
Fighting Tomato Fungal Diseases with a Biocontrol Product Based on Amoeba Lysate


Abstract
:1. Introduction
2. Results
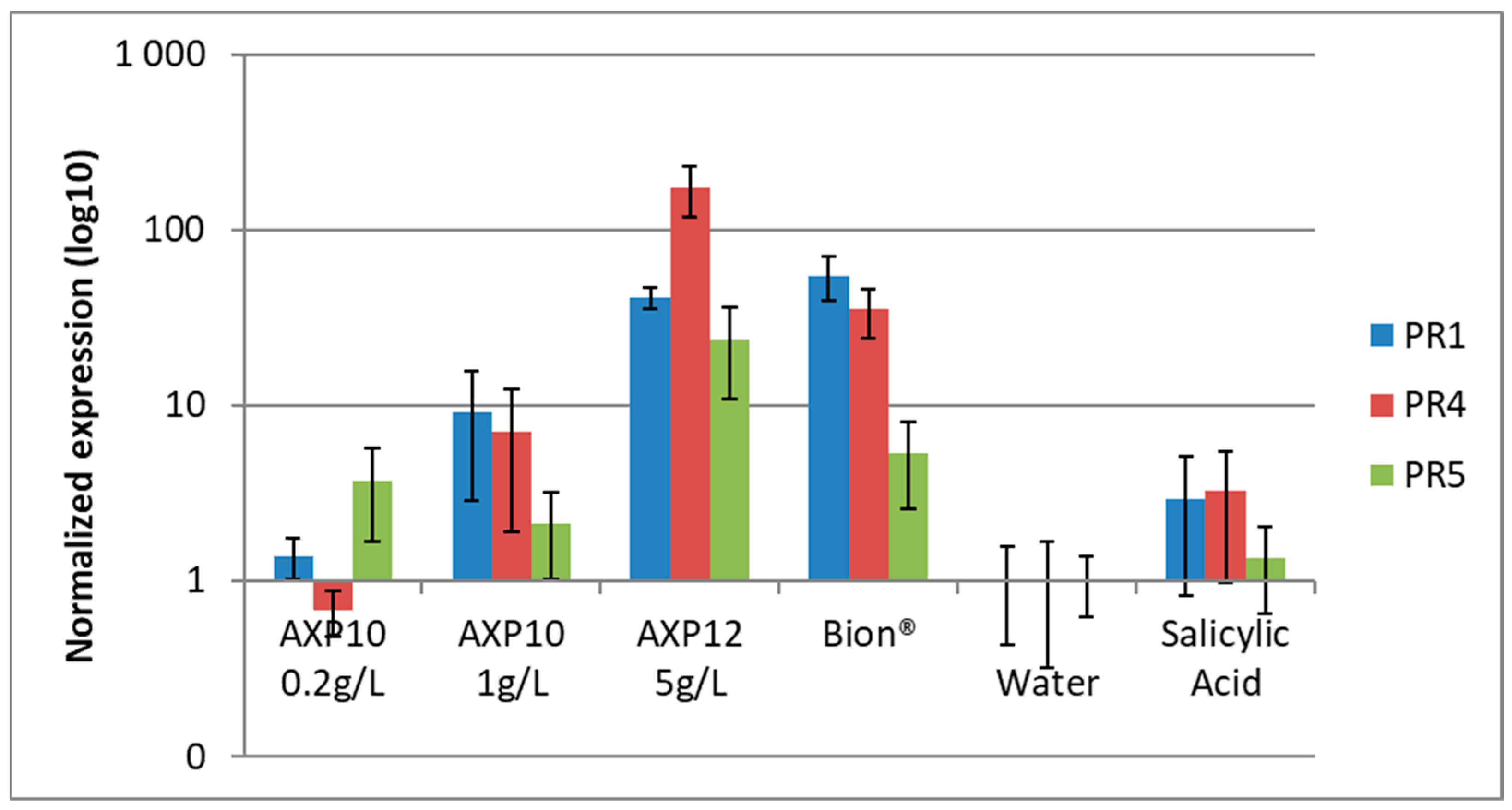
2.1. Elicitor Effect on Tomato Genes
2.2. Efficacy against Diseases
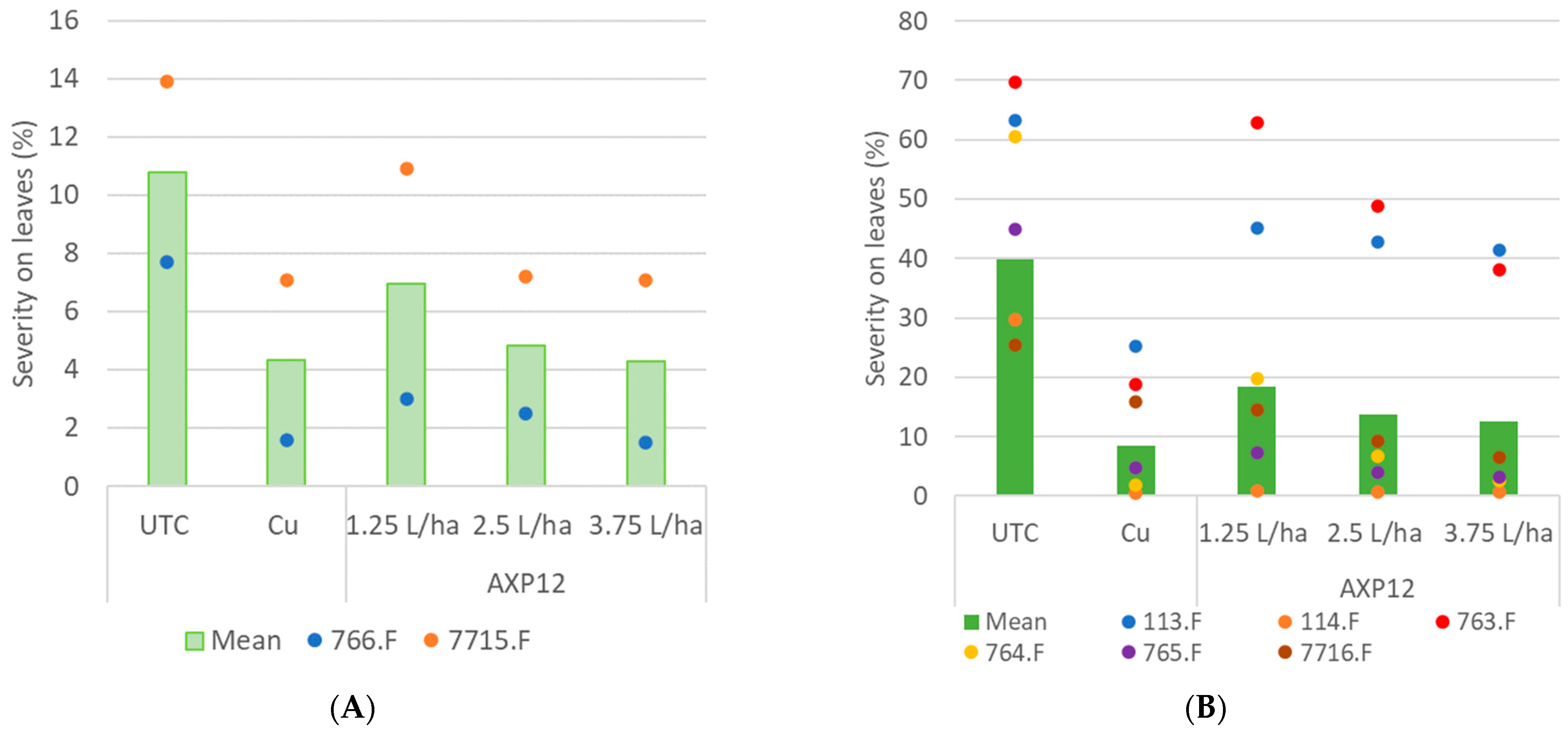
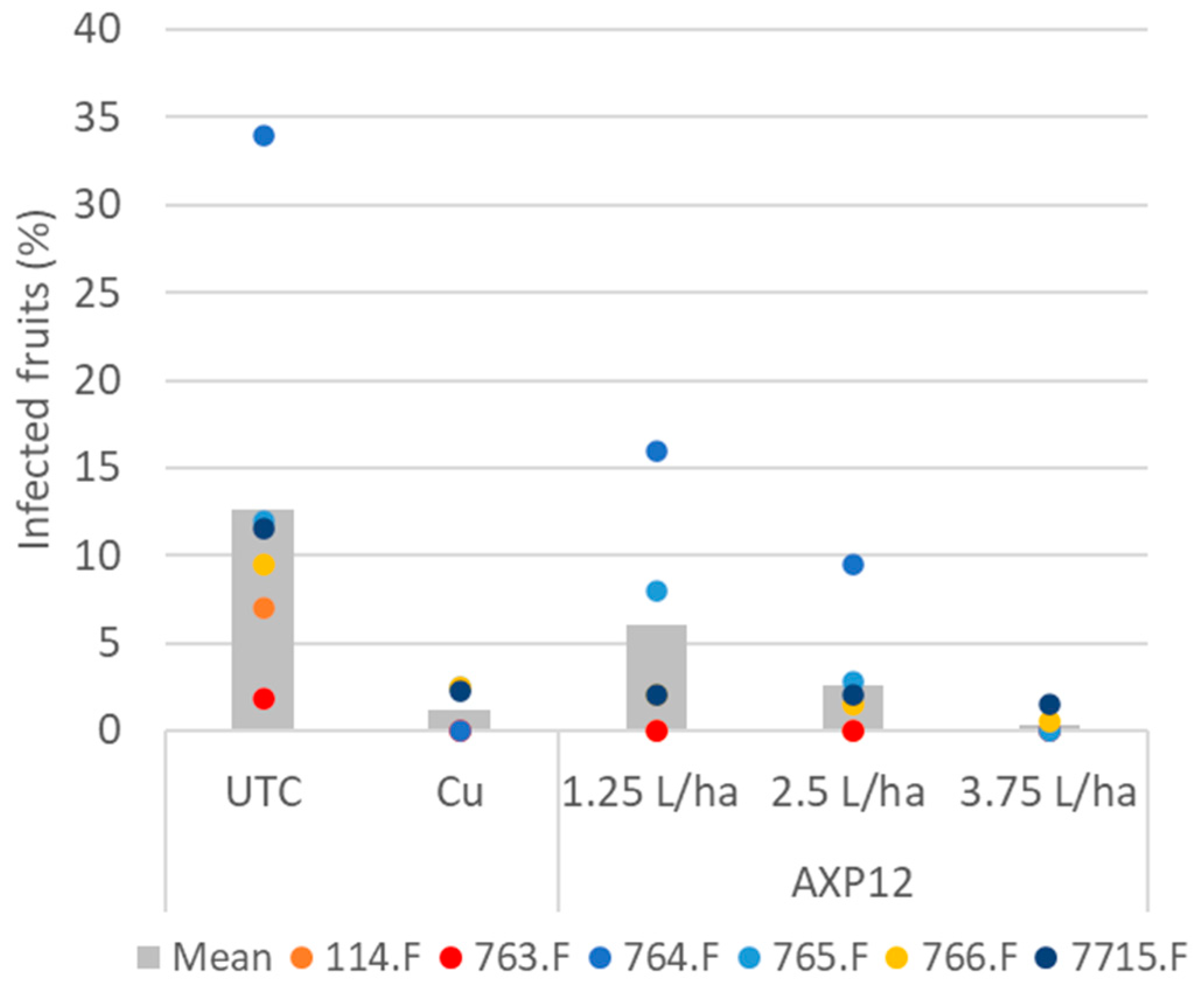
2.2.1. Tomato Late Blight
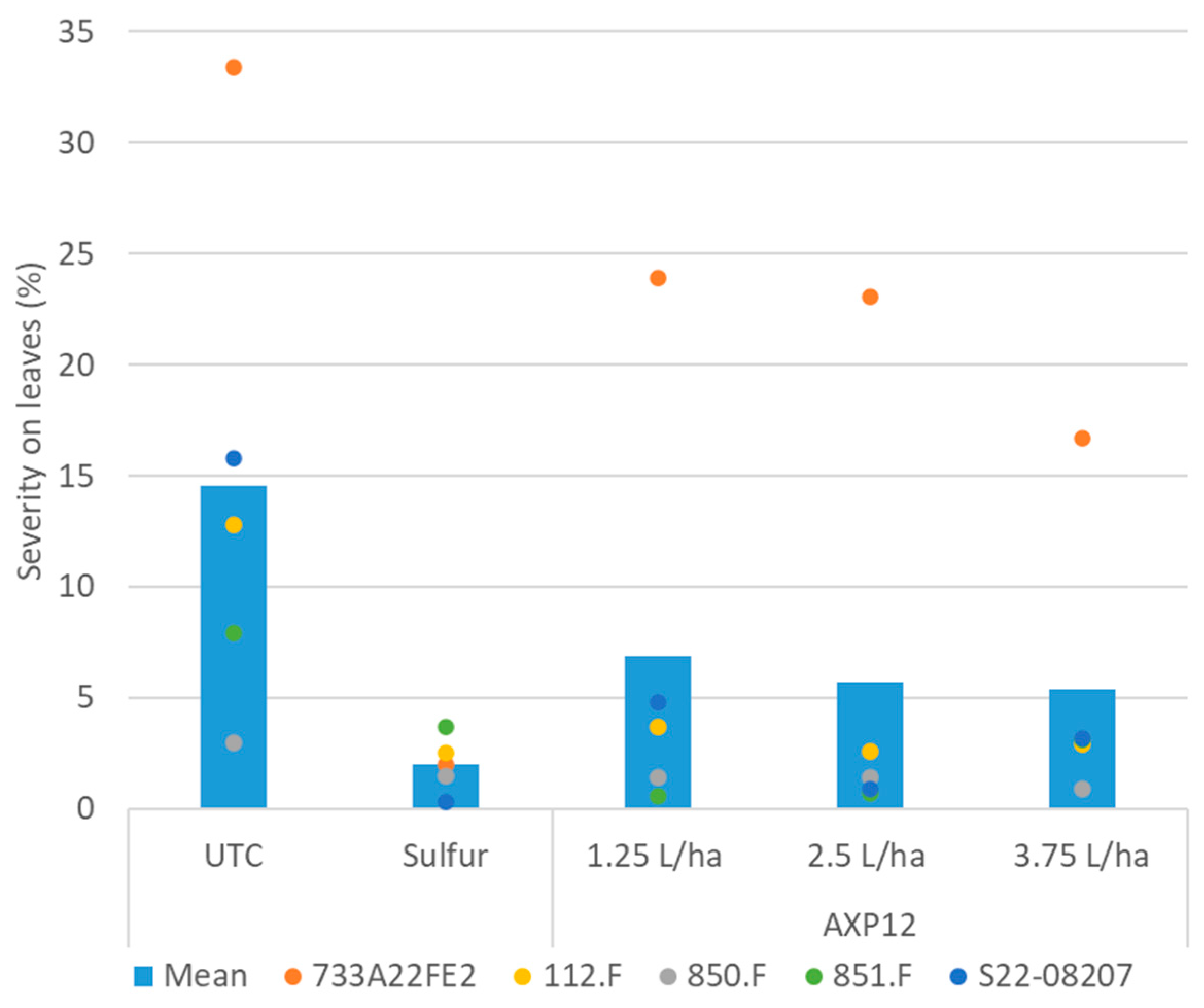
2.2.2. Powdery Mildew
3. Discussion
4. Materials and Methods
4.1. Active Substance and Formulation
4.2. Stimulation of Plant Defense PR Protein Genes
4.2.1. Plant Material and Products
4.2.2. Plant Treatment and Sampling
4.2.3. RT-qPCR Analysis
4.3. Greenhouse and Field Trials
4.3.1. Trials with Phytophthora Infestans
4.3.2. Trials with Oidium neolycopersici or Leveillula taurica
4.3.3. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Croze, A.; Carlino, A.; Quélard, B.; Saha, A.; Convert, T.; Eberst, J.-B.; Demanèche, S. Intracellular Behaviour of Legionella non-pneumophila Strains within Three Amoeba Strains, Including Willaertia magna C2c Maky. Pathogens 2021, 10, 1350. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Mameri, M.R.; Trajkovic-Bodennec, S.; Bodennec, J.; Pernin, P. Impact of Inter-Amoebic Phagocytosis on the L. pneumophila Growth. FEMS Microbiol. Lett. 2020, 367, fnaa147. [Google Scholar] [CrossRef] [PubMed]
- Demanèche, S.; Mirabel, L.; Abbe, O.; Eberst, J.-B.; Souche, J.-L. A New Active Substance Derived from Lyzed Willaertia magna C2c Maky Cells to Fight Grapevine Downy Mildew. Plants 2020, 9, 1013. [Google Scholar] [CrossRef]
- Troussieux, S.; Gilgen, A.; Souche, J.-L. A New Biocontrol Tool to Fight Potato Late Blight Based on Willaertia magna C2c Maky Lysate. Plants 2022, 11, 2756. [Google Scholar] [CrossRef] [PubMed]
- European Commission. A European Green Deal. Available online: https://ec.europa.eu/info/strategy/priorities-2019-2024/european-green-deal_en (accessed on 10 August 2020).
- Curl, C.L.; Spivak, M.; Phinney, R.; Montrose, L. Synthetic Pesticides and Health in Vulnerable Populations: Agricultural Workers. Curr. Environ. Health Rep. 2020, 7, 13–29. [Google Scholar] [CrossRef]
- Tago, D.; Andersson, H.; Treich, N. Pesticides and Health: A Review of Evidence on Health Effects, Valuation of Risks, and Benefit-Cost Analysis. Adv. Health Econ. Health Serv. Res. 2014, 24, 203–295. [Google Scholar]
- Bergougnoux, V. The History of Tomato: From Domestication to Biopharming. Biotechnol. Adv. 2014, 32, 170–189. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 13 July 2022).
- Fry, W.E.; Goodwin, S.B. Re-Emergence of Potato and Tomato Late Blight in the United States. Plant Dis. 1997, 81, 1349–1357. [Google Scholar] [CrossRef]
- Bahramisharif, A.; Rose, L.E. Efficacy of Biological Agents and Compost on Growth and Resistance of Tomatoes to Late Blight. Planta 2019, 249, 799–813. [Google Scholar] [CrossRef]
- Fry, W. Phytophthora infestans: The Plant (and R Gene) Destroyer. Mol. Plant Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef]
- Nelson, S.C. Late Blight of Tomato (Phytophthora infestans); University of Hawaii: Honolulu, HI, USA, 2008. [Google Scholar]
- Leesutthiphonchai, W.; Vu, A.L.; Ah-Fong, A.M.V.; Judelson, H.S. How Does Phytophthora infestans Evade Control Efforts? Modern Insight into the Late Blight Disease. Phytopathology 2018, 108, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, P.; Singh, P.; Kethiravan, D.; Ramathani, I.; Ramakrishnan, N. Late Blight in Tomato: Insights into the Pathogenesis of the Aggressive Pathogen Phytophthora infestans and Future Research Priorities. Planta 2021, 253, 119. [Google Scholar] [CrossRef] [PubMed]
- Commission Implementing Regulation (EU) 2018/1981—Of 13 December 2018—Renewing the Approval of the Active Substances Copper Compounds, as Candidates for Substitution, in Accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council Concerning the Placing of Plant Protection Products on the Market, and Amending the Annex to Commission Implementing Regulation (EU) No 540/2011. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32018R1981&rid=3 (accessed on 29 September 2023).
- Jacob, D.; David, D.R.; Sztjenberg, A.; Elad, Y. Conditions for Development of Powdery Mildew of Tomato Caused by Oidium neolycopersici. Phytopathology 2008, 98, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.; Whipps, J.M.; Gurr, S.J. The Tomato Powdery Mildew Fungus Oidium neolycopersici. Mol. Plant Pathol. 2001, 2, 303–309. [Google Scholar] [CrossRef]
- Stevanović, M.; Stanković, I.; Vučurović, A.; Dolovac, N.; Pfaf-Dolovac, E.; Krstić, B.; Bulajić, A. First Report of Oidium neolycopersici on Greenhouse Tomatoes in Serbia. Plant Dis. 2012, 96, 912. [Google Scholar] [CrossRef]
- Fletcher, J.T.; Smewin, B.J.; Cook, R.T.A. Tomato Powdery Mildew. Plant Pathol. 1988, 37, 594–598. [Google Scholar] [CrossRef]
- Zheng, Z.; Nonomura, T.; Bóka, K.; Matsuda, Y.; Visser, R.G.F.; Toyoda, H.; Kiss, L.; Bai, Y. Detection and Quantification of Leveillula taurica Growth in Pepper Leaves. Phytopathology 2013, 103, 623–632. [Google Scholar] [CrossRef]
- Llorens, E.; Agustí-Brisach, C.; González-Hernández, A.I.; Troncho, P.; Vicedo, B.; Yuste, T.; Orero, M.; Ledó, C.; García-Agustín, P.; Lapeña, L. Bioassimilable Sulphur Provides Effective Control of Oidium neolycopersici in Tomato, Enhancing the Plant Immune System. Pest Manag. Sci. 2017, 73, 1017–1023. [Google Scholar] [CrossRef]
- Ma, M.; Taylor, P.W.J.; Chen, D.; Vaghefi, N.; He, J.-Z. Major Soilborne Pathogens of Field Processing Tomatoes and Management Strategies. Microorganisms 2023, 11, 263. [Google Scholar] [CrossRef]
- Achuo, A.E.; Audenaert, K.; Meziane, H.; Höfte, M. The SA-Dependent Defense Pathway Is Active against Different Pathogens in Tomato and Tobacco. Meded. Rijksuniv. Gent Fak. Van Landbouwkd. Toegepaste Biol. Wet. 2002, 67, 149–157. [Google Scholar]
- Block, A.; Schmelz, E.; O’Donnell, P.J.; Jones, J.B.; Klee, H.J. Systemic Acquired Tolerance to Virulent Bacterial Pathogens in Tomato. Plant Physiol. 2005, 138, 1481–1490. [Google Scholar] [CrossRef]
- dos Santos, C.; Franco, O.L. Pathogenesis-Related Proteins (PRs) with Enzyme Activity Activating Plant Defense Responses. Plants 2023, 12, 2226. [Google Scholar] [CrossRef]
- Pascual, S.; Nombela, G.; Avilés, M.; Muñiz, M. Induced Resistance in Tomato to Whitefly Bemisia tabaci by Bion. Integr. Control Prot. Crops Mediterr. Clim. 2003, 26, 61–64. [Google Scholar]
- Petrov, N.; Andonova, R. Bion and Exin as Sar Elicitors against Potato Virus Y Infection in Tomato. Sci. Technol. 2012, 2, 46–49. [Google Scholar]
- Faino, L.; Carli, P.; Testa, A.; Cristinzio, G.; Frusciante, L.; Ercolano, M.R. Potato R1 Resistance Gene Confers Resistance against Phytophthora infestans in Transgenic Tomato Plants. Eur. J. Plant Pathol. 2010, 128, 233–241. [Google Scholar] [CrossRef]
- Herman, M.A.B.; Restrepo, S.; Smart, C.D. Defense Gene Expression Patterns of Three SAR-Induced Tomato Cultivars in the Field. Physiol. Mol. Plant Pathol. 2007, 71, 192–200. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef]
- Fiocchetti, F.; Caruso, C.; Bertini, L.; Vitti, D.; Saccardo, F.; Tucci, M. Over-Expression of a Pathogenesis-Related Protein Gene in Transgenic Tomato Alters the Transcription Patterns of Other Defence Genes. J. Hortic. Sci. Biotechnol. 2006, 81, 27–32. [Google Scholar] [CrossRef]
- Hermann, S.; Orlik, M.; Boevink, P.; Stein, E.; Scherf, A.; Kleeberg, I.; Schmitt, A.; Schikora, A. Biocontrol of Plant Diseases Using Glycyrrhiza glabra Leaf Extract. Plant Dis. 2022, 106, 3133–3144. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Medina, A.; Fernandez, I.; Sánchez-Guzmán, M.; Jung, S.; Pascual, J.; Pozo, M. Deciphering the Hormonal Signalling Network behind the Systemic Resistance Induced by Trichoderma harzianum in Tomato. Front. Plant Sci. 2013, 4, 206. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G.N. Plant Diseases Caused by Fungi. Plant Pathol. 2005, 4, 385–614. [Google Scholar]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U. Potato and Tomato Late Blight Caused by Phytophthora infestans: An Overview of Pathology and Resistance Breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Saville, A.; Graham, K.; Grünwald, N.J.; Myers, K.; Fry, W.E.; Ristaino, J.B. Fungicide Sensitivity of U.S. Genotypes of Phytophthora infestans to Six Oomycete-Targeted Compounds. Plant Dis. 2015, 99, 659–666. [Google Scholar] [CrossRef] [PubMed]
- de Vries, S.; von Dahlen, J.K.; Schnake, A.; Ginschel, S.; Schulz, B.; Rose, L.E. Broad-Spectrum Inhibition of Phytophthora infestans by Fungal Endophytes. FEMS Microbiol. Ecol. 2018, 94, fiy037. [Google Scholar] [CrossRef]
- Drenker, C.; El Mazouar, D.; Bücker, G.; Weißhaupt, S.; Wienke, E.; Koch, E.; Kunz, S.; Reineke, A.; Rondot, Y.; Linkies, A. Characterization of a Disease-Suppressive Isolate of Lysobacter enzymogenes with Broad Antagonistic Activity against Bacterial, Oomycetal and Fungal Pathogens in Different Crops. Plants 2023, 12, 682. [Google Scholar] [CrossRef]
- Ghadamgahi, F.; Tarighi, S.; Taheri, P.; Saripella, G.V.; Anzalone, A.; Kalyandurg, P.B.; Catara, V.; Ortiz, R.; Vetukuri, R.R. Plant Growth-Promoting Activity of Pseudomonas aeruginosa FG106 and Its Ability to Act as a Biocontrol Agent against Potato, Tomato and Taro Pathogens. Biology 2022, 11, 140. [Google Scholar] [CrossRef]
- Hamaoka, K.; Aoki, Y.; Suzuki, S. Isolation and Characterization of Endophyte Bacillus velezensis KOF112 from Grapevine Shoot Xylem as Biological Control Agent for Fungal Diseases. Plants 2021, 10, 1815. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Choi, G.J.; Lee, H.B.; Lee, S.-W.; Lim, H.K.; Jang, K.S.; Son, S.W.; Lee, S.O.; Cho, K.Y.; Sung, N.D.; et al. Some Fungal Endophytes from Vegetable Crops and Their Anti-Oomycete Activities against Tomato Late Blight. Lett. Appl. Microbiol. 2007, 44, 332–337. [Google Scholar] [CrossRef]
- Kim, B.; Han, J.W.; Thi Ngo, M.; Le Dang, Q.; Kim, J.-C.; Kim, H.; Choi, G.J. Identification of Novel Compounds, Oleanane- and Ursane-Type Triterpene Glycosides, from Trevesia palmata: Their Biocontrol Activity against Phytopathogenic Fungi. Sci. Rep. 2018, 8, 14522. [Google Scholar] [CrossRef]
- Lamsal, K.; Kim, S.W.; Lee, Y.S.K.; Kim, Y.S. Biocontrol of Late Blight and Plant Growth Promotion in Tomato Using Rhizobacterial Isolates. Microb. Ecol. Divers. 2013, 23, 897–904. [Google Scholar] [CrossRef]
- Colombo, M.; Masiero, S.; Rosa, S.; Caporali, E.; Toffolatti, S.L.; Mizzotti, C.; Tadini, L.; Rossi, F.; Pellegrino, S.; Musetti, R.; et al. NoPv1: A Synthetic Antimicrobial Peptide Aptamer Targeting the Causal Agents of Grapevine Downy Mildew and Potato Late Blight. Sci. Rep. 2020, 10, 17574. [Google Scholar] [CrossRef]
- Linkies, A.; Jacob, S.; Zink, P.; Maschemer, M.; Maier, W.; Koch, E. Characterization of Cultural Traits and Fungicidal Activity of Strains Belonging to the Fungal Genus Chaetomium. J. Appl. Microbiol. 2021, 131, 375–391. [Google Scholar] [CrossRef]
- Hasni, I.; Decloquement, P.; Demanèche, S.; Mameri, R.M.; Abbe, O.; Colson, P.; La Scola, B. Insight into the Lifestyle of Amoeba Willaertia magna during Bioreactor Growth Using Transcriptomics and Proteomics. Microorganisms 2020, 8, 771. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Zhang, A.; Ma, Q.; Xing, L. Functional Characterization of Tomato ShROP7 in Regulating Resistance against Oidium neolycopersici. Int. J. Mol. Sci. 2022, 23, 8557. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ariza, J.; Balestrini, R.; Novero, M.; Bonfante, P. Cell-Specific Gene Expression of Phosphate Transporters in Mycorrhizal Tomato Roots. Biol. Fertil. Soils 2009, 45, 845–853. [Google Scholar] [CrossRef]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The Beneficial Effect of Trichoderma Spp. on Tomato Is Modulated by the Plant Genotype. Mol. Plant Pathol. 2011, 12, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Çakır, B.; Gül, A.; Yolageldi, L.; Özaktan, H. Response to Fusarium oxysporum f.Sp. Radicis-Lycopersici in Tomato Roots Involves Regulation of SA- and ET-Responsive Gene Expressions. Eur. J. Plant Pathol. 2014, 139, 379–391. [Google Scholar] [CrossRef]
- Scott, A.J.; Knott, M. A Cluster Analysis Method for Grouping Means in the Analysis of Variance. Biometrics 1974, 30, 507–512. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Newman-Keuls Test and Tukey Test. In Encyclopedia of Research Design; Sage: Thousand Oaks, CA, USA, 2010; Volume 2, pp. 897–902. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial Number | UTC * | Copper 1.4 L/ha | 1.25 L/ha | AXP12 2.5 L/ha | 3.75 L/ha |
|---|---|---|---|---|---|
| 113.F | a | c | b | b | b |
| 114.F | a | b | b | b | b |
| 763.F | a | c | a | b | b |
| 764.F | a | d | b | c | d |
| 765.F | a | b | b | b | b |
| 766.F | a | b | b | b | b |
| 7715.F | a | b | b | b | b |
| 7716.F | a | b | b | c | d |
| Trial Number | UTC * | Copper 1.4 L/ha | 1.25 L/ha | AXP12 2.5 L/ha | 3.75 L/ha |
|---|---|---|---|---|---|
| 114.F | a | b | b | b | b |
| 763.F | a | b | b | b | b |
| 764.F | a | d | b | c | d |
| 765.F | a | c | b | c | c |
| 766.F | a | b | b | b | b |
| 7715.F | a | b | b | b | b |
| Trial Number | UTC * | Sulfur 7.5 kg/ha | 1.25 L/ha | AXP12 2.5 L/ha | 3.75 L/ha |
|---|---|---|---|---|---|
| 733A22FE2 | a | d | b | b | c |
| 112.F | a | b | b | b | b |
| 850.F | a | b | b | b | b |
| 851.F | a | c | b | b | c |
| S22-08207.F | a | c | b | c | b |
| Product | Concentration | Treatment |
|---|---|---|
| AXP10 | 0.2 g/L | First spraying |
| Second spraying | ||
| Third spraying | ||
| AXP10 | 1 g/L | First spraying |
| Second spraying | ||
| Third spraying | ||
| AXP12 | 5 g/L | First spraying |
| Second spraying | ||
| Third spraying | ||
| Water | / | First spraying |
| Water | / | Second spraying |
| Bion® | 0.015% | Third spraying |
| Water | / | First spraying |
| Second spraying | ||
| Third spraying | ||
| Water | / | First spraying |
| Water | / 1 mM | Second spraying |
| Salicylic acid | Third spraying |
| Target Gene | Primer Name | Primer Sequence | Reference |
|---|---|---|---|
| 18S-1 | Forward | AAAAGGTCGACGCGGGCT | [49] |
| Reverse | CGACAGAAGGGACGAGAC | ||
| act | Forward | CACCACTGCTGAACGGGAA | [50] |
| Reverse | GGAGCTGCTCCTGGCAGTTT | ||
| loxA | Forward | TGAACCATGGTGGGCTGAAA | [50] |
| Reverse | CTGCCCGAAATTGACTGCTG | ||
| PR1-3 | Forward | GCACTAAACCTAAAGAAAAATGGG | [50] |
| Reverse | AAGTTGGCATCCCAAGACATA | ||
| PR4-1 | Forward | ATGGGGTTGTTCAACATCTCATTGTTACT | [51] |
| Reverse | TTAATAAGGACGTTCTCCAACCCAGTT | ||
| PR5-1 | Forward | CCCCAACAAAACCTAGTGGA | [32] |
| Reverse | ACCAGGGCAAGTAAATGTGC |
| Trial Reference | Type | Country | Variety |
|---|---|---|---|
| 113.F | Open Field | Italy | Fokker |
| 114.F | Open Field | Italy | Nunhems 6438 |
| 763.F | Open Field | Italy | Fokker |
| 764.F | Open Field | Italy | Heinz |
| 765.F | Open Field | Italy | Heinz |
| 766.F | Greenhouse | Italy | Sir Elyan |
| 7715.F | Open Field | Spain | Encomienda |
| 7716.F | Greenhouse | Spain | Huevo de Toro |
| Modality | Product | Form | CONC. | Rate (kg/ha) | AS | Rate (g/ha) |
|---|---|---|---|---|---|---|
| 1 | UTC | |||||
| 2 | TBCS | WG | 40% | 2.00 | copper | 800 |
| 6 | AXP12 | SC | 20% | 1.25 | AXP10 | 250 |
| 7 | AXP12 | SC | 20% | 2.50 | AXP10 | 500 |
| 8 | AXP12 | SC | 20% | 3.75 | AXP10 | 750 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troussieux, S.; Gilgen, A.; Souche, J.-L. Fighting Tomato Fungal Diseases with a Biocontrol Product Based on Amoeba Lysate. Plants 2023, 12, 3603. https://doi.org/10.3390/plants12203603
Troussieux S, Gilgen A, Souche J-L. Fighting Tomato Fungal Diseases with a Biocontrol Product Based on Amoeba Lysate. Plants. 2023; 12(20):3603. https://doi.org/10.3390/plants12203603
Chicago/Turabian StyleTroussieux, Sandrine, Annabelle Gilgen, and Jean-Luc Souche. 2023. "Fighting Tomato Fungal Diseases with a Biocontrol Product Based on Amoeba Lysate" Plants 12, no. 20: 3603. https://doi.org/10.3390/plants12203603