
Use of Pseudomonas protegens to Control Root Rot Disease Caused by Boeremia exigua var. exigua in Industrial Chicory (Cichorium intybus var. sativum Bisch.)

 ,
,
Abstract
:1. Introduction
2. Results
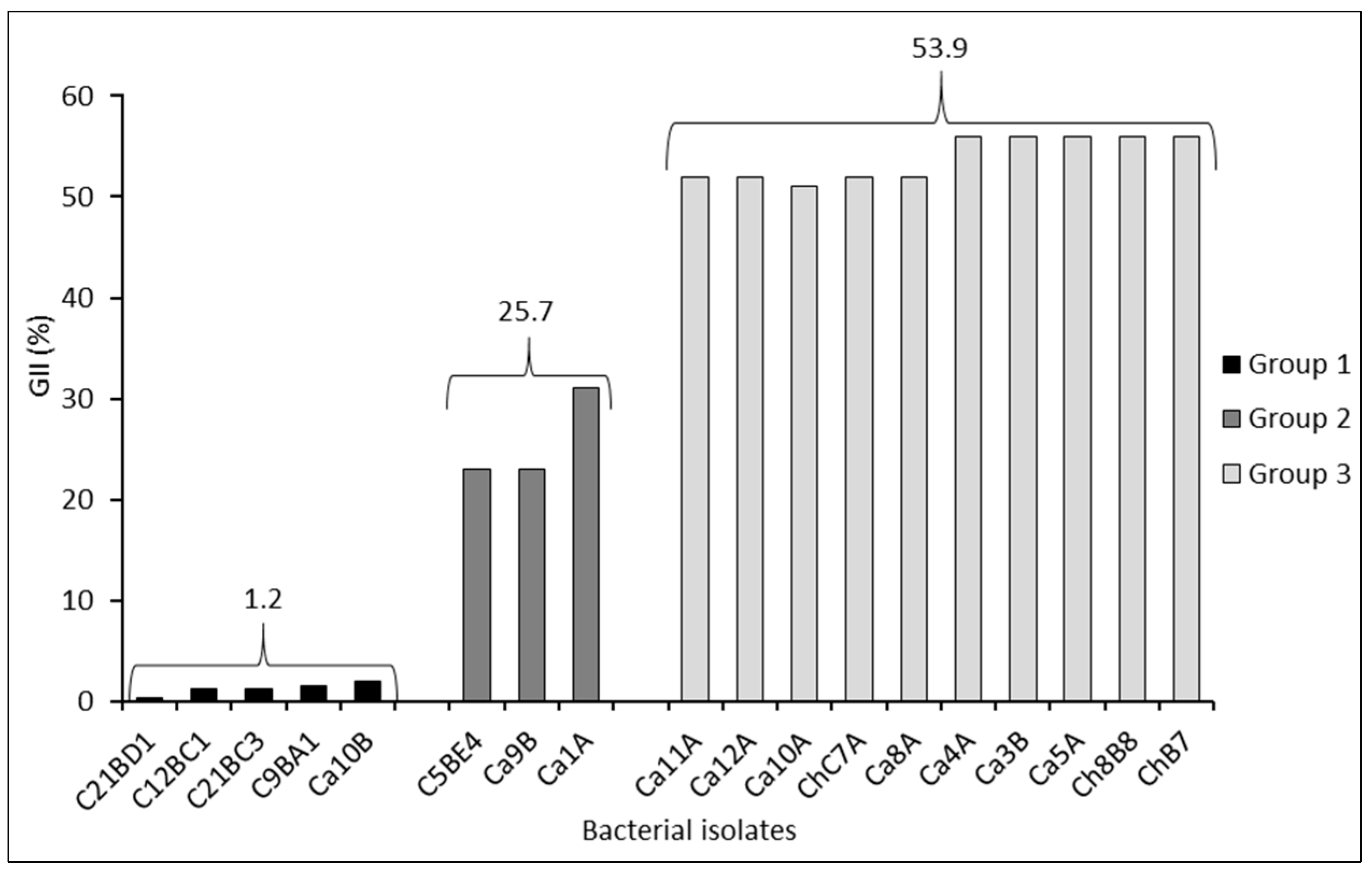
2.1. In Vitro Preselection of the Antagonistic Effect of Pseudomonas protegens Strains on the Pathogenic Fungus B. exigua var. exigua under In Vitro Conditions
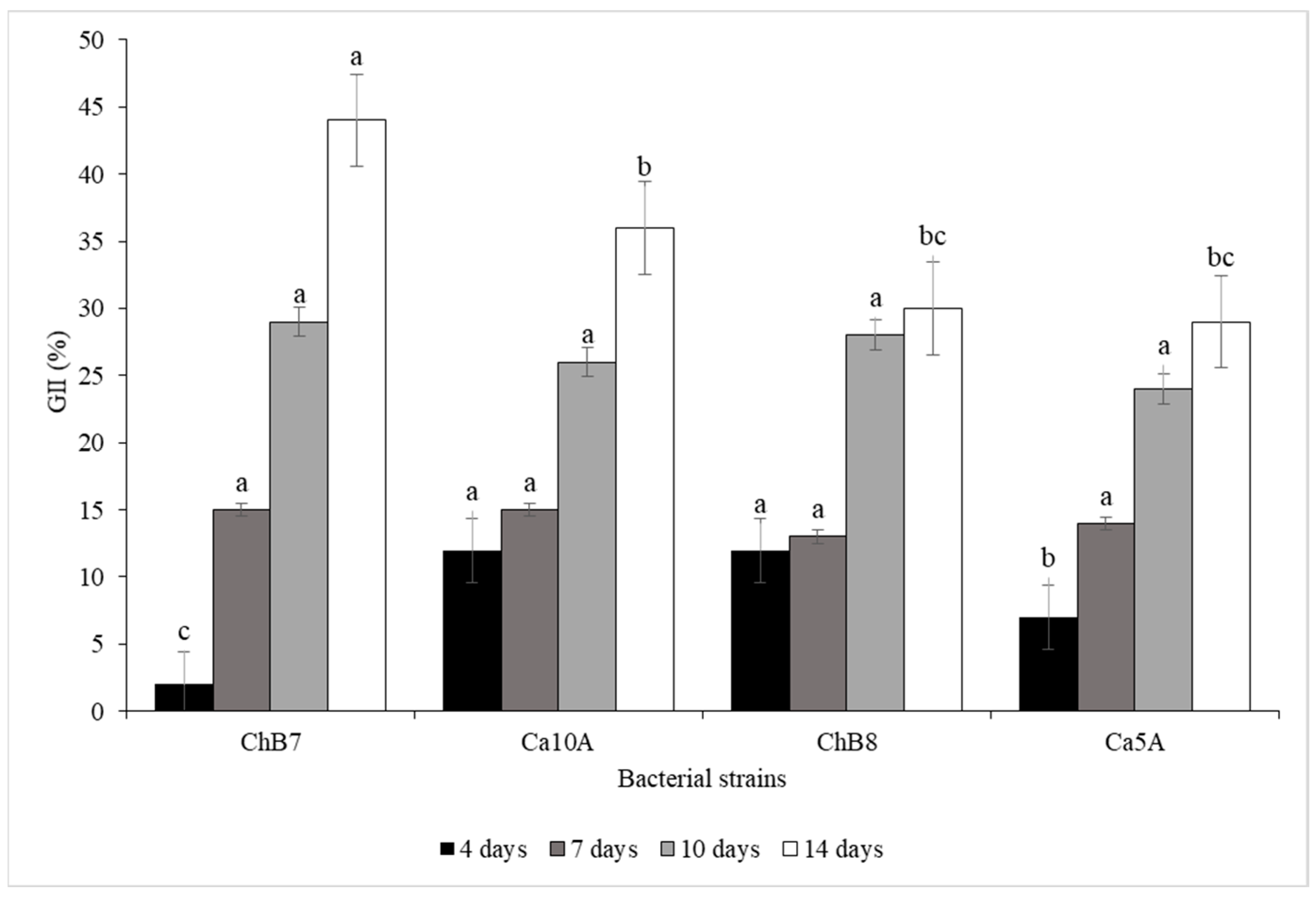
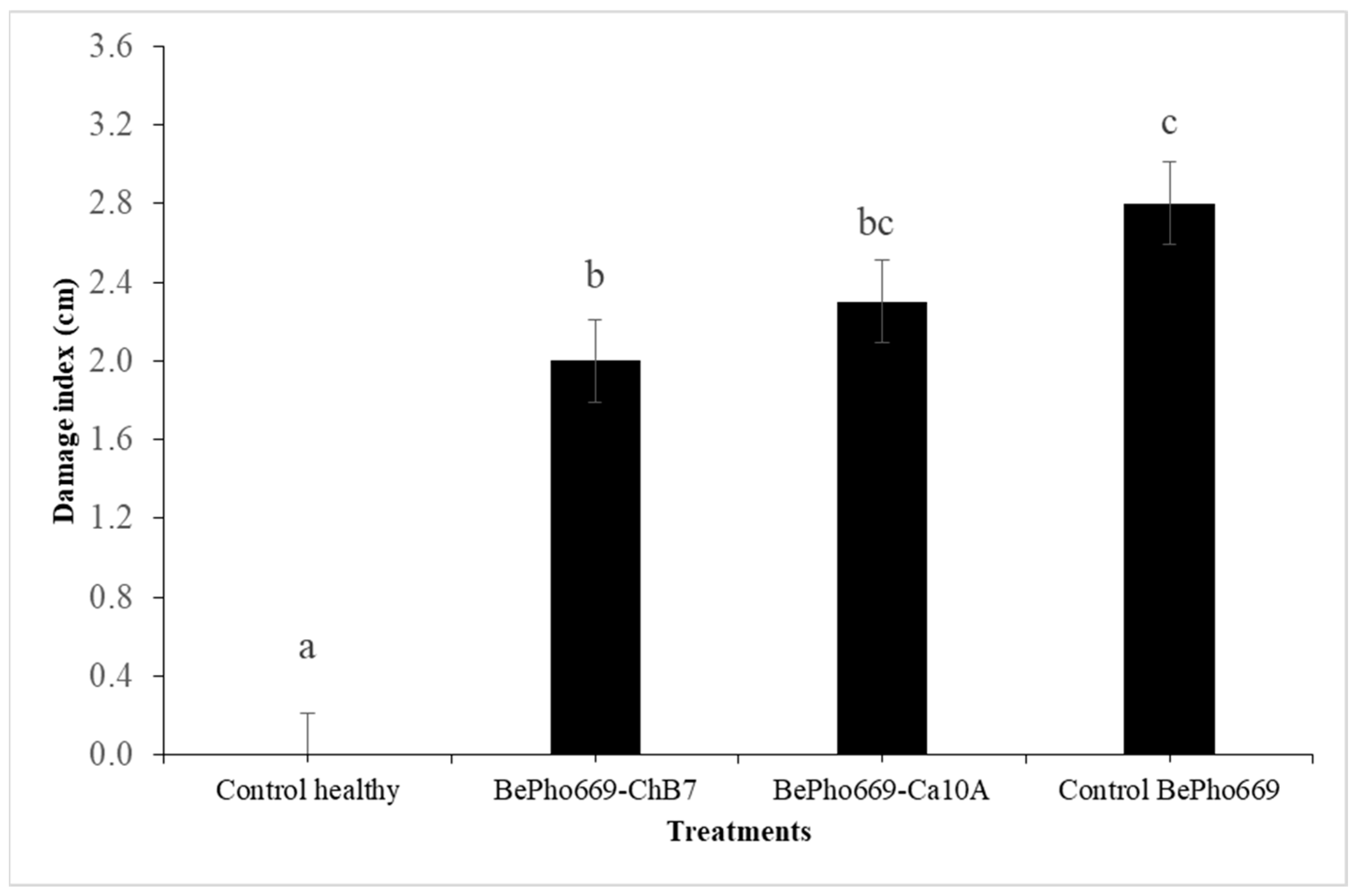
2.2. Evaluation of the Control Effectiveness of Pseudomonas protegens on Boeremia exigua var. exigua in Roots of Industrial Chicory under Controlled Conditions
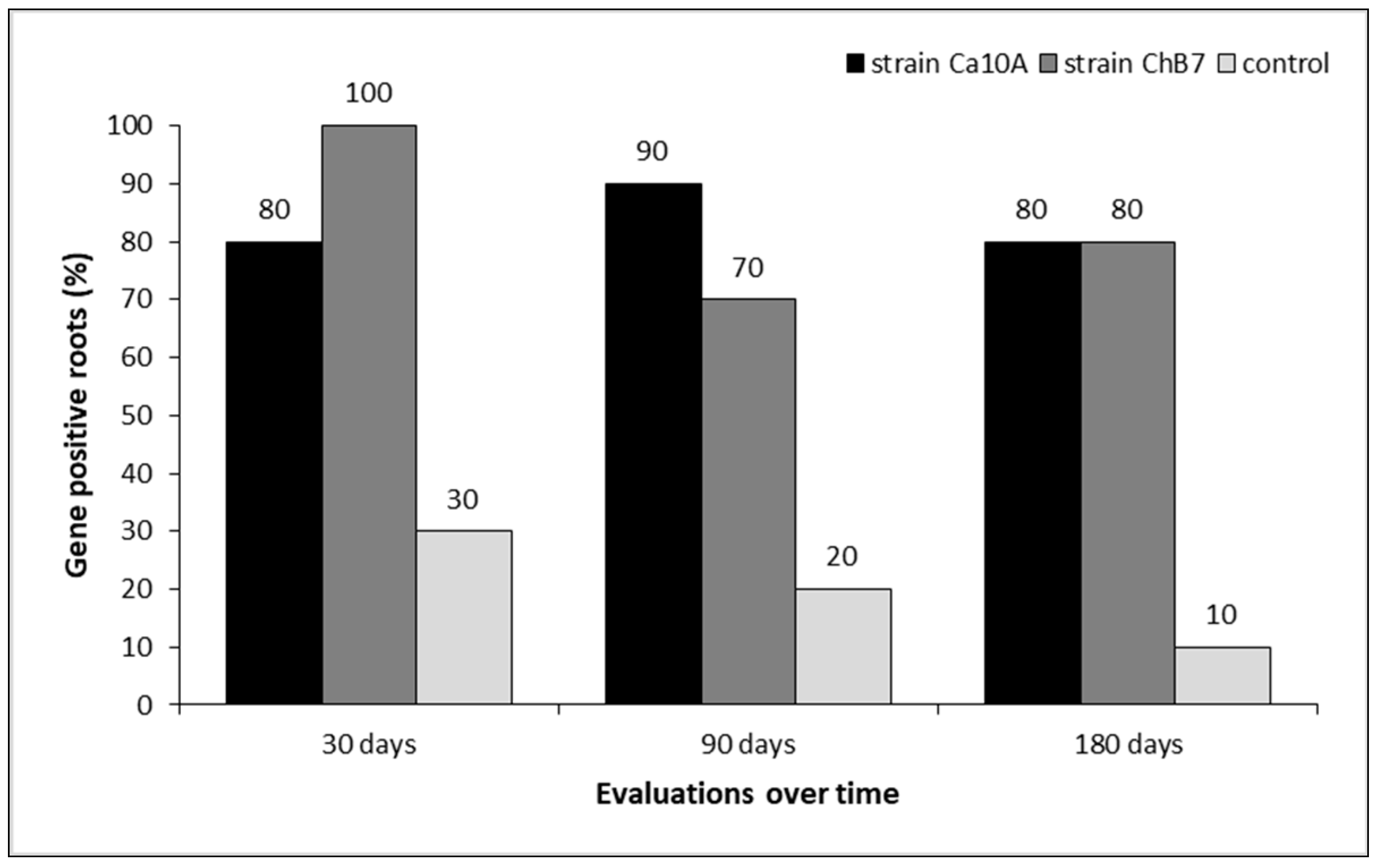
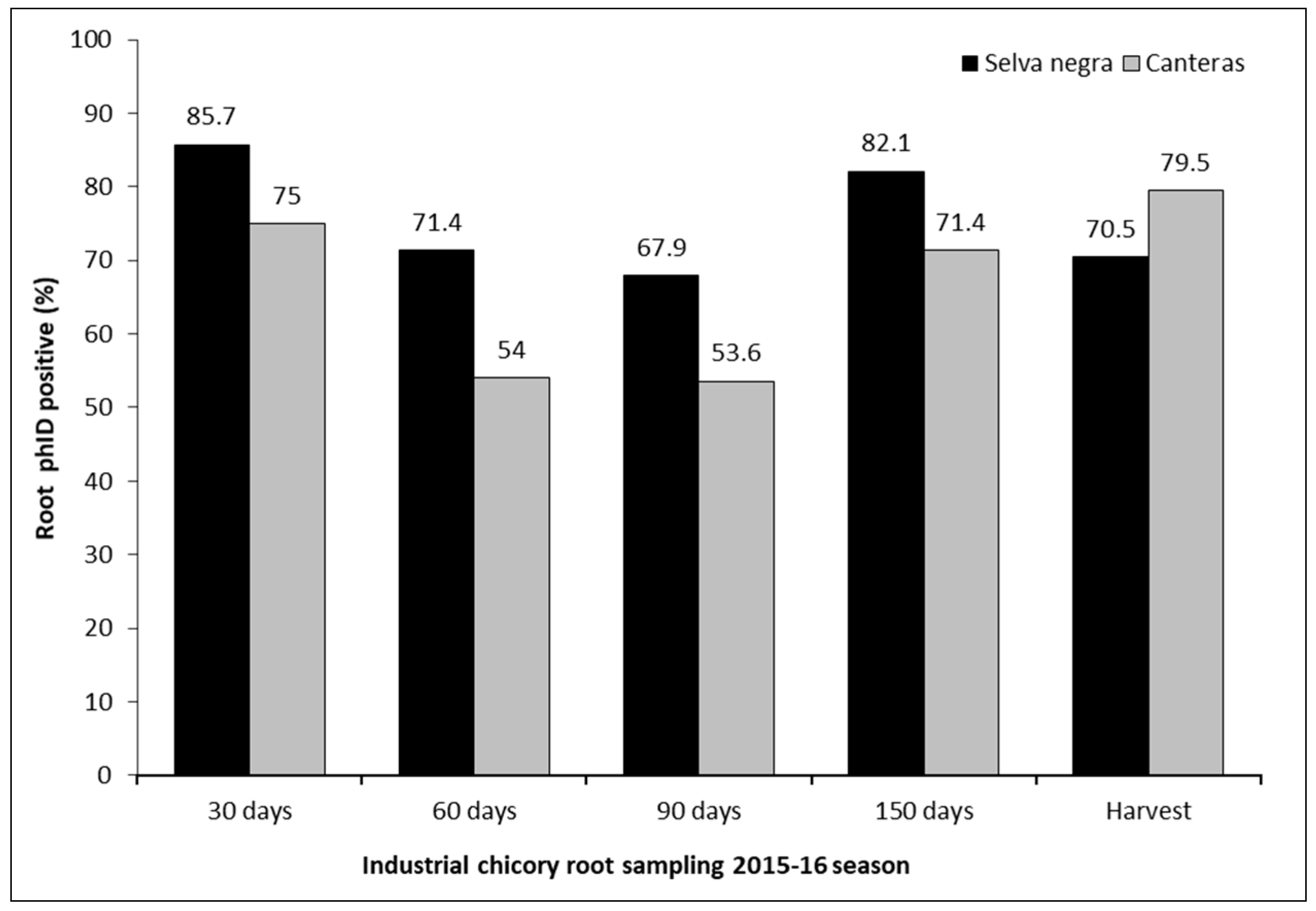
2.3. Evaluation of the Root Colonization Capacity and Control Effectiveness of Antagonistic Bacteria (P. protegens) on Root Rot Caused by the Pathogen B. exigua var. exigua in Roots of Industrial Chicory under Controlled and Field Conditions
3. Discussion
4. Materials and Methods
4.1. Isolates of B. exigua var. exigua
4.2. In Vitro Preselection of the Antagonistic Effect of Bacterial Isolates on the Pathogenic Fungus B. exigua var. exigua
4.3. Evaluation of the Control Effectiveness of P. protegens on B. exigua var. exigua in Roots of Industrial Chicory Grown under Controlled Conditions
4.4. Evaluation of the Colonization Capacity of P. protegens in Industrial Chicory Roots Grown in Pots under Controlled Conditions
4.5. Evaluation of the Root Colonization Capacity and Control Effectiveness of Antagonistic Bacteria on Root Rot Caused by the Pathogen B. exigua var. exigua in Industrial Chicory Grown under Field Conditions
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vargas, M.; Loyola, C.; Zapata, N.; Rivera, V.; Secor, G.; Bolton, M.; France, A. First Report of Root Rot of Chicory Caused by Phytophthora cryptogea in Chile. Plant Dis. 2012, 96, 591. [Google Scholar] [CrossRef]
- Muys, C.; Thienpont, C.-N.; Dauchot, N.; Maudoux, O.; Draye, X.; Cutsem, P.V. Integration of AFLPs, SSRs and SNPs markers into a new genetic map of industrial chicory (Cichorium intybus L. var. sativum). Plant Breed. 2014, 133, 130–137. [Google Scholar] [CrossRef]
- Velasco, V.; Soto, V.H.; Williams, P.; Campos, J.; Astudillo, R.; Rodríguez, H. Meat quality parameters of broiler chickens fed diets containing chicory (Cichorium intybus) vinasse. Chil. J. Agric. Anim. Sci. 2018, 34, 26–32. [Google Scholar] [CrossRef]
- Grinbergs, D.; France, A.; Varrelmann, M. First report of Boeremia exigua var. exigua (syn. Phoma exigua var. exigua) causing black root rot on industrial chicory (Cichorium intybus var. sativum) in Chile. Plant Dis. 2016, 100, 2328. [Google Scholar] [CrossRef]
- Boerema, G.H. Phoma Identification Manual: Differentiation of Specific and Infra-Specific Taxa in Culture; CABI: Wallingford, UK, 2004. [Google Scholar]
- Agrios, G.N. Plant Pathology; Academic Press: San Diego, CA, USA, 1997; Volume 4. [Google Scholar]
- Irinyi, L.; Kövics, G.J.; Sándor, E. Taxonomical re-evaluation of Phoma-like soybean pathogenic fungi. Mycol. Res. 2009, 113, 249–260. [Google Scholar] [CrossRef]
- Koike, S.T.; Subbarao, K.V.; Verkley, G.J.M.; Fogle, D.; O’Neill, T.M. Phoma basal rot of Romaine lettuce in California caused by Phoma exigua: Occurrence, characterization, and control. Plant Dis. 2006, 90, 1268–1275. [Google Scholar] [CrossRef]
- Deb, D.; Khan, A.; Deyet, N. Phoma disease. Epidemiology and control. Plant Pathol. 2020, 69, 1203–1217. [Google Scholar] [CrossRef]
- Orafti. Manual Técnico Achicoria Industrial Temporada 2018–2019. Orafti (Chile). Available online: http://www.orafti.cl/download/manualtecnicocultivo2014-2015.pdf (accessed on 17 October 2023).
- Mavrodi, O.V.; Walter, N.; Elateek, S.; Taylor, C.G.; Okubara, P.A. Suppression of Rhizoctonia and Pythium root rot of wheat by new strains of Pseudomonas. Biol. Control 2012, 62, 93–102. [Google Scholar] [CrossRef]
- Hewedy, O.A.; Abdel Lateif, K.S.; Seleiman, M.F.; Shami, A.; Albarakaty, F.M.; M El-Meihy, R. Phylogenetic diversity of Trichoderma strains and their antagonistic potential against soil-borne pathogens under stress conditions. Biology 2020, 23, 189. [Google Scholar] [CrossRef]
- Ahmed, H.F.A.; Seleiman, M.F.; Al-Saif, A.M.; Alshiekheid, M.A.; Battaglia, M.L.; Taha, R.S. Biological control of celery powdery mildew disease caused by Erysiphe heraclei DC in vitro and in vivo conditions. Plants 2021, 29, 2342. [Google Scholar] [CrossRef]
- Elbrense, H.; Elmasry, A.M.A.; Seleiman, M.F.; AL-Harbi, M.S.; Abd El-Raheem, A.M. 2021. Can symbiotic bacteria (Xenorhabdus and Photorhabdus) be more efficient than their entomopathogenic nematodes against Pieris rapae and Pentodon algerinus larvae? Biology 2021, 10, 999. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Moreno-Ramírez, L.; Valdez-Salas, B.; Seleiman, M.F.; El-Hendawy, S.; Aldhuwaib, K.J.; Alotaibi, M.; González-Mendoza, D. New Bacillus subtilis strains isolated from Prosopis glandulosa rhizosphere for suppressing Fusarium spp. and enhancing growth of Gossypium hirsutum L. Biology 2023, 12, 73. [Google Scholar] [CrossRef]
- Blackburn, D.; Shapiro-Ilan, D.I.; Adams, B.J. Biological control and nutrition: Food for thought. Biol. Control 2016, 97, 131–138. [Google Scholar] [CrossRef]
- Ramette, A.; Frapolli, M.; Saux, M.F.-L.; Gruffaz, C.; Meyer, J.-M.; Défago, G.; Sutra, L.; Moënne-Loccoz, Y. Pseudomonas protegens sp. nov., widespread plant-protecting bacteria producing the biocontrol compounds 2,4-diacetylphloroglucinol and pyoluteorin. Syst. Appl. Microbiol. 2011, 34, 180–188. [Google Scholar] [CrossRef]
- Weller, D.M.; Landa, B.; Mavrodi, O.; Schroeder, K.; De La Fuente, L.; Bankhead, S.B.; Molar, R.A.; Bonsall, R.; Mavrodi, D.; Thomashow, L. Role of 2, 4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in the defense of plant roots. Plant Biol. 2006, 9, 4–20. [Google Scholar] [CrossRef]
- De La Fuente, L.; Mavrodi, D.V.; Landa, B.B.; Thomashow, L.S.; Weller, D.M. phlD-based genetic diversity and detection of genotypes of 2, 4-diacetylphloroglucinol-producing Pseudomonas fluorescens. FEMS Microbiol. Ecol. 2006, 56, 64–78. [Google Scholar] [CrossRef]
- Brazelton, J.N.; Pfeufer, E.E.; Sweat, T.A.; Gardener, B.B.M.; Coenen, C. 2,4-diacetylphloroglucinol alters plant root development. Molecular Plant Microbe Interact. 2008, 21, 1349–1358. [Google Scholar] [CrossRef]
- Nowak-Thompson, B.; Chaney, N.; Wing, J.S.; Gould, S.J.; Loper, J.E. Characterization of the pyoluteorin biosynthetic gene cluster of Pseudomonas fluorescens Pf-5. J. Bacteriol. 1999, 181, 2166–2174. [Google Scholar] [CrossRef]
- Brodhagen, M.; Henkels, M.D.; Loper, J.E. Positive autoregulation and signaling properties of pyoluteorin, an antibiotic produced by the biological control organism Pseudomonas fluorescens Pf-5. Appl. Environ. Microbiol. 2004, 70, 1758–1766. [Google Scholar] [CrossRef]
- Kidarsa, T.A.; Goebel, N.C.; Zabriskie, T.M.; Loper, J.E. Phloroglucinol mediates cross-talk between the pyoluteorin and 2,4-diacetylphloroglucinol biosynthetic pathways in Pseudomonas fluorescens Pf-5. Mol. Microbiol. 2011, 81, 395–414. [Google Scholar] [CrossRef]
- Troppens, D.M.; Dmitriev, R.I.; Papkovsky, D.B.; O’Gara, F.; Morrissey, J.P. Genome-wide investigation of cellular targets and mode of action of the antifungal bacterial metabolite 2,4-diacetylphloroglucinol in Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Durán, P.; Jorquera, M.; Viscardi, S.; Carrion, V.J.; Mora, M.d.l.L.; Pozo, M.J. Screening and characterization of potentially suppressive soils against Gaeumannomyces graminis under extensive wheat cropping by Chilean indigenous communities. Front. Microbiol. 2017, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Doussoulin, H.A.; Arismendi, N.L.; Moya-Elizondo, E.A. Pseudomonas spp. producing antimicrobial compounds regulate fungal communities inhabiting wheat crown in Southern Chile. Agronomy 2022, 12, 710. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.; Arismendi, N.; Castro, M.P.; Doussoulin, H. Distribution and prevalence of crown rot pathogens affecting wheat crops in southern Chile. Chil. J. Agr. Res. 2015, 75, 78–84. [Google Scholar] [CrossRef]
- Henkels, M.D.; Kidarsa, T.A.; Shaffer, B.T.; Goebel, N.C.; Burlinson, P.; Mavrodi, D.V.; Bentley, M.A.; Rangel, L.I.; Davis, E.W.; Thomashow, L.S.; et al. Pseudomonas protegens Pf-5 causes discoloration and pitting of mushroom caps due to the production of antifungal metabolites. Molecular Plant Microbe Interac. 2014, 27, 733–746. [Google Scholar] [CrossRef]
- Yan, Q.; Philmus, B.; Chang, J.H.; Loper, J.E. Novel mechanism of metabolic co-regulation coordinates the biosynthesis of secondary metabolites in Pseudomonas protegens. ELife 2017, 6, e22835. [Google Scholar] [CrossRef]
- Jousset, A.; Bonkowski, M. The model predator Acanthamoeba castellanii induces the production of 2,4-DAPG by the biocontrol strain Pseudomonas fluorescens Q2-87. Soil Biol. Biochem. 2010, 42, 1647–1649. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Weller, D.M. Exploiting genotypic diversity of 2,4-diacetylphloroglucinol-producing Pseudomonas spp.: Characterization of superior root-colonizing P. fluorescens strain Q8r1-96. Appl. Environ. Microbiol. 2001, 67, 2545–2554. [Google Scholar] [CrossRef]
- Grobelak, A.; Napora, A.; Kacprzak, M. Using plant growth-promoting rhizobacteria (PGPR) to improve plant growth. Ecol. Eng. 2015, 84, 22–28. [Google Scholar] [CrossRef]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R. Isolation and characterization of PGPR and their effect on growth, yield and nutrient content in wheat (Triticum aestivum L.). Biocatal. Agric. Biotechnol. 2014, 3, 121–128. [Google Scholar] [CrossRef]
- Subhashini, D.V.; Padmaja, K. Effect of bioinoculants on seedling vigour in tobacco (Nicotiana tabacum) nurseries. Indian J. Agric. Sci. 2011, 80, 124. [Google Scholar]
- Loyola Contreras, C.A. Phytophthora cryptogea y Phoma exigua var. Exigua: Patógenos causantes de la Pudrición Radicular en Achicoria Industrial (Cichorium Intybus var. Sativum Bisch.). Agricultural Engineer Thesis, Universidad de Concepción, Concepción, Chile, 2013; 30p. [Google Scholar]
- Stolpe, N. Description of the Principal Soil Types of Region VIII of Chile; Publicaciones del Departamento de Suelos y Recursos Naturales; Facultad de Agronomía, Universidad de Concepción: Concepción, Chile, 2006; Volume 84. [Google Scholar]
- Dandamudi Vallabhaneni, S. Biocontrol of Rhizoctonia solani in tobacco (Nicotiana tabacum) seed beds using Pseudomonas fluorescens. Agric. Res. 2016, 5, 137–144. [Google Scholar] [CrossRef]
- Millas, P.; France, A. Manejo de Enfermedades en Agricultura Agroecológica. In Agroecología: Fundamentos, Técnicas de Producción y Experiencia en la Región de los Ríos; Colección Libros INIA; Céspedes, L., Vargas, S., Eds.; Instituto de Investigaciones Agropecuarias: Osorno, Chile, 2021; Available online: https://hdl.handle.net/20.500.14001/68311 (accessed on 23 October 2023).
- Imperiali, N.; Dennert, F.; Schneider, J.; Laessle, T.; Velatta, C.; Fesselet, M.; Wyler, M.; Mascher, F.; Mavrodi, O.; Mavrodi, D.; et al. Relationships between root pathogen resistance, abundance and expression of Pseudomonas antimicrobial genes, and soil properties in representative Swiss agricultural soils. Front. Plant Sci. 2017, 8, 427. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Mercado-Blanco, J.; Bakker, P.A.H.M. Interactions between plants and beneficial Pseudomonas spp.: Exploiting bacterial traits for crop protection. Antonie Van Leeuwenhoek 2007, 92, 367–389. [Google Scholar] [CrossRef]
- Vera Palma, C.A.; Madariaga Burrows, R.P.; González, M.G.; Sepúlveda, B.R.; Moya-Elizondo, E.A. Integration between Pseudomonas protegens strains and fluquinconazole for the control of take-all in wheat. Crop Protect. 2019, 121, 163–172. [Google Scholar] [CrossRef]
- Castro Tapia, M.P.; Madariaga Burrows, R.P.; Ruiz Sepúlveda, B.; Vargas Concha, M.; Vera Palma, C.; Moya-Elizondo, E.A. Antagonistic activity of Chilean strains of Pseudomonas protegens against fungi causing crown and root rot of wheat (Triticum aestivum L.). Front. Plant Sci. 2020, 11, 951. [Google Scholar] [CrossRef]
- Höfte, M.; Altier, N. Fluorescent pseudomonads as biocontrol agents for sustainable agricultural systems. Res. Microbiol. 2010, 161, 464–471. [Google Scholar] [CrossRef]
- Spanoghe, P.; Ryckaert, B.; Van Gheluwe, C.; Van Labeke, M.-C. Fate of vinclozolin, thiabendazole and dimethomorph during storage, handling and forcing of chicory. Pest Manag. Sci. 2010, 66, 126–131. [Google Scholar] [CrossRef]
- Vega, Y. In Vitro Evaluation of Growth Promotion Mechanisms in Bacteria with Antimicrobial Activity on Phytopathogenic Fungi. Agricultural Engineer Thesis, Universidad de Concepción, Concepción, Chile, 2017; 30p. [Google Scholar]
- Govindasamy, V.; Senthilkumar, M.; Mageshwaran, V.; Annapurna, K. Detection and characterization of ACC deaminase in plant growth promoting rhizobacteria. J. Plant Biochem. Biotechnol. 2009, 18, 71–76. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.; Cattan, N.; Arismendi, N.; Doussoulin, H. Determination of 2,4-diacetylphloroglucinol (2,4-DAPG) and phenazine-producing Pseudomonas spp. in wheat crops in southern Chile. Phytopathology 2013, 103 (Suppl. 2), 100. [Google Scholar]
- McSpadden Gardener, B.B.; Mavrodi, D.V.; Thomashow, L.S.; Weller, D.M. A rapid Polymerase Chain Reaction-Based assay characterizing rhizosphere populations of 2,4-diacetylphloroglucinol-producing bacteria. Phytopathology 2001, 91, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.J.; Pasche, J.S.; Gudmestad, N.C. Biological significance of mefenoxam resistance in Phytophthora erythroseptica and its implications for the management of pink rot of potato. Plant Dis. 2006, 90, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, P. Descripciones de Suelos VIII Región; Universidad de Concepción, Facultad de Agronomía, Departamento de Suelo: Concepción, Chile, 1998. [Google Scholar]
- Vettraino, A.M.; Belisario, A.; Maccaroni, M.; Anselmi, N.; Vannini, A. First report of Phytophthora cryptogea in walnut stands in Italy. Plant Dis. 2002, 86, 328. [Google Scholar] [CrossRef]
- Vettraino, A.; Flamini, L.; Pizzichini, L.; Prodi, A.; Nipoti, P.; Vannini, A.; Lagnese, R. First report of root and collar rot by Phytophthora cryptogea on sweet cherry in Italy. Plant Dis. 2008, 92, 177. [Google Scholar] [CrossRef]
- Schroeder, K.L.; Paulitz, T.C. Effect of inoculum density and soil tillage on the development and severity of Rhizoctonia root rot. Phytopathology 2008, 98, 304–314. [Google Scholar] [CrossRef]
- SAS Institute Inc. The SAS System for Windows Version 8.0.; SAS Institute Inc.: Cary, NC, USA, 1998. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Fresh Leaf Weight (ton ha−1) | Healthy Root Weight (ton ha−1) | Diseased Root Weight (ton ha−1) | |||
|---|---|---|---|---|---|---|
| Non-inoculated control | 16.5 ± 0.6 | b | 98.6 ± 3.7 | a | 11.7 ± 1.1 | ab |
| Control inoculated with Pho669 | 13.5 ± 1.0 | a | 93.4 ± 2.9 | a | 18.5 ± 2.9 | c |
| Seed inoculated with ChB7 | 14.8 ± 0.6 | ab | 97.8 ± 1.7 | a | 10.1 ± 1.2 | a |
| Seed inoculated with ChB7 + Pho669 | 14.0 ± 0.5 | a | 100.5 ± 0.9 | ab | 11.1± 0.2 | ab |
| Seed inoculated with Ca10A | 16.2 ± 0.3 | b | 105.2 ± 2.1 | ab | 10,3 ± 0.9 | a |
| Seed inoculated with Ca10A + Pho669 | 13.0 ± 0.6 | a | 102.8 ± 2.8 | ab | 7.6 ± 0.3 | a |
| Seed inoculated with Ca10A + ChB7 | 14.4 ± 0.4 | a | 111.9 ± 3.0 | b | 9.7 ± 1.7 | a |
| Seed inoculated with Ca10A + ChB7 + Pho669 | 15.6 ± 0.6 | ab | 95.9 ± 0.7 | a | 17.6 ± 1.2 | c |
| Coefficient of variation (CV) % | 10.2 | 6.9 | 33.4 | |||
| p-value | 0.041 | 0.021 | 0.005 | |||
| Treatments | Fresh Leaf Weight (ton ha−1) | Healthy Root Weight (ton ha−1) | Diseased Root Weight (ton ha−1) | |||
|---|---|---|---|---|---|---|
| Non-inoculated control | 10.3 ± 0.7 | a | 59.6 ± 1.8 | ab | 3.3 ± 1.0 | a |
| Control inoculated with Pho669 | 11.5 ± 0.8 | ab | 53.5 ± 1.7 | a | 5.5 ± 0.7 | ab |
| Seed inoculated with ChB7 | 11.5 ± 0.4 | ab | 64.8 ± 1.4 | b | 2.7 ± 0.2 | a |
| Seed inoculated with ChB7 + Pho669 | 11.9 ± 0.6 | ab | 61.5 ± 1.2 | ab | 9.4 ± 0.9 | b |
| Seed inoculated with Ca10A | 10.7 ± 0.6 | ab | 61.3 ± 1.5 | ab | 2.0 ± 0.2 | a |
| Seed inoculated with Ca10A + Pho669 | 11.5 ± 0.6 | ab | 62.6 ± 1.8 | ab | 2.0 ± 0.2 | a |
| Seed inoculated with Ca10A + ChB7 | 12.7 ± 0.5 | b | 66.2 ± 1.3 | b | 4.3 ± 0.5 | ab |
| Seed inoculated with Ca10A + ChB7+ Pho669 | 10.1 ± 0.7 | a | 60.5 ± 1.8 | ab | 4.3 ± 1.3 | a |
| Coefficient of variation (CV) % | 12.5 | 10.8 | 42.5 | |||
| p-value | 0.001 | 0.043 | 0.004 | |||
| Treatments | Root Rot Incidence (%) | Root Rot Severity (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Selva Negra | Canteras | Selva Negra | Canteras | |||||
| Non-inoculated control | 62.7 ± 1.4 | a | 16.2 ± 2.3 | a | 25.8 ± 0.3 | a | 7.7 ± 1.5 | a |
| Control inoculated with Pho669 | 81.1 ± 5.6 | b | 22.9 ± 1.9 | a | 35.6 ± 1.7 | b | 11.9 ± 1.3 | a |
| Seed inoculated with ChB7 | 56.9 ± 1.6 | a | 20.3 ± 1.3 | a | 28.0 ± 1.4 | ab | 9.6 ± 0.6 | a |
| Seed inoculated with ChB7 + Pho669 | 66.8 ± 0.2 | a | 43.1 ± 4.3 | b | 25.9 ± 0.2 | a | 23.5 ± 3.4 | b |
| Seed inoculated with Ca10A | 61.0 ± 2.8 | a | 22.0 ± 1.6 | a | 23.4 ± 1.0 | a | 9.4 ± 0.7 | a |
| Seed inoculated with Ca10A+ Pho669 | 62.7 ± 2.2 | a | 23.5 ± 3.7 | a | 23.8 ± 1.0 | a | 10.6 ± 1.7 | a |
| Seed inoculated with Ca10A + ChB7 | 58.3 ± 2.2 | a | 24.9 ± 2.2 | ab | 23.4 ± 0.8 | a | 11.7 ± 1.1 | a |
| Seed inoculated with Ca10A + ChB7 + Pho669 | 66.9 ± 1.0 | a | 29.9 ± 4.6 | ab | 27.8 ± 0.5 | ab | 13.1 ± 2.3 | ab |
| Coefficient of variation (CV) % | 12.5 | 26.3 | 8.7 | 28.9 | ||||
| p-value | 0.033 | 0.023 | 0.002 | 0.041 | ||||
| Treatment | Description |
|---|---|
| T1 | Non-inoculated control |
| T2 | Control inoculated with Pho669 |
| T3 | Seed inoculated with strain ChB7 |
| T4 | Seed inoculated with strains ChB7 and Pho669 |
| T5 | Seed inoculated with strain Ca10A |
| T6 | Seed inoculated with strains Ca10A and Pho669 |
| T7 | Seed inoculated with strains Ca10A and ChB7 |
| T8 | Seed inoculated with strains Ca10A and ChB7, and Pho669 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quezada-D’Angelo, T.; San Martín, J.; Ruiz, B.; Oyarzúa, P.; Vargas, M.; Fischer, S.; Cortés, P.; Astete, P.; Moya-Elizondo, E. Use of Pseudomonas protegens to Control Root Rot Disease Caused by Boeremia exigua var. exigua in Industrial Chicory (Cichorium intybus var. sativum Bisch.). Plants 2024, 13, 263. https://doi.org/10.3390/plants13020263
Quezada-D’Angelo T, San Martín J, Ruiz B, Oyarzúa P, Vargas M, Fischer S, Cortés P, Astete P, Moya-Elizondo E. Use of Pseudomonas protegens to Control Root Rot Disease Caused by Boeremia exigua var. exigua in Industrial Chicory (Cichorium intybus var. sativum Bisch.). Plants. 2024; 13(2):263. https://doi.org/10.3390/plants13020263
Chicago/Turabian StyleQuezada-D’Angelo, Tamara, Juan San Martín, Braulio Ruiz, Pía Oyarzúa, Marisol Vargas, Susana Fischer, Pamela Cortés, Patricio Astete, and Ernesto Moya-Elizondo. 2024. "Use of Pseudomonas protegens to Control Root Rot Disease Caused by Boeremia exigua var. exigua in Industrial Chicory (Cichorium intybus var. sativum Bisch.)" Plants 13, no. 2: 263. https://doi.org/10.3390/plants13020263