown rugose fruit virus infecting tomato in China. Plant Dis. 2019, 103, 2973. [Google Scholar] [CrossRef]
- Amer, M.A.; Mahmoud, S.Y. First report of tomato brown rugose fruit virus on tomato in Egypt. New Dis. Rep. 2020, 41, 24. [Google Scholar] [CrossRef]
- Alfaro-Fernández, A.; Castillo, P.; Sanahuja, E.; Rodríguez-Salido, M.C.; Font, M.I. First report of tomato brown rugose fruit virus in tomato in Spain. Plant Dis. 2021, 105, 515. [Google Scholar] [CrossRef]
- Beris, D.; Malandraki, I.; Kektsidou, O.; Theologidis, I.; Vassilakos, N.; Varveri, C. First report of tomato brown rugose fruit virus infecting tomato in Greece. Plant Dis. 2020, 104, 2035. [Google Scholar] [CrossRef]
- Fidan, H.; Sarikaya, P.; Calis, O. First report of tomato brown rugose fruit virus on tomato in Turkey. New Dis. Rep. 2019, 39, 18. [Google Scholar] [CrossRef]
- Hamborg, Z.; Blystad, D.-R. The first report of Tomato brown rugose fruit virus in tomato in Norway. Plant Dis. 2022, 106, 2004. [Google Scholar] [CrossRef] [PubMed]
- Mahillon, M.; Kellenberger, I.; Dubuis, N.; Brodard, J.; Bunter, M.; Weibel, J.; Sandrini, F.; Schumpp, O. First report of Tomato brown rugose fruit virus in tomato in Switzerland. New Dis. Rep. 2022, 45, e12065. Available online: https://bsppjournals.onlinelibrary.wiley.com/doi/full/10.1002/ndr2.12065 (accessed on 2 January 2024). [CrossRef]
- Menzel, W.; Knierim, D.; Winter, S.; Hamacher, J.; Heupel, M. First report of tomato brown rugose fruit virus infecting tomato in Germany. New Dis. Rep. 2019, 39, 1. [Google Scholar] [CrossRef]
- Orfanidou, C.G.; Cara, M.; Merkuri, J.; Papadimitriou, K.; Katis, N.I.; Maliogka, V.I. First report of tomato brown rugose fruit virus in tomato in Albania. J Plant Path. 2022, 104, 855. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Davino, S. First report of tomato brown rugose fruit virus on tomato crops in Italy. Plant Dis. 2019, 103, 1443. [Google Scholar] [CrossRef]
- Skelton, A.; Buxton-Kirk, A.; Ward, R.; Harju, V.; Frew, L.; Fowkes, A.; Long, M.; Negus, A.; Forde, S.; Adams, I.P.; et al. First report of Tomato brown rugose fruit virus in tomato in the United Kingdom. New Dis. Rep. 2019, 40, 12. [Google Scholar] [CrossRef]
- Skelton, A.; Gentit, P.; Porcher, L.; Visage, M.; Fowkes, A.; Adams, I.P.; Harju, V.; Webster, G.; Pufal, H.; McGreig, S.; et al. First report of Tomato brown rugose fruit virus in tomato in France. New Dis. Rep. 2022, 45, e12061. Available online: https://bsppjournals.onlinelibrary.wiley.com/doi/10.1002/ndr2.12061 (accessed on 2 January 2024). [CrossRef]
- van de Vossenberg, B.T.L.H.; Visser, M.; Bruinsma, M.; Koenraadt, H.M.S.; Westenberg, M.; Botermans, M. Real-time tracking of Tomato brown rugose fruit virus (ToBRFV) outbreaks in the Netherlands using Nextstrain. PLoS ONE 2020, 15, e0234671. [Google Scholar] [CrossRef] [PubMed]
- Cambrón-Crisantos, J.M.; Rodríguez-Mendoza, J.; Valencia-Luna, J.B.; Alcasio-Rangel, S.; García-Ávila, C.J.; López-Buenfil, J.A.; Ochoa-Martínez, D.L. First report of tomato brown rugose fruit virus (ToBRFV) in Michoacan, Mexico. Rev. Mex. Fitopatol. 2018, 37, 185–192. [Google Scholar] [CrossRef]
- Camacho-Beltrán, E.; Pérez-Villarreal, A.; Leyva-López, N.E.; Rodríguez-Negrete, E.A.; Ceniceros-Ojeda, E.A.; Méndez-Lozano, J. Occurrence of tomato brown rugose fruit virus infecting tomato crops in Mexico. Plant Dis. 2019, 103, 1440. [Google Scholar] [CrossRef]
- Ling, K.-S.; Tian, T.; Gurung, S.; Salati, R.; Gilliard, A. First report of tomato brown rugose fruit virus infecting greenhouse tomato in the United States. Plant Dis. 2019, 103, 1439. [Google Scholar] [CrossRef]
- Sarkes, A.; Fu, H.; Feindel, D.; Harding, M.; Feng, J. Development and evaluation of a loop-mediated isothermal amplification (LAMP) assay for the detection of Tomato brown rugose fruit virus (ToBRFV). PLoS ONE 2020, 15, e0230403. [Google Scholar] [CrossRef]
- Dey, K.K.; Velez-Climent, M.; Soria, P.; Batuman, O.; Mavrodieva, V.; Wei, G.; Zhou, J.; Adkins, A.; McVay, J. Frist report of tomato brown rugose fruit virus infecting tomato in Florida, USA. New Dis. Rep. 2021, 44, e12028. [Google Scholar] [CrossRef]
- Obregón, V.G.; Ibañez, J.M.; Lattar, T.E.; Juszczak, S.; Groth-Helms, D. First report of tomato brown rugose fruit virus in tomato in Argentina. New Dis. Rep. 2023, 48, e12203. [Google Scholar] [CrossRef]
- EPPO. Tomato Brown Rugose Fruit Virus (ToBRFV) Datasheet. EPPO Global Database. 2022. Available online: https://gd.eppo.int/taxon/TOBRFV/datasheet (accessed on 22 September 2023).
- Fraser, R.S.S.; Loughlin, S.A.R. Resistance to tobacco mosaic virus in tomato: Effects of the Tm-1 gene on virus multiplication. J. General. Virol. 1980, 48, 87–96. [Google Scholar] [CrossRef]
- Holmes, F.O. Inheritance of resistance to infection by tobacco-mosaic virus in tomato. Phytopathology 1954, 44, 640–642. [Google Scholar]
- Pelham, J. Strain-genotype interaction of tobacco mosaic virus in tomato. Ann. Appl. Biol. 1972, 71, 219–228. [Google Scholar] [CrossRef]
- de Ronde, D.; Butterbach, P.; Kormelink, R. Dominant resistance against plant viruses. Front. Plant Sci. 2014, 5, 307. [Google Scholar] [CrossRef]
- Pfitzner, A.J.P. Resistance to tobacco mosaic virus and tomato mosaic virus in tomato. In Natural Resistance Mechanisms of Plants to Viruses; Loebenstein, G., Carr, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 399–413. [Google Scholar]
- Maayan, Y.; Pandaranayaka, E.P.J.; Srivastava, D.A.; Lapidot, M.; Levin, I.; Dombrovsky, A.; Harel, A. Using genomic analysis to identify tomato Tm-2 resistance-breaking mutations and their underlying evolutionary path in a new and emerging tobamovirus. Arch. Virol. 2018, 163, 1863–1875. [Google Scholar] [CrossRef]
- Chanda, B.; Gilliard, A.; Jaiswal, N.; Ling, K.-S. Comparative analysis of host range, ability to infect tomato cultivars with Tm-22 gene, and real-time reverse transcription PCR detection of tomato brown rugose fruit virus. Plant Dis. 2021, 105, 3643–3652. [Google Scholar] [CrossRef]
- Alexander, L.J. Transfer of a dominant type of resistance to the four known Ohio pathogenic strains of tobacco mosaic virus (TMV) from Lycopersicon peruvianum to L. esculentum. Phytopathology 1963, 53, 869. [Google Scholar]
- Paudel, D.B.; Sanfacon, H. Exploring the diversity of mechanisms associated with plant tolerance to virus infection. Front. Plant Sci. 2018, 9, 1575. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.-C.; Yeam, I.; Jahn, M.M. Genetics of plant virus resistance. Annu. Rev. Phytopathol. 2005, 43, 581–621. [Google Scholar] [CrossRef] [PubMed]
- Ponz, F.; Bruening, G. Mechanisms of resistance to plant viruses. Ann. Rev. Phytopathol. 1986, 24, 355–381. [Google Scholar] [CrossRef]
- Calil, I.P.; Fontes, E.P.B. Plant immunity against viruses: Antiviral immune receptors in focus. Ann. Bot. 2017, 119, 711–723. [Google Scholar] [CrossRef]
- Zinger, A.; Lapidot, M.; Harel, A.; Doron-Faigenboim, A.; Gelbart, D.; Levin, I. Identification and mapping of tomato genome loci controlling tolerance and resistance to tomato brown rugose fruit virus. Plants 2021, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Jewehan, A.; Salem, N.; Tóth, Z.; Salamon, P.; Szabó, Z. Screening of Solanum (sections Lycopersicon and Juglandifolia) germplasm for reactions to the tomato brown rugose fruit virus (ToBRFV). J. Plant Dis. Protect. 2022, 129, 117–123. [Google Scholar] [CrossRef]
- Jewehan, A.; Salem, N.; Tóth, Z.; Salamon, P.; Szabó, Z. Evaluation of responses to tomato brown rugose fruit virus (ToBRFV) and selection of resistant lines in Solanum habrochaites and Solanum peruvianum germplasm. J. Gen. Plant Pathol. 2022, 88, 187–196. [Google Scholar] [CrossRef]
- Kabas, A.; Fidan, H.; Kucukaydin, H.; Atan, H.N.; Kabas, A.; Fidan, H.; Kucukaydin, H.; Atan, H.N. Screening of wild tomato species and interspecific hybrids for resistance/tolerance to Tomato brown rugose fruit virus (ToBRFV). Chil. J. Agric. Res. 2022, 82, 189–196. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Smith, E. Seed Transmission of Tobamoviruses: Aspects of Global Disease Distribution. Interchopen 2017. [Google Scholar] [CrossRef]
- Chanda, B.; Shamimuzzaman, M.; Gilliard, A.; Ling, K.-S. Effectiveness of disinfectants against the spread of tobamoviruses: Tomato brown rugose fruit virus and Cucumber green mottle mosaic virus. Virol. J. 2021, 18, 7. [Google Scholar] [CrossRef] [PubMed]
- Davino, S.; Caruso, A.G.; Bertacca, S.; Barone, S.; Panno, S. Tomato brown rugose fruit virus: Seed transmission rate and efficacy of different seed disinfection treatments. Plants 2020, 9, 1615. [Google Scholar] [CrossRef] [PubMed]
- Dombrovsky, A.; Mor, N.; Gantz, S.; Lachman, O.; Smith, E. Disinfection Efficacy of Tobamovirus-Contaminated Soil in Greenhouse-Grown Crops. Horticulturae 2022, 8, 563. [Google Scholar] [CrossRef]
- Ehlers, J.; Zarghani, S.N.; Kroschewski, B.; Büttner, C.; Bandte, M. Cleaning of Tomato brown rugose fruit virus (ToBRFV) from contaminated clothing of greenhouse employees. Horticulturae 2022, 8, 751. [Google Scholar] [CrossRef]
- Ehlers, J.; Zarghani, S.N.; Kroschewski, B.; Büttner, C.; Bandte, M. Decontamination of tomato brown rugose fruit virus-contaminated shoe soles under practical conditions. Horticulturae 2022, 8, 1210. [Google Scholar] [CrossRef]
- Ling, K.-S.; Gilliard, A.C.; Zia, B. Disinfectants useful to manage the emerging tomato brown rugose fruit virus in greenhouse tomato production. Horticulturae 2022, 8, 1193. [Google Scholar] [CrossRef]
- Rodríguez-Díaz, C.I.; Zamora-Macorra, E.J.; Ochoa-Martínez, D.L.; González-Garza, R. Disinfectants effectiveness in Tomato brown rugose fruit virus (ToBRFV) transmission in tobacco plants. Rev. Mex. Fitopatol. (Mex. J. Phytopathol.) 2022, 40, 240–253. [Google Scholar] [CrossRef]
- Samarah, N.; Sulaiman, A.; Salem, N.M.; Turina, M. Disinfection treatments eliminated tomato brown rugose fruit virus in tomato seeds. Eur. J. Plant Pathol. 2021, 159, 153–162. [Google Scholar] [CrossRef]
- Kole, C. Wild Crop Relatives: Genomic and Breeding Resources: Vegetables; Springer: Berlin, Germany, 2011. [Google Scholar]
- Chanda, B.; Rivera, Y.; Nunziata, S.O.; Galvez, M.E.; Gilliard, A.C.; Ling, K.-S. Complete genome sequence of a tomato brown rugose fruit virus isolated in the United States. Microbiol. Resour. Announc. 2020, 9, e00630-20. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Total Number of Accessions Screened | Number of Accessions in Resistance/Tolerance |
|---|---|---|
| Solanum arcanum | 10 | 0 |
| Solanum chilense | 17 | 0 |
| Solanum corneliomulleri | 17 | 1 |
| Solanum habrochaites | 50 | 4 |
| S. huaylasense | 3 | 0 |
| Solanum lycopersicum | 10 | 0 |
| Solanum lycopersicum var. cerasiforme | 1 | 0 |
| Solanum neorickii | 1 | 0 |
| Solanum pennellii | 1 | 0 |
| Solanum peruvianum | 73 | 3 |
| Solanum pimpinellifolium | 140 | 31 |
| Solanum subsect. lycopersicon hybrid | 153 | 5 |
| Total | 476 | 44 |
| Plant ID | Taxonomy | Disease Severity Index (%) |
|---|---|---|
| PI 129144 | Solanum corneliomulleri | 0 |
| PI 126445 | Solanum habrochaites | 17.6 |
| PI 209978 | Solanum habrochaites | 0 |
| PI 247087 | Solanum habrochaites | 0 |
| LA 2107 | Solanum habrochaites | 0 |
| PI 306811 | Solanum peruvianum | 16 |
| PI 390667 | Solanum peruvianum | 0 |
| PI 390671 | Solanum peruvianum | 0 |
| PI 127805 | Solanum pimpinellifolium | 14.2 |
| PI 143524 | Solanum pimpinellifolium | 14.2 |
| PI 143527 | Solanum pimpinellifolium | 0 |
| PI 211838 | Solanum pimpinellifolium | 0 |
| PI 230327 | Solanum pimpinellifolium | 0 |
| PI 344102 | Solanum pimpinellifolium | 0 |
| PI 344103 | Solanum pimpinellifolium | 0 |
| PI 346340 | Solanum pimpinellifolium | 0 |
| PI 390692 | Solanum pimpinellifolium | 0 |
| PI 390693 | Solanum pimpinellifolium | 0 |
| PI 390694 | Solanum pimpinellifolium | 0 |
| PI 390695 | Solanum pimpinellifolium | 0 |
| PI 390698 | Solanum pimpinellifolium | 0 |
| PI 390699 | Solanum pimpinellifolium | 18 |
| PI 390700 | Solanum pimpinellifolium | 0 |
| PI 390702 | Solanum pimpinellifolium | 11.4 |
| PI 390710 | Solanum pimpinellifolium | 0 |
| PI 390712 | Solanum pimpinellifolium | 0 |
| PI 390713 | Solanum pimpinellifolium | 0 |
| PI 390714 | Solanum pimpinellifolium | 0 |
| PI 390716 | Solanum pimpinellifolium | 0 |
| PI 390717 | Solanum pimpinellifolium | 0 |
| PI 390720 | Solanum pimpinellifolium | 8.4 |
| PI 390722 | Solanum pimpinellifolium | 0 |
| PI 390723 | Solanum pimpinellifolium | 16 |
| PI 390724 | Solanum pimpinellifolium | 0 |
| PI 390725 | Solanum pimpinellifolium | 3.4 |
| PI 390726 | Solanum pimpinellifolium | 0 |
| PI 390727 | Solanum pimpinellifolium | 0 |
| PI 390750 | Solanum pimpinellifolium | 0 |
| PI 432362 | Solanum pimpinellifolium | 8.8 |
| PI 127799 | Solanum subsect. lycopersicon hybrid | 17.8 |
| PI 129143 | Solanum subsect. lycopersicon hybrid | 0 |
| PI 143522 | Solanum subsect. lycopersicon hybrid | 18 |
| PI 233930 | Solanum subsect. lycopersicon hybrid | 0 |
| PI 237640 | Solanum subsect. lycopersicon hybrid | 3.4 |
| Zinger et al., 2021 [42] | Kabas et al., 2022 [45] | Jewehan et al., 2022a [43] | Jewehan et al., 2022b [44] | This Study | |
|---|---|---|---|---|---|
| Total lines | 160 | 44 | 636 | 173 | 476 |
| Tolerant lines | S. pimpinellifolium (9); S. Lycopersicum (8) | S. pimpinnelifolium (1); S. penellii (1); and S. chilense (2) | S. pimpinelifolium (26); S. chilense (1); S. lycopersicum var. cerasiforme (4) | S. corneliomulleri (1); S. habrochaites (4); S. peruvianum (3); S. pimpinellifolium (27); and S. subsect. lycopersicon hybrid (5) | |
| Resistant lines | S. lycopersicum (1) | S. ochrantum (5) | S. habrochaites (9); S. peruvianum (1) | S. pimpinellifolium (4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaiswal, N.; Chanda, B.; Gilliard, A.; Shi, A.; Ling, K.-S. Evaluation of Tomato Germplasm against Tomato Brown Rugose Fruit Virus and Identification of Resistance in Solanum pimpinellifolium. Plants 2024, 13, 581. https://doi.org/10.3390/plants13050581
Jaiswal N, Chanda B, Gilliard A, Shi A, Ling K-S. Evaluation of Tomato Germplasm against Tomato Brown Rugose Fruit Virus and Identification of Resistance in Solanum pimpinellifolium. Plants. 2024; 13(5):581. https://doi.org/10.3390/plants13050581
Chicago/Turabian StyleJaiswal, Namrata, Bidisha Chanda, Andrea Gilliard, Ainong Shi, and Kai-Shu Ling. 2024. "Evaluation of Tomato Germplasm against Tomato Brown Rugose Fruit Virus and Identification of Resistance in Solanum pimpinellifolium" Plants 13, no. 5: 581. https://doi.org/10.3390/plants13050581