
Morpho-Physiological Evaluation of Solanum betaceum Cav. In Vitro Cloned Plants: A Comparison of Different Micropropagation Methods
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
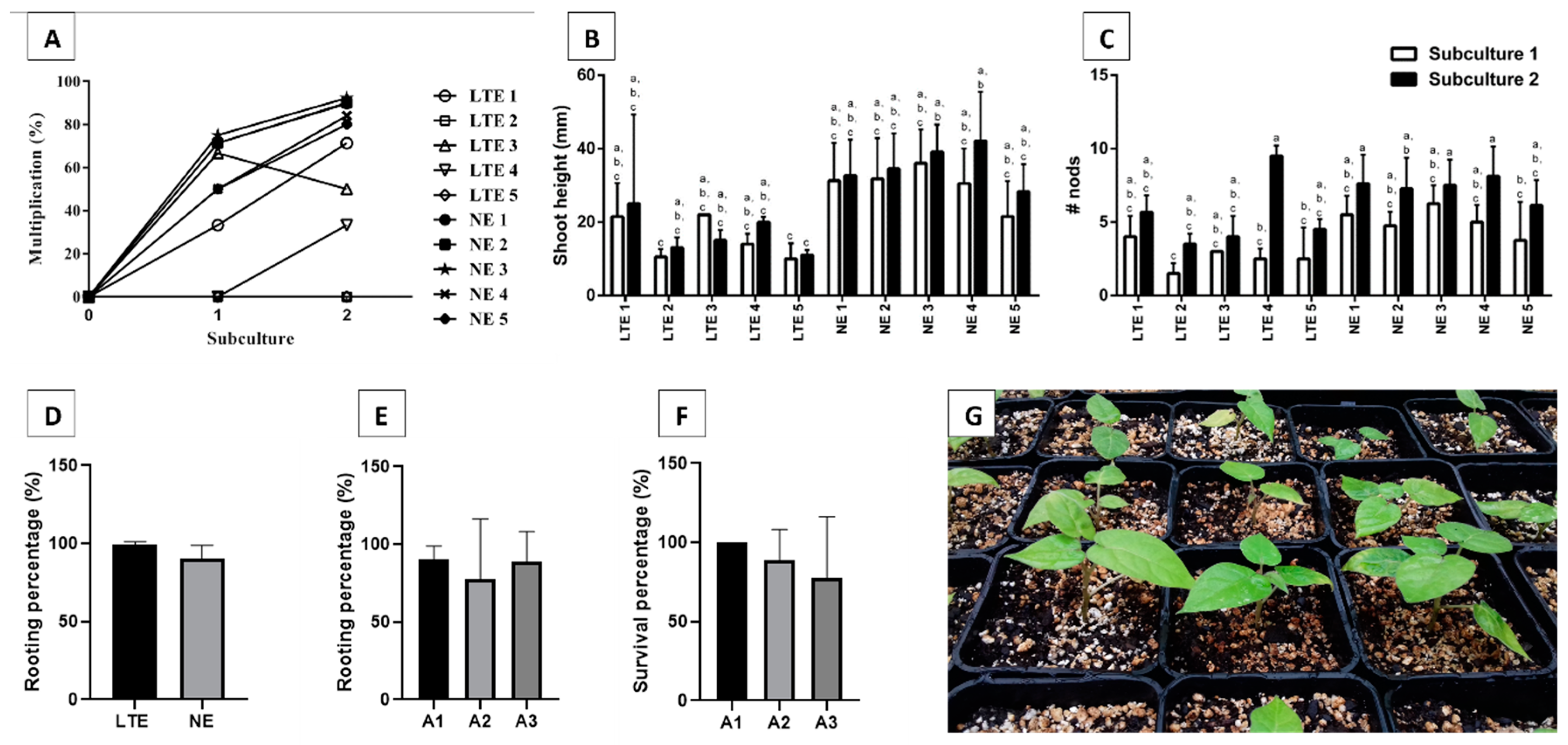
2.1. Axillary Shoot Proliferation of NE and LTE Lines, Rooting, and Acclimatization
2.2. Organogenesis Induction Shoots Elongation, Rooting, and Acclimatization
2.3. Assessment of the Embryogenic Capacity of Line ES, Embryo Conversion, and Plantlet Survival
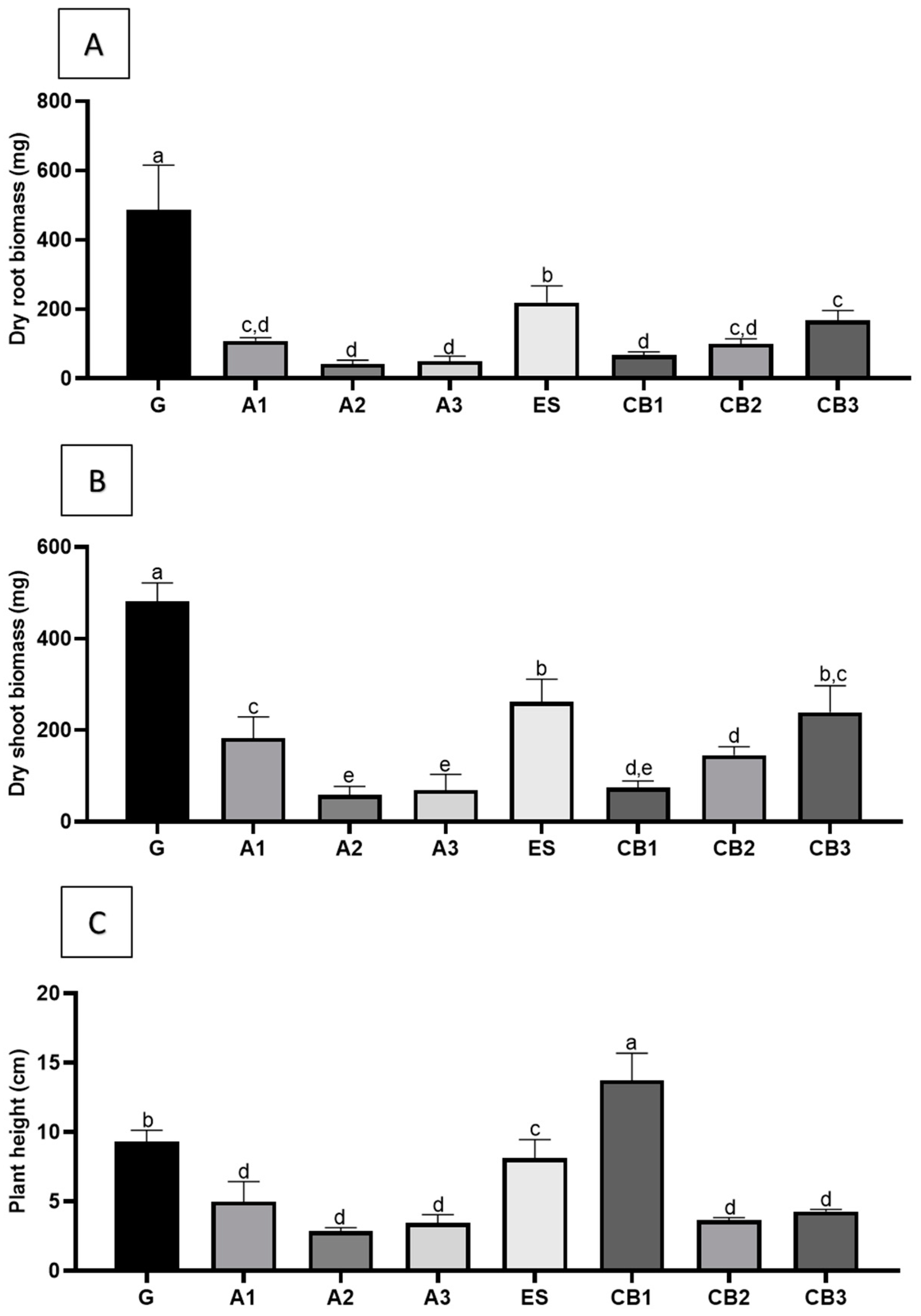
2.4. Physiological and Morphological Parameters from Plant-Derived from All the In Vitro Micropropagation Methods
3. Discussion
3.1. Axillary Shoot Proliferation and Acclimatization
3.2. Organogenesis Induction Optimization and Derived Shoots Elongation, Rooting, and Acclimatization
3.3. Embryogenic Capacity
3.4. Analysis of Physiological and Morphological Parameters of the Cloned Plants
4. Conclusions
5. Materials and Methods
5.1. Plant Material and In Vitro Establishment
5.2. Axillary Shoot Proliferation
5.2.1. Proliferation Rates of Newly Established (NE) vs. Long-Term Established (LTE) Lines
5.2.2. Rooting and Acclimatization of Micropropagated Shoots
5.3. Organogenesis
5.3.1. Organogenesis Induction
5.3.2. Shoot Elongation, Rooting, and Plant Acclimatization
5.4. Somatic Embryogenesis
5.4.1. Somatic Embryo Development from Induced Embryogenic Callus (EC)
5.4.2. Somatic Embryo Conversion and Plant Acclimatization
5.5. Seed-Derived Plants Establishment
6. Analysis of Physiological Parameters
7. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Martin, D.; Lopes, T.; Correia, S.; Canhoto, J.; Marques MP, M.; de Carvalho LA, B. Nutraceutical properties of tamarillo fruits: A vibrational study. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 252, 119501. [Google Scholar]
- Rito, M.; Marques, J.; Costa, R.; Correia, S.; Lopes, T.; Martin, D.; Canhoto, J.; Carvalho, L.; Marques, M.P. Antioxidant potential of tamarillo fruits—Chemical and infrared spectroscopy analysis. Antioxidants 2023, 12, 536. [Google Scholar] [CrossRef]
- Tridge. Available online: https://www.tridge.com/intelligences/tamarillo/export (accessed on 23 December 2022).
- Correia, S.; Braga, A.; Martins, J.; Correia, B.; Pinto, G.; Canhoto, J. Effects of polyploidy on physiological performance of acclimatized Solanum betaceum Cav. plants under water deficit. Forests 2023, 14, 208. [Google Scholar] [CrossRef]
- Guimarães, M.; Tomé, M.; Cruz, G. Cyphomandra betacea (Cav.) Sendt. (Tamarillo). In Biotechnology in Agriculture and Forestry, Trees IV; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 120–137. [Google Scholar]
- Obando, M.; Jordan, M. Regenerative responses of Cyphomandra betacea (Cav.) Sendt. (tamarillo) cultivated in vitro. Acta Hortic. 2001, 560, 429–432. [Google Scholar] [CrossRef]
- Santos, A.L. Estudos de Organogénese em Cyphomandra betacea (Cav.) Sendt. Master’s Thesis, University of Porto, Porto, Portugal, 2012. [Google Scholar]
- Guimarães, M.L.; Cruz, G.S.; Montezuma-de-Carvalho, J.M. Somatic embryogenesis and plant regeneration in Cyphomandra betacea (Cav.) Sendt. Plant Cell Tissue Organ Cult. 1988, 15, 161–167. [Google Scholar] [CrossRef]
- Correia, S.; Canhoto, J.M. Somatic embryogenesis of tamarillo (Solanum betaceum Cav.). In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Jain, S.M., Gupta, P., Eds.; Springer: Cham, Switzerland, 2018; pp. 171–179. [Google Scholar]
- Correia, S.I.; Lopes, M.L.; Canhoto, J.M. Somatic embryogenesis in tamarillo (Cyphomandra betacea): Recent advances. Acta Hortic. 2008, 839, 157–164. [Google Scholar] [CrossRef]
- Correia, S.; Lopes, M.L.; Canhoto, J.M. Somatic embryogenesis induction system for cloning an adult Cyphomandra betacea (Cav.) Sendt. (tamarillo). Trees 2011, 25, 1009–1020. [Google Scholar] [CrossRef]
- Graça, D.; Correia, S.; Ozudogru, E.A.; Lambardi, M.; Canhoto, J.M. Cryopreservation of tamarillo (Solanum betaceum Cav.) embryogenic cultures. In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants, Forestry Sciences; Jain, S.M., Gupta, P., Eds.; Springer: Cham, Switzerland, 2018; Volume 85, pp. 95–101. [Google Scholar]
- Montalbán, I.A.; de Diego, N.; Moncaleán, P. Bottlenecks in Pinus radiata somatic embryogenesis: Improving maturation and germination. Trees 2010, 24, 1061–1071. [Google Scholar] [CrossRef]
- Pullman, G.S.; Montello, P.; Cairney, J.; Xu, N.; Feng, X. Loblolly pine (Pinus taeda L.) somatic embryogenesis: Maturation improvements by metal analyses of zygotic and somatic embryos. Plant Sci. 2003, 164, 955–969. [Google Scholar]
- Egertsdotter, U.; Ahmad, I.; Clapham, D. Automation and scale up of somatic embryogenesis for commercial plant production, with emphasis on conifers. Front. Plant Sci. 2019, 10, 109. [Google Scholar]
- Loyola-Vargas, V.M. The history of somatic embryogenesis. In Somatic Embryogenesis: Fundamental Aspects and Applications; Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Springer: Cham, Switzerland, 2016; pp. 11–22. [Google Scholar]
- Vujovic, T.; Ruzic, D.; Cerovic, R. In vitro shoot multiplication as influenced by repeated subculturing of shoots of contemporary fruit rootstocks. Sci. Hortic. 2012, 39, 101–107. [Google Scholar] [CrossRef]
- Lodha, D.; Patel, A.K.; Shekhawat, N.S. A high-frequency in vitro multiplication, micromorphological studies and ex vitro rooting of Cadaba fruticosa (L.) Druce (Bahuguni): A multipurpose endangered medicinal shrub. Physiol. Mol. Biol. Plants 2015, 21, 407–415. [Google Scholar] [CrossRef]
- Zagorskaya, M.; Yegorova, N. Effect of prolonged cultivation on the micropropagation in vitro of mint cultivars and breeding samples. In BIO Web of Conferences; EDP Sciences: Les Ulis, France, 2018. [Google Scholar]
- Hegazi, G.A.; Zayed, M.S.; Salem, H.M.; Ibrahim, W.M. Effect of explant type and sequential subcultures on in vitro multiple shoots formation of Jojoba. J. Appl. Environ. Biol. Sci. 2014, 4, 214–222. [Google Scholar]
- Grigoriadou, K.; Eleni, M. Micropropagation and salt tolerance of in vitro grown Crithmum maritimum L. Plant Cell Tissue Org. Cult. 2008, 94, 209–217. [Google Scholar] [CrossRef]
- Salgado Pirata, M.; Correia, S.; Canhoto, J. Ex vitro simultaneous acclimatization and rooting of in vitro propagated tamarillo plants (Solanum betaceum Cav.): Effect of the substrate and mineral nutrition. Agronomy 2022, 12, 1082. [Google Scholar] [CrossRef]
- Waweru, D.M.; Arimi, J.M.; Marete, E.; Jacquier, J.C.; Harbourne, N. Current Status of Utilization and Potential of Dovyalis caffra Fruit: Major Focus on Kenya—A review. Sci. Afr. 2022, 16, e01097. [Google Scholar] [CrossRef]
- Anis, M.; Faisal, M. In vitro regeneration and mass multiplication of Psoralea corylifolia—An endangered medicinal plant. Indian J. Biotech. 2005, 4, 261–264. [Google Scholar]
- Barwale, U.; Kerns, H.; Widholm, J. Plant regeneration from callus cultures of several soybean genotypes via embryogenesis and organogenesis. Plant 1986, 167, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Ntui, V.; Thirukkumaran, G.; Iioka, S.; Mii, M. Efficient plant regeneration via organogenesis in “Egusi” melon (Colocynthis citrulus L.). Sci. Hoticulturae 2009, 119, 397–402. [Google Scholar] [CrossRef]
- Moyom, M.; Finnie, J.; Staden, J. Recalcitrant effects associated with the development of basal callus-like tissue on caulogenesis and rhizogenesis in Sclerocarya birrea. Plant Growth Regul. 2011, 63, 187–195. [Google Scholar] [CrossRef]
- Gomes, F.; Canhoto, J.M. Micropropagation of strawberry tree (Arbutus unedo L.) from adult plants. In Vitro Cell Dev. Biol.-Plant 2009, 45, 72–82. [Google Scholar] [CrossRef]
- Magioli, C.; Mansur, E. (Solanum melongena L.): Tissue culture, genetic transformation and use as an alternative model plant. Acta Bot. Bras. 2005, 19, 139–148. [Google Scholar] [CrossRef]
- Atkinson, R.G.; Gardner, R.C. Regeneration of transgenic tamarillo plants. Plant Cell Rep. 1993, 12, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.; Ismail, M.; Rahman, M. In vitro response from cotyledon and hypocotyls explants in tomato by inducing 6-benzylaminopurine. Afr. J. Biotechnol. 2012, 9, 4802–4807. [Google Scholar]
- Skoog, F.; Miller, C. Chemical regulation of growth and organ formation in plant tissues cultured. In Vitro Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar] [PubMed]
- Moyom, M.; Finnie, J.; Staden, J. In vitro morphogenesis of organogenic nodules derived from Scleorocarya birrea subsp. caffra leaf explants. Plant Cell Tissue Org. Cult. 2009, 98, 273–280. [Google Scholar] [CrossRef]
- Sujatha, M.; Makkar, H.; Becker, K. Shoot bud proliferation from axillary nodes and leaf sections of non-toxic Jatropha curcas L. Plant Growth Regul. 2005, 47, 83–90. [Google Scholar] [CrossRef]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.; Torphe, T. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell Dev. Biol.—Plant 1996, 32, 271–289. [Google Scholar] [CrossRef]
- Dal Vesco, L.; Guerra, M. Organogénese e micropropagação de goiabeira serrana. Rev. Bras. Frutic 1999, 21, 60–64. [Google Scholar]
- Mezzetti, B.; Savini, G.; Carnevali, F.; Mott, D. Plant genotype and growth regulators interaction affecting in vitro morphogenesis of blackberry and raspberry. Biol. Plant. 1996, 39, 139–150. [Google Scholar] [CrossRef]
- Niedz, R.P.; Evens, T.J. The effects of benzyladenine and meta Topolin on in vitro shoot regeneration of sweet orange. ARPN J. Agric. Biol. Sci. 2011, 6, 64–73. [Google Scholar]
- Bordón, Y.; Guardiola, J.L.; García Luis, A. Genotype affects the morphogenic response in vitro of epicotyl segments of Citrus rootstocks. Ann. Bot. 2000, 86, 159–166. [Google Scholar] [CrossRef]
- Moreira-Dias, J.M.; Molina, R.V.; Bordón, Y.; Guardiola, J.L.; Garcia-Luis, A. Direct and indirect shoot organogenic pathways in epicotyl cuttings of Troyer citrange differ in hormone requirements and in their response to light. Ann. Bot. 2000, 85, 103–110. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O. Meta-topolin and related cytokinins as a solution to some in vitro problems. In Meta-Topolin: A Growth Regulator for Plant Biotechnology and Agriculture; Ahmad, N., Strnad, M., Eds.; Springer: Singapore, 2021; pp. 85–91. [Google Scholar]
- Canhoto, J.M.; Lopes, M.L.; Cruz, G.S. Protocol for somatic embryogenesis: Tamarillo (Cyphomandra betacea (Cav.) Sendt.). In Protocol for Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 379–389. [Google Scholar]
- Correia, S.; Vinhas, R.; Manadas, B.; Lourenço, A.S.; Veríssimo, P.; Canhoto, J.M. Comparative proteomic analysis of auxin-induced embryogenic and nonembryogenic tissues of the solanaceous tree Cyphomandra betacea (Tamarillo). J. Proteome Res. 2012, 3, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Passamani, L.Z.E.M.; Reis, R.S.; Vale, E.M.; Sousa, K.R.; Aragão, V.P.; Santa-Catarina, C.; Silveira, V. Long-term culture with 2,4-dichlorophenoxyacetic acid affects embryogenic competence in sugarcane callus via changes in starch, polyamine and protein profiles. Plant Cell Tissue Organ. Cult. 2020, 140, 415–429. [Google Scholar] [CrossRef]
- Fehér, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tissue Org. Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Currais, L.; Loureiro, J.; Santos, C.; Canhoto, J.M. Ploidy stability in embryogenic cultures and regenerated plantlets of tamarillo. Plant Cell Tissue Org. Cult. 2013, 114, 149–159. [Google Scholar] [CrossRef]
- Lopes, M.L.; Ferreira, M.R.; Carloto, J.M.; Cruz, G.S.; Canhoto, J.M. Somatic embryogenesis induction in tamarillo (Cyphomandra betacea). In Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Newton, R.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; Volume 6, pp. 433–455. [Google Scholar]
- Bhatia, P.; Ashwath, N. Comparative performance of micropropagated and seed-grown tomato plants. Biol. Plant. 2004, 48, 625–628. [Google Scholar] [CrossRef]
- Menéndez-Yuffá, A.; Barry-Etienne, D.; Bertrand, B.; Georget, F.; Etienne, H. A comparative analysis of the development and quality of nursery plants derived from somatic embryogenesis and from seedlings for large-scale propagation of coffee (Coffea arabica L.). Plant Cell Tissue Org. Cult. 2010, 102, 297. [Google Scholar] [CrossRef]
- Maximova, S.N.; Young, A.; Pishak, S.; Guiltinan, M.J. Field performance of Theobroma cacao L. plants propagated via somatic embryogenesis. In Vitro Cell. Dev. Biol.—Plant 2008, 44, 487–493. [Google Scholar] [CrossRef]
- Maxwell, K.; Jonhson, G.N. Chorophill fluorescence—A practical guide. J. Exp. Bot. 2010, 51, 659–668. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jesus, C.; Meijón, M.; Monteiro, P.; Correia, B.; Amaral, J.; Escandón, M.; Pinto, G. Salicylic acid application modulates physiological and hormonal changes in Eucalyptus globulus under water deficit. Environ. Exp. Bot. 2015, 118, 56–66. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Average Number of Roots per Shoot | Longest Root Length (cm) | Shoot Average Length after Rooting Stage (cm) |
|---|---|---|---|
| CB1 | 2.83 ± 1.1 | 5.33 ± 0.5 | 1.75 ± 0.7 |
| CB2 | 2.87 ± 1.7 | 5.63 ± 1.7 | 2.97 ± 1.3 |
| CB3 | 3.13 ± 1.8 | 5.88 ± 1.3 | 2.75 ± 1.9 |
| CB4 | N/A | N/A | N/A |
| CB5 | 2.63 ± 1.3 | 7.31 ± 2.5 | 3.00 ± 1.0 |
| CB6 | N/A | N/A | N/A |
| Treatment | Explant | Average Number of Shoots per Explant | Average Shoot Elongation (cm) |
|---|---|---|---|
| CB1 | L | 0.5 | 1.25 ± 0.1 |
| P | 0.25 | 0.87 ± 0.3 | |
| CB2 | L | 1.375 | 1.58 ± 0.8 |
| P | 0.5 | 0.62 ± 0.4 | |
| CB3 | L | 0.875 | 0.87 ± 0.5 |
| P | 0.5 | 0.67 ± 0.6 | |
| CB4 | L | 0.125 | 0.98 ± 0.2 |
| P | 0.25 | 0.89 ± 0.6 | |
| CB5 | L | 0.625 | 1.00 ± 0.1 |
| P | 0.375 | 0.86 ± 0.6 | |
| CB6 | L | 0.25 | 1.35 ± 0.6 |
| P | 0.25 | 1.10 ± 0.3 |
| Medium | Composition |
|---|---|
| CB1 | MS + 1.0 mg·L−1 BA |
| CB2 | MS + 2.0 mg·L−1 BA |
| CB3 | MS + 3.0 mg·L−1 BA |
| CB4 | MS + 1.0 mg·L −1 BA + 2.0 mg·L−1 mT |
| CB5 | MS + 2.0 mg·L −1 BA + 1.0 mg·L−1 mT |
| CB6 | MS + 1.0 mg·L−1 BA + 0.25 mg·L−1 NAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, M.; Lopes, T.; Puga, A.P.; Pinto, G.; Canhoto, J.; Correia, S. Morpho-Physiological Evaluation of Solanum betaceum Cav. In Vitro Cloned Plants: A Comparison of Different Micropropagation Methods. Plants 2023, 12, 1884. https://doi.org/10.3390/plants12091884
Correia M, Lopes T, Puga AP, Pinto G, Canhoto J, Correia S. Morpho-Physiological Evaluation of Solanum betaceum Cav. In Vitro Cloned Plants: A Comparison of Different Micropropagation Methods. Plants. 2023; 12(9):1884. https://doi.org/10.3390/plants12091884
Chicago/Turabian StyleCorreia, Mariana, Tércia Lopes, Ana Patrícia Puga, Glória Pinto, Jorge Canhoto, and Sandra Correia. 2023. "Morpho-Physiological Evaluation of Solanum betaceum Cav. In Vitro Cloned Plants: A Comparison of Different Micropropagation Methods" Plants 12, no. 9: 1884. https://doi.org/10.3390/plants12091884