
The Impact of Water Potential and Temperature on Native Species’ Capability for Seed Germination in the Loess Plateau Region, China

Abstract
:1. Introduction
2. Results
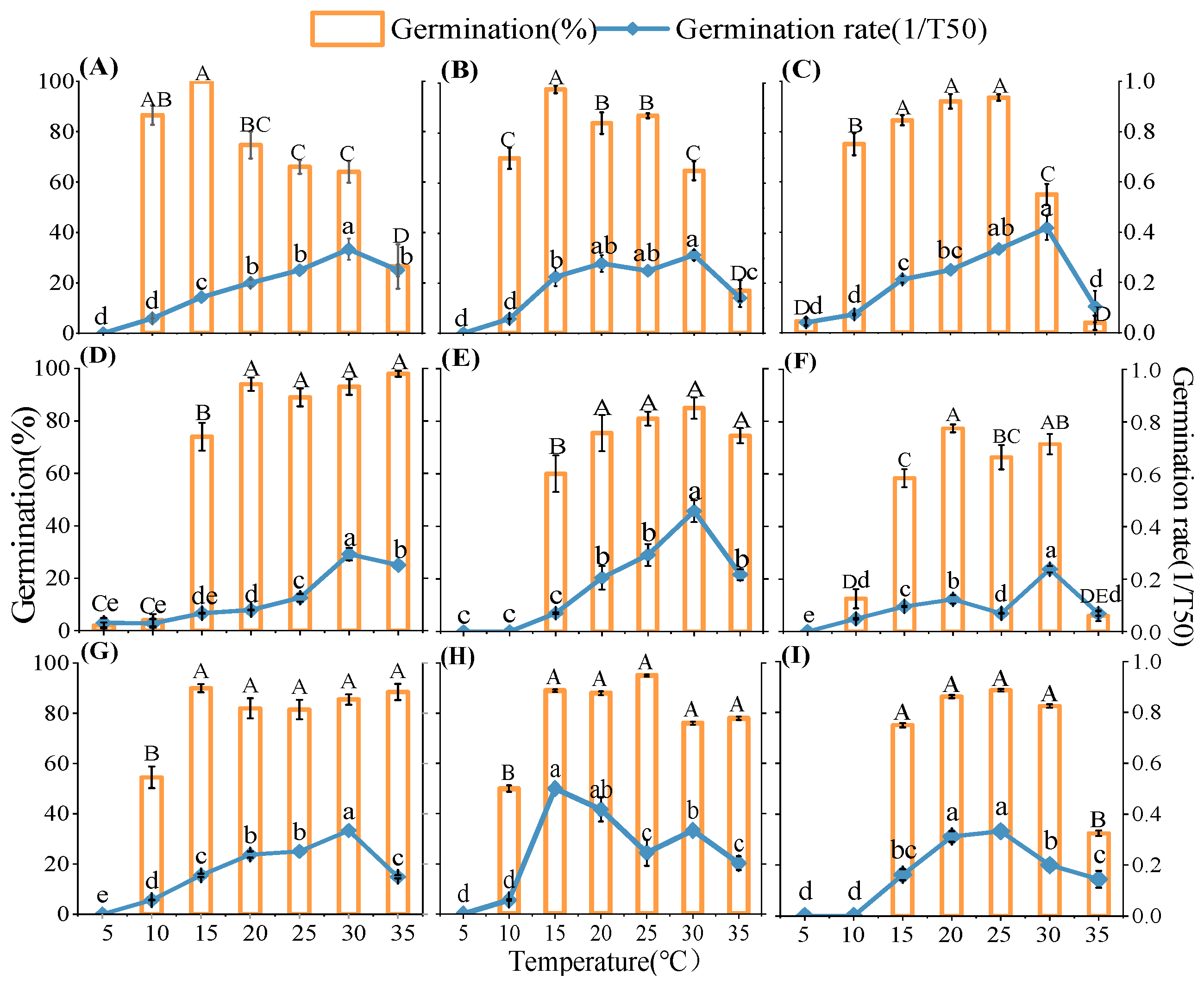
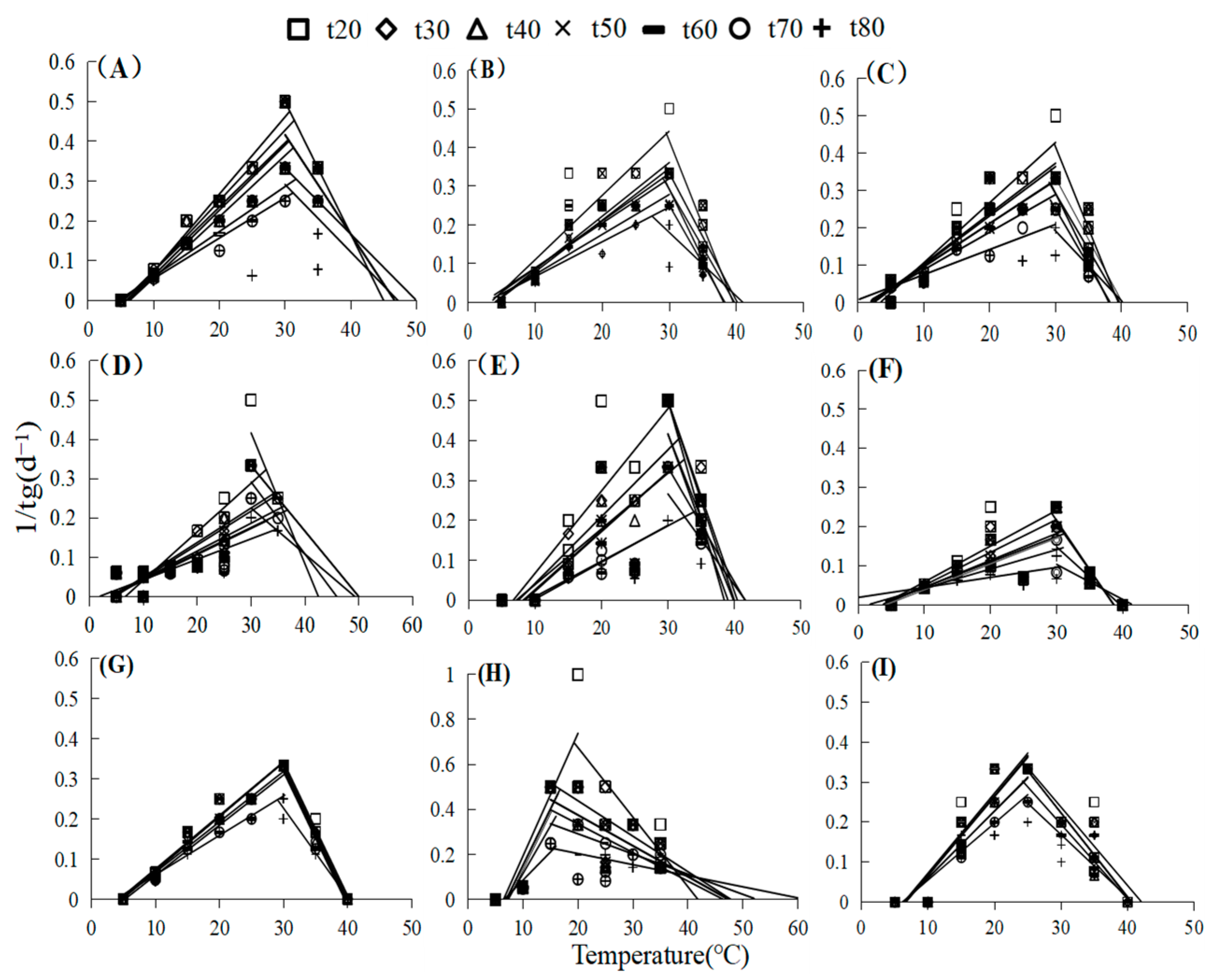
2.1. Germination Responses to Temperature
2.2. Germination Responses to the Water Potential
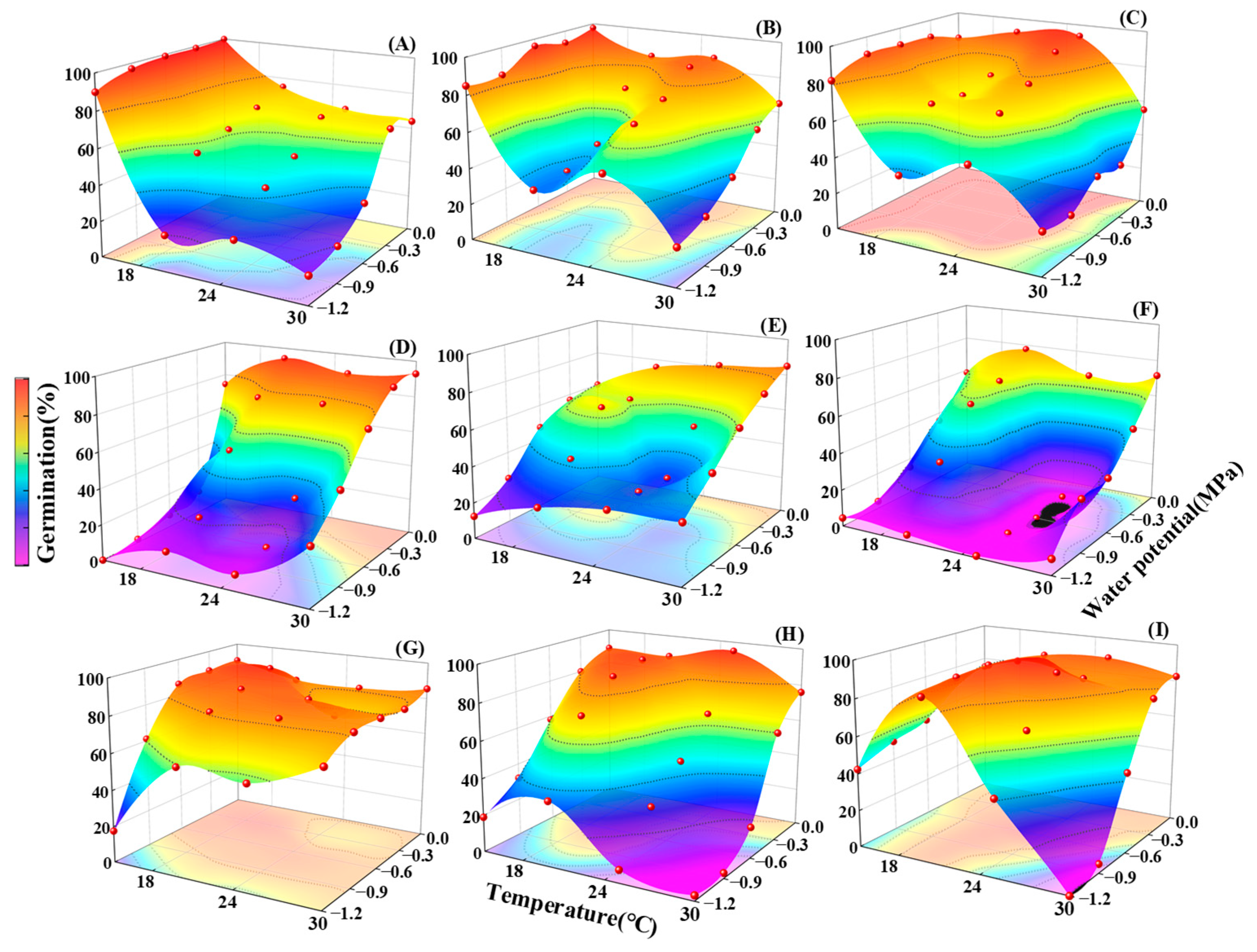
2.3. Germination Responses to the Interaction of Temperature and Water Potential
2.4. Relationships between Germination Thresholds and Seed Traits
3. Discussion
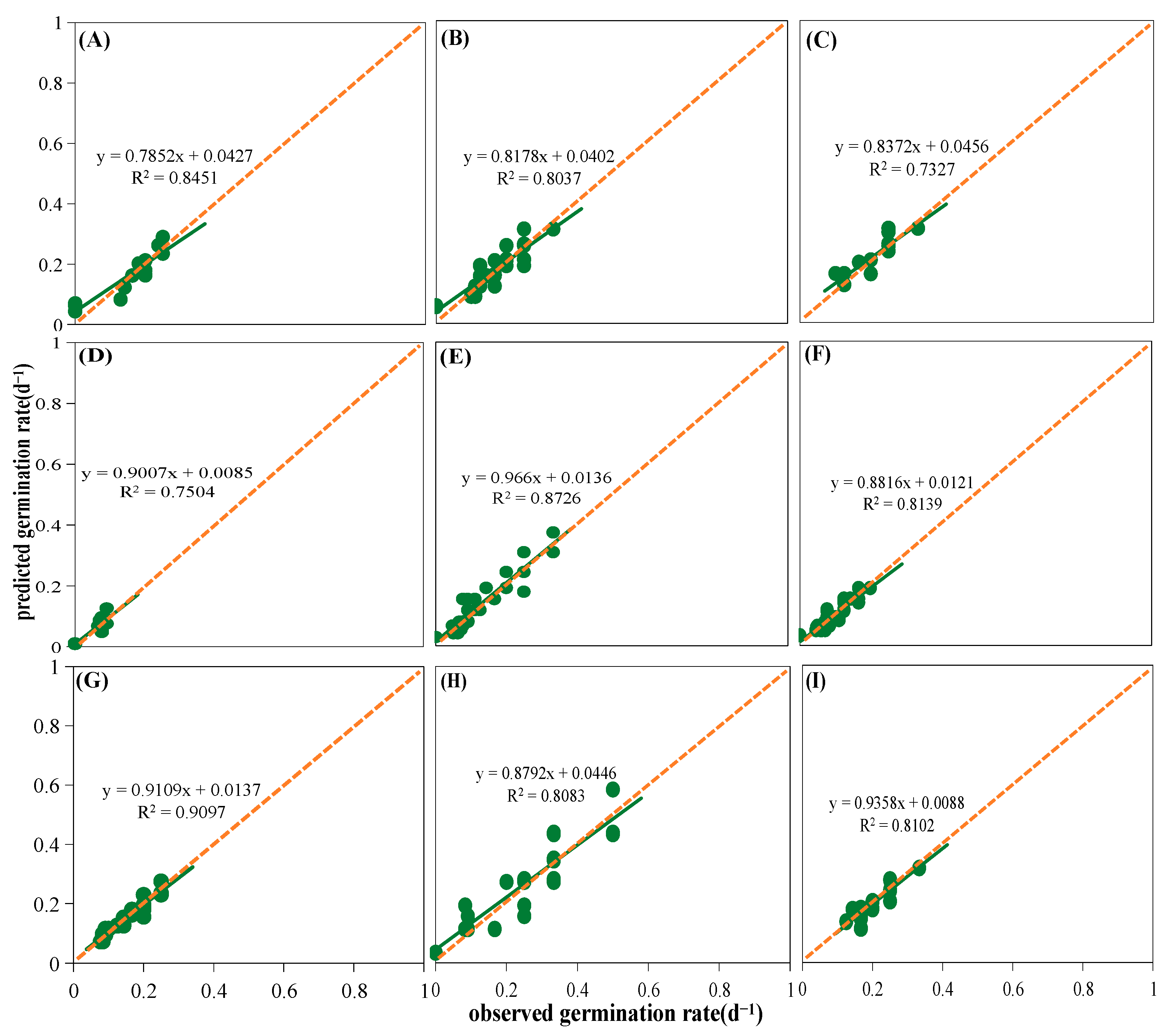
3.1. Effects of Temperature and Water Potential and Their Interaction in Seed Germination
3.2. Ecological Correlates of Seed Performance
3.3. Implications for Management and Conservation
4. Materials and Methods
4.1. Study Site
4.2. Seed Collection and Preparation
4.3. Experimental Design
4.3.1. Temperature Effects
4.3.2. Water Potential Effects
4.3.3. Interactions of Temperature and Water Potential
4.4. Mathematical Models
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sourav, M.; Ashok, K.M. Increase in Compound Drought and Heatwaves in a Warming World. Geophys. Res. Lett. 2021, 48, e2020GL090617. [Google Scholar] [CrossRef]
- Kumar Puran, T.; Sourav, M.; Ashok, K.M.; Michael, M.; Williams, A.P. Climate change will accelerate the high-end risk of compound drought and heatwave events. Proc. Natl. Acad. Sci. USA 2023, 120, e2219825120. [Google Scholar] [CrossRef]
- Liu, M.; Qiao, N.; Zhang, B.; Liu, F.; Miao, Y.; Chen, J.; Sun, Y.; Wang, P.; Wang, D. Differential responses of the seed germination of three functional groups to low temperature and darkness in a typical steppe, Northern China. PeerJ 2022, 10, e14485. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Change Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Sun, G.Q.; Li, L.; Li, J.; Liu, C.; Wu, Y.P.; Gao, S.; Wang, Z.; Feng, G.L. Impacts of climate change on vegetation pattern: Mathematical modeling and data analysis. Phys. Life Rev. 2022, 43, 239–270. [Google Scholar] [CrossRef]
- Salete Capellesso, E.; Cequinel, A.; Marques, R.; Luisa Sausen, T.; Bayer, C.; Marques, M.C.M. Co-benefits in biodiversity conservation and carbon stock during forest regeneration in a preserved tropical landscape. For. Ecol. Manag. 2021, 492, 119222. [Google Scholar] [CrossRef]
- Dalgleish, H.J.; Koons, D.N.; Adler, P.B. Can life-history traits predict the response of forb populations to changes in climate variability? J. Ecol. 2010, 98, 209–217. [Google Scholar] [CrossRef]
- Gurvich, D.E.; Pérez-Sánchez, R.; Bauk, K.; Jurado, E.; Ferrero, M.C.; Funes, G.; Flores, J. Combined effect of water potential and temperature on seed germination and seedling development of cacti from a mesic Argentine ecosystem. Flora 2017, 227, 18–24. [Google Scholar] [CrossRef]
- Pautasso, M.; Dehnen-Schmutz, K.; Holdenrieder, O.; Pietravalle, S.; Salama, N.; Jeger, M.J.; Lange, E.; Hehl-Lange, S. Plant health and global change—Some implications for landscape management. Biol. Rev. Camb. Philos. Soc. 2010, 85, 729–755. [Google Scholar] [CrossRef] [PubMed]
- Bakhshandeh, E.; Pirdashti, H.; Vahabinia, F.; Gholamhossieni, M. Quantification of the Effect of Environmental Factors on Seed Germination and Seedling Growth of Eruca (Eruca sativa) Using Mathematical Models. J. Plant Growth Regul. 2019, 39, 190–204. [Google Scholar] [CrossRef]
- Jay Ram, L.; Philippe, D.; Christian, S.; Ming Pei, Y.; Martin, J.B.; Jean Noël, A. Abiotic and biotic factors affecting crop seed germination and seedling emergence: A conceptual framework. Plant Soil 2018, 432, 1–28. [Google Scholar] [CrossRef]
- Rouhollah, A.; Ahmadreza, M.; Sanam, G. Effect of environmental factors on seed germination and emergence of Lepidium vesicarium. Plant Species Biol. 2015, 31, 178–187. [Google Scholar] [CrossRef]
- Ahmadi, A. The effect of temperature, salinity and soil burial on canary grass (Phalaris minor) germination. Jordan J. Agric. Sci. 2017, 13, 1052954. [Google Scholar]
- Bao, G.; Zhang, P.; Wei, X.; Zhang, Y.; Liu, W. Comparison of the effect of temperature and water potential on the seed germination of five Pedicularis kansuensis populations from the Qinghai–Tibet plateau. Front. Plant Sci. 2022, 13, 1052954. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.W.; Fan, Y.; Baskin, C.C.; Baskin, J.M.; Wang, Y.R. Comparison of the effects of temperature and water potential on seed germination of Fabaceae species from desert and subalpine grassland. Am. J. Bot. 2015, 102, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.Z.; Rasheed, A.; El-Keblawy, A.; Gairola, S.; Phartyal, S.S.; Gul, B.; Hameed, A. Inter-provenance variation in seed germination response of a cash crop halophyte Suaeda fruticosa to different abiotic factors. Flora 2022, 292, 152079. [Google Scholar] [CrossRef]
- Rowena, L.L.; Marta, J.G.; Michael, R.; Scott, J.K.; Louise, C.; Danica, E.G.; Lucy, C.; David, A.W.; Cherry, H.; William, E.F.-S. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise. Biol. Rev. 2014, 90, 31–59. [Google Scholar] [CrossRef]
- Donohue, K.; Rubio de Casas, R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, Postgermination Adaptation, and Species Ecological Ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Cheng, J.; Huang, H.; Liu, W.; Zhou, Y.; Han, W.; Wang, X.; Zhang, Y. Unraveling the Effects of Cold Stratification and Temperature on the Seed Germination of Invasive Spartina alterniflora across Latitude. Front. Plant Sci. 2022, 13, 911804. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The Control of Seed Dormancy and Germination by Temperature, Light and Nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Ejaz Ahmad, W.; Rashid, A.; Abdul, H.; Tariq, A. Alleviation of temperature stress by nutrient management in crop plants: A review. J. Soil Sci. Plant Nutr. 2012, 12, 221–244. [Google Scholar] [CrossRef]
- Diego, B.; Roberto, L.B.-A. A framework for the interpretation of temperature effects on dormancy and germination in seed populations showing dormancy. Seed Sci. Res. 2015, 25, 147–158. [Google Scholar] [CrossRef]
- Kamel, K.; Ghorbel, M.; Mohamed, C. Modeling the influence of temperature, salt and osmotic stresses on seed germination and survival capacity of Stipa tenacissima L. Plant Biol. 2023, 157, 325–338. [Google Scholar] [CrossRef]
- Bertuzzi, T.; Pastrana-Ignes, V.; Curti, R.N.; Batlla, D.; Baskin, C.C.; Sühring, S.; Galíndez, G. Variation in thermal and hydrotime requirements for seed germination of Chaco seasonally dry forest species in relation to population environmental conditions and seed mass. Austral Ecol. 2022, 47, 1232–1247. [Google Scholar] [CrossRef]
- Sampayo-Maldonado, S.; Ordoñez-Salanueva, C.A.; Mattana, E.; Ulian, T.; Way, M.; Castillo-Lorenzo, E.; Dávila-Aranda, P.D.; Lira-Saade, R.; Téllez-Valdéz, O.; Rodriguez-Arevalo, N.I.; et al. Thermal Time and Cardinal Temperatures for Germination of Cedrela odorata L. Forests 2019, 10, 841. [Google Scholar] [CrossRef]
- Alvarado, V.; Bradford, K.J. A hydrothermal time model explains the cardinal temperatures for seed germination. Plant Cell Environ. 2002, 25, 1061–1069. [Google Scholar] [CrossRef]
- Hardegree, S.P.; Jones, T.A.; Van Vactor, S.S. Variability in thermal response of primed and non-primed seeds of squirreltail [Elymus elymoides (Raf.) Swezey and Elymus multisetus (J. G. Smith) M. E. Jones]. Ann. Bot. 2002, 89, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Dantas, B.F.; Moura, M.S.B.; Pelacani, C.R.; Angelotti, F.; Taura, T.A.; Oliveira, G.M.; Bispo, J.S.; Matias, J.R.; Silva, F.F.S.; Pritchard, H.W.; et al. Rainfall, not soil temperature, will limit the seed germination of dry forest species with climate change. Oecologia 2020, 192, 529–541. [Google Scholar] [CrossRef]
- Hosseini Sanehkoori, F.; Pirdashti, H.; Bakhshandeh, E. Quantifying water stress and temperature effects on camelina (Camelina sativa L.) seed germination. Environ. Exp. Bot. 2021, 186, 104450. [Google Scholar] [CrossRef]
- Trudgill, D.L.; Squire, G.R.; Ken, T. A thermal time basis for comparing the germination requirements of some British herbaceous plants. New Phytol. 2000, 145, 107–114. [Google Scholar] [CrossRef]
- Coyne, C.J.; Kumar, S.; von Wettberg, E.J.B.; Marques, E.; Berger, J.D.; Redden, R.J.; Ellis, T.H.N.; Brus, J.; Zablatzká, L.; Smýkal, P. Potential and limits of exploitation of crop wild relatives for pea, lentil, and chickpea improvement. Legume Sci. 2020, 2, e36. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Kamkar, B.; Jami Al-Alahmadi, M.; Mahdavi-Damghani, A.; Villalobos, F.J. Quantification of the cardinal temperatures and thermal time requirement of opium poppy (Papaver somniferum L.) seeds to germinate using non-linear regression models. Ind. Crops Prod. 2012, 35, 192–198. [Google Scholar] [CrossRef]
- Zhang, R.; Luo, K.; Chen, D.; Baskin, J.; Baskin, C.; Wang, Y.; Hu, X. Comparison of Thermal and Hydrotime Requirements for Seed Germination of Seven Stipa Species From Cool and Warm Habitats. Front. Plant Sci. 2020, 11, 560714. [Google Scholar] [CrossRef] [PubMed]
- Greve, P.; Roderick, M.L.; Ukkola, A.M.; Wada, Y. The aridity Index under global warming. Environ. Res. Lett. 2019, 14, 124006. [Google Scholar] [CrossRef]
- Miguel, B.; Manuel, D.-B.; Santiago, S.; Rocío, H.-C.; Yanchuang, Z.; Juan, G.; Nicolas, G.; Hugo, S.; Vincent, M.; Anika, L.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef]
- Guillén, S.; Terrazas, T.; De la Barrera, E.; Casas, A. Germination differentiation patterns of wild and domesticated columnar cacti in a gradient of artificial selection intensity. Genet. Resour. Crop Evol. 2010, 58, 409–423. [Google Scholar] [CrossRef]
- Chengjie, G.; Fangyan, L.; Chunhua, Z.; Defeng, F.; Kun, L.; Kai, C. Germination responses to water potential and temperature variation among provenances of Pinus yunnanensis. Flora 2021, 276–277, 151786. [Google Scholar] [CrossRef]
- Martínez-López, M.; Tinoco-Ojanguren, C.; Martorell, C. Drought tolerance increases with seed size in a semiarid grassland from southern Mexico. Plant Ecol. 2020, 221, 989–1003. [Google Scholar] [CrossRef]
- Souza, C.S.; Ramos, D.M.; Barbosa, E.R.M.; Borghetti, F. Germination of grass species from dry and wet grasslands in response to osmotic stress under present and future temperatures. Am. J. Bot. 2022, 109, 2018–2029. [Google Scholar] [CrossRef]
- Bradford, K.J. A Water Relations Analysis of Seed Germination Rates. Plant Physiol. 1990, 94, 840–849. [Google Scholar] [CrossRef]
- Kent, J.B. Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Esmaeil, B.; Ali Jabraeil, J.M. Population-based threshold models: A reliable tool for describing aged seeds response of rapeseed under salinity and water stress. Environ. Exp. Bot. 2020, 176, 104077. [Google Scholar] [CrossRef]
- Romano, A.; Bravi, R. Hydrotime model to evaluate the effects of a set of priming agents on seed germination of two leek cultivars under water stress. Seed Sci. Technol. 2021, 49, 159–174. [Google Scholar] [CrossRef]
- Edson, S.; Massanori, T.; Victor José Mendes, C. Germination response of Hylocereus setaceus (Salm-Dyck ex DC:) Ralf Bauer (Cactaceae) seeds to temperature and reduced water potentials. Braz. J. Biol. 2010, 70, 135–144. [Google Scholar] [CrossRef]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef]
- D’Aguillo, M.C.; Edwards, B.R.; Donohue, K. Can the Environment have a Genetic Basis? A Case Study of Seedling Establishment in Arabidopsis thaliana. J. Hered. 2019, 110, 467–478. [Google Scholar] [CrossRef]
- Nur, M.; Baskin, C.C.; Lu, J.J.; Tan, D.Y.; Baskin, J.M. A new type of non-deep physiological dormancy: Evidence from three annual Asteraceae species in the cold deserts of Central Asia. Seed Sci. Res. 2014, 24, 301–314. [Google Scholar] [CrossRef]
- Veselá, A.; Duongová, L.; Münzbergová, Z. Plant origin and trade-off between generative and vegetative reproduction determine germination behaviour of a dominant grass species along climatic gradients. Flora 2022, 297, 152177. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Jamali, M. Halothermal and hydrothermal time models describe germination responses of canola seeds to ageing. Plant Biol. 2021, 23, 621–629. [Google Scholar] [CrossRef]
- Alinia, M.; Jalali, A.H.; Kazemeini, S.A.; Bakhshandeh, E. Modeling seed germination response of maize with different shapes and sizes using halotime and halothermal time concept. Acta Physiol. Plant. 2022, 44, 133. [Google Scholar] [CrossRef]
- Ambika, S.; Manonmani, V.; Somasundar, G. Review on Effect of Seed Size on Seedling Vigour and Seed Yield. Res. J. Seed Sci. 2014, 7, 31–38. [Google Scholar] [CrossRef]
- Gremer, J.R.; Venable, D.L. Bet hedging in desert winter annual plants: Optimal germination strategies in a variable environment. Ecol. Lett. 2014, 17, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Susko, D.J.; Cavers, P.B. Seed size effects and competitive ability in Thlaspi arvense L. (Brassicaceae). Botany 2008, 86, 259–267. [Google Scholar] [CrossRef]
- Wang, G.; Lynch, A.L.; Cruz, V.M.V.; Heinitz, C.C.; Dierig, D.A. Temperature requirements for guayule seed germination. Ind. Crops Prod. 2020, 157, 112934. [Google Scholar] [CrossRef]
- Liu, Y.; Hou, X.; Qiao, J.; Zhang, W.; Fang, M.; Lin, M. Evaluation of soil erosion rates in the hilly-gully region of the Loess Plateau in China in the past 60 years using global fallout plutonium. Catena 2023, 220, 106666. [Google Scholar] [CrossRef]
- Jia, X.X.; Shao, M.A.; Zhu, Y.J.; Luo, Y. Soil moisture decline due to afforestation across the Loess Plateau, China. J. Hydrol. 2017, 546, 113–122. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, T.; Wang, L.; Wang, X.; Wei, T.; Wang, P. Effects of long-term vegetation restoration on soil physicochemical properties mainly achieved by the coupling contributions of biological synusiae to the Loess Plateau. Ecol. Indic. 2023, 152, 110353. [Google Scholar] [CrossRef]
- Huang, L.; Zhao, W.; Shao, M.A. Response of plant physiological parameters to soil water availability during prolonged drought is affected by soil texture. J. Arid. Land 2021, 13, 688–698. [Google Scholar] [CrossRef]
- Hu, J.; Li, K.; Deng, C.; Gong, Y.; Liu, Y.; Wang, L. Seed Germination Ecology of Semiparasitic Weed Pedicularis kansuensis in Alpine Grasslands. Plants 2022, 11, 1777. [Google Scholar] [CrossRef]
- Zaferanieh, M.; Batool, M.; Benjamin, T. Effect of temperature and water potential on Alyssum homolocarpum seed germination: Quantification of the cardinal temperatures and using hydro thermal time. S. Afr. J. Bot. 2020, 131, 259–266. [Google Scholar] [CrossRef]
- Allison, R.K. Regulatory mechanisms involved in the transition from seed development to germination. Crit. Rev. Plant Sci. 1990, 9, 155–195. [Google Scholar] [CrossRef]
- Luis, S.-L.; Marina, G.-R.; David, D.-P.; Fernando, G.-C.; Viridiana, C.-A.; Viridiana, Z.-V.; Viridiana, L.-L.; Rafael, M.-S.; Irma, B.-L.; Sobeida, S.-N. Early carbon mobilization and radicle protrusion in maize germination. J. Exp. Bot. 2012, 63, 4513–4526. [Google Scholar] [CrossRef]
- Kazım, M.; Demir, I.; Matthews, S. Mean germination time estimates the relative emergence of seed lots of three cucurbit crops under stress conditions. Seed Sci. Technol. 2010, 38, 14–25. [Google Scholar] [CrossRef]
- Badano, E.I.; Sánchez-Montes de Oca, E.J. Seed fate, seedling establishment and the role of propagule size in forest regeneration under climate change conditions. For. Ecol. Manag. 2022, 503, 119776. [Google Scholar] [CrossRef]
- Haj Sghaier, A.; Khaeim, H.; Tarnawa, Á.; Kovács, G.P.; Gyuricza, C.; Kende, Z. Germination and Seedling Development Responses of Sunflower (Helianthus annuus L.) Seeds to Temperature and Different Levels of Water Availability. Agriculture 2023, 13, 608. [Google Scholar] [CrossRef]
- Sharma, P.; Sharma, M.M.M.; Patra, A.; Vashisth, M.; Mehta, S.; Singh, B.; Tiwari, M.; Pandey, V. Influence of high-temperature stress on rice growth and development—A review. Heliyon 2022, 8, 123–152. [Google Scholar] [CrossRef]
- Pankaj, S.; Mayur Mukut Murlidhar, S.; Anupam, P.; Medhavi, V.; Sahil, M.; Singh, B.M.; Manish, T.; Vimal, P. The Role of Key Transcription Factors for Cold Tolerance in Plants; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Wei, L.N.; Zhang, C.P.; Dong, Q.M.; Yang, Z.Z.; Chu, H.; Yu, Y.; Yang, X.X. Effects of temperature and water potential on seed germination of 13 Poa L. species in the Qinghai-Tibetan Plateau. Glob. Ecol. Conserv. 2021, 25, e01442. [Google Scholar] [CrossRef]
- Khan, W.; Shah, S.; Ullah, A.; Ullah, S.; Amin, F.; Iqbal, B.; Ahmad, N.; Abdel-Maksoud, M.A.; Okla Mk El-Zaidy, M.; Al-Qahtani, W.H.; et al. Utilizing hydrothermal time models to assess the effects of temperature and osmotic stress on maize (Zea mays L.) germination and physiological responses. BMC Plant Biol. 2023, 23, 414. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Du, G. Seed germination response to diurnally alternating temperatures: Comparative studies on alpine and subalpine meadow populations. Glob. Ecol. Conserv. 2023, 44, e02503. [Google Scholar] [CrossRef]
- de Lima, C.F.F.; Kleine-Vehn, J.; Kleine-Vehn, J.; Feraru, E. Getting to the root of belowground high temperature responses in plants. J. Exp. Bot. 2021, 72, 7404–7413. [Google Scholar] [CrossRef]
- Mahtab, N.; Arash, M.; Hoseine, S.M.B. The evaluation response of onion (Allium cepa) seed germination to temperature by Thermal-time analysis and determine cardinal temperatures by using nonlinear regression. J. Crop Sci. 2017, 48, 961–971. [Google Scholar] [CrossRef]
- Farahinia, P.; Sadat-Noori, S.A.; Mortazavian, M.M.; Soltani, E.; Foghi, B. Hydrotime model analysis of Trachyspermum ammi (L.) Sprague seed germination. J. Appl. Res. Med. Aromat. Plants 2017, 5, 88–91. [Google Scholar] [CrossRef]
- Luis, M.-M.; Clare, C.; Lucy, C.; Shane, R.T.; Wolfgang, L.; Jason, C.S. Interactions between seed functional traits and burial depth regulate germination and seedling emergence under water stress in species from semi-arid environments. J. Arid. Environ. 2017, 147, 25–33. [Google Scholar] [CrossRef]
- Seyed Farhad, S.; Shirmohamadi-Aliakbarkhani, Z. Quantifying seed germination response of melon (Cucumis melo L.) to temperature and water potential: Thermal time, hydrotime and hydrothermal time models. S Afr. J. Bot. 2020, 130, 240–249. [Google Scholar] [CrossRef]
- Cochrane, A.; Hoyle, G.L.; Yates, C.J.; Wood, J.; Nicotra, A.B. Predicting the impact of increasing temperatures on seed germination among populations of Western Australian Banksia (Proteaceae). Seed Sci. Res. 2014, 24, 195–205. [Google Scholar] [CrossRef]
- Trudgill, D.L.; Perry, J.N. Thermal time and ecological strategies—A unifying hypothesis. Ann. Appl. Biol. 1994, 125, 521–532. [Google Scholar] [CrossRef]
- Trudgill, D.L.; Honek, A.; Li, D.; Straalen, N.M. Thermal time—Concepts and utility. Ann. Appl. Biol. 2005, 146, 1–14. [Google Scholar] [CrossRef]
- Stevens, N.; Seal, C.E.; Archibald, S.; Bond, W. Increasing temperatures can improve seedling establishment in arid-adapted savanna trees. Oecologia 2014, 175, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, J.; Wang, Y. Thermal time model analysis for seed germination of four Vicia species. Chin. J. Plant Ecol. 2012, 36, 841–848. [Google Scholar] [CrossRef]
- López, A.S.; López, D.R.; Arana, M.V.; Batlla, D.; Marchelli, P. Germination response to water availability in populations of Festuca pallescens along a Patagonian rainfall gradient based on hydrotime model parameters. Sci. Rep. 2021, 11, 10653. [Google Scholar] [CrossRef]
- Čanak, P.; Jeromela, A.M.; Vujošević, B.; Kiprovski, B.; Mitrović, B.; Alberghini, B.; Facciolla, E.; Monti, A.; Zanetti, F. Is Drought Stress. Tolerance Affected by Biotypes and Seed Size in the Emerging Oilseed Crop Camelina? Agronomy 2020, 10, 1856. [Google Scholar] [CrossRef]
- Bellie, S. Global climate change and its impacts on water resources planning and management: Assessment and challenges. Stoch. Environ. Res. Risk Assess. 2010, 25, 583–600. [Google Scholar] [CrossRef]
- Rosbakh, S.; Phartyal, S.S.; Poschlod, P. Seed germination traits shape community assembly along a hydroperiod gradient. Ann. Bot. 2019, 125, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, S.; Bradford, K.J.; Huxman, T.E.; Venable, D.L. The contribution of germination functional traits to population dynamics of a desert plant community. Ecology 2016, 97, 250–261. [Google Scholar] [CrossRef]
- Soares da Mota, L.A.; Garcia, Q.S. Germination patterns and ecological characteristics of Vellozia seeds from high-altitude sites in south-eastern Brazil. Seed Sci. Res. 2013, 23, 67–74. [Google Scholar] [CrossRef]
- Arène, F.; Affre, L.; Doxa, A.; Saatkamp, A. Temperature but not moisture response of germination shows phylogenetic constraints while both interact with seed mass and lifespan. Seed Sci. Res. 2017, 27, 110–120. [Google Scholar] [CrossRef]
- Antoine, G.; Françoise, C.; Marie-Hélène, W.; Carolyne, D. How do seed and seedling traits influence germination and emergence parameters in crop species? A comparative analysis. Seed Sci. Res. 2016, 26, 317–331. [Google Scholar] [CrossRef]
- Taylor, A.G. Seed Storage, Germination, Quality, and Enhancements; CABI eBooks: Wallingford, UK, 2020. [Google Scholar] [CrossRef]
- Westoby, M.; Jurado, E.; Leishman, M. Comparative evolutionary ecology of seed size. Trends Ecol. Evol. 1992, 7, 368–372. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New Forests 2017, 49, 1–34. [Google Scholar] [CrossRef]
- Wang, D.; Jiao, J.; Lei, D.; Wang, N.; Du, H.; Jia, Y. Effects of seed morphology on seed removal and plant distribution in the Chinese hill-gully Loess Plateau region. Catena 2013, 104, 144–152. [Google Scholar] [CrossRef]
- Liang, Y.; Gao, G.; Li, J.; Dunkerley, D.; Fu, B. Runoff and soil loss responses of restoration vegetation under natural rainfall patterns in the Loess Plateau of China: The role of rainfall intensity fluctuation. Catena 2023, 225, 107013. [Google Scholar] [CrossRef]
- Song, Y.; Gao, X. Effects of low temperature and nitrogen addition during cold stratification on seed germination of Korean pine (Pinus koraiensis). Can. J. For. Res. 2021, 51, 1698–1706. [Google Scholar] [CrossRef]
- Li, L.; Chen, J.; Han, X.; Zhang, W.; Shao, C. Shrubby Steppe Ecosystem. Ecosyst. China 2020, 2, 339–364. [Google Scholar] [CrossRef]
- Wang, P.; Su, X.; Zhou, Z.; Wang, N.; Liu, J.; Zhu, B. Differential effects of soil texture and root traits on the spatial variability of soil infiltrability under natural revegetation in the Loess Plateau of China. Catena 2023, 220, 106693. [Google Scholar] [CrossRef]
- Li, W.; Ma, X.; Ma, N.; Zhao, Y.; Qiao, Y.; Wang, P.; Sun, H. Effects of Grazing Intensity on Stoichiomerty of Biological Soil Crusts in the Hilly Loess Plateau region. Acta Agrestia Sin. 2021, 29, 2547–2555. [Google Scholar]
- Silveira, F.A.O.; Fernandes, G.W. Effect of light, temperature and scarification on the germination of Mimosa foliolosa (Leguminosae) seeds. Seed Sci. Technol. 2006, 34, 585–592. [Google Scholar] [CrossRef]
- Wang, D. Seed Life-History Strategies of Plants and Restoration by Seed Addition in the Hill-Gully Loess Plateau Region. Ph.D. Thesis, Northwest A&F University, Xianyang, China, 2015. [Google Scholar]
- Poesen, J. Transport of rock fragments by rill flow-a field study. Catena 1987, 8, 35–54. [Google Scholar]
- García-Fayos, P.; Bochet, E.; Artemi, C. Seed removal susceptibility through soil erosion shapes vegetation composition. Plant Soil 2010, 334, 289–297. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The Osmotic Potential of Polyethylene Glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Gummerson, R.J. The Effect of Constant Temperatures and Osmotic Potentials on the Germination of Sugar Beet. J. Exp. Bot. 1986, 37, 729–741. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Length (mm) | Width (mm) | Height (mm) | Individual Mass (g) | Shape | Appendages | FI | Seed Storage Time (Month) |
|---|---|---|---|---|---|---|---|---|---|
| Artemisia scoparia | Asteraceae | 0.662 ± 0.035 | 0.325 ± 0.019 | 0.215 ± 0.013 | 0.020 | Oval-circular | None | 2.295 | 12 |
| Artemisia giraldii | Asteraceae | 0.923 ± 0.024 | 0.420 ± 0.018 | 0.360 ± 0.020 | 0.061 | Oval | None | 1.865 | 12 |
| Artemisia sacrorum | Asteraceae | 1.091 ± 0.048 | 0.477 ± 0.014 | 0.350 ± 0.030 | 0.085 | Oval | None | 2.240 | 12 |
| Periploca sepium | Apocynaceae | 8.152 ± 0.068 | 1.820 ± 0.050 | 0.852 ± 0.009 | 5.506 | Long-circular | Hair | 5.852 | 12 |
| Bothriochloa ischaemum | Poaceae | 1.986 ± 0.085 | 0.744 ± 0.030 | 0.470 ± 0.036 | 0.432 | Long-spindle | Awn | 2.904 | 12 |
| Patrinia scabiosifolia | Caprifoliaceae | 2.244 ± 0.029 | 1.156 ± 0.024 | 1.110 ± 0.037 | 0.810 | Ellipsoid | Wing | 1.532 | 12 |
| Linum usitatissimum | Linaceae | 2.722 ± 0.106 | 1.446 ± 0.083 | 0.666 ± 0.064 | 0.849 | Long-oval | None | 3.129 | 12 |
| Lespedeza davurica | Fabaceae | 3.238 ± 0.185 | 1.770 ± 0.053 | 1.156 ± 0.038 | 2.129 | Obovate | None | 2.166 | 12 |
| Sophora davidii | Fabaceae | 3.064 ± 0.038 | 3.992 ± 0.087 | 2.952 ± 0.047 | 23.769 | Oval | None | 1.195 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, G.; He, X.; Wang, N.; Liu, J.; Zhou, Z. The Impact of Water Potential and Temperature on Native Species’ Capability for Seed Germination in the Loess Plateau Region, China. Plants 2024, 13, 693. https://doi.org/10.3390/plants13050693
Hu G, He X, Wang N, Liu J, Zhou Z. The Impact of Water Potential and Temperature on Native Species’ Capability for Seed Germination in the Loess Plateau Region, China. Plants. 2024; 13(5):693. https://doi.org/10.3390/plants13050693
Chicago/Turabian StyleHu, Guifang, Xinyue He, Ning Wang, Jun’e Liu, and Zhengchao Zhou. 2024. "The Impact of Water Potential and Temperature on Native Species’ Capability for Seed Germination in the Loess Plateau Region, China" Plants 13, no. 5: 693. https://doi.org/10.3390/plants13050693