
Application of Cold Storage and Short In Vitro Germination for Somatic Embryos of Pinus radiata and P. sylvestris

Abstract
:1. Introduction
2. Results
2.1. Pinus radiata
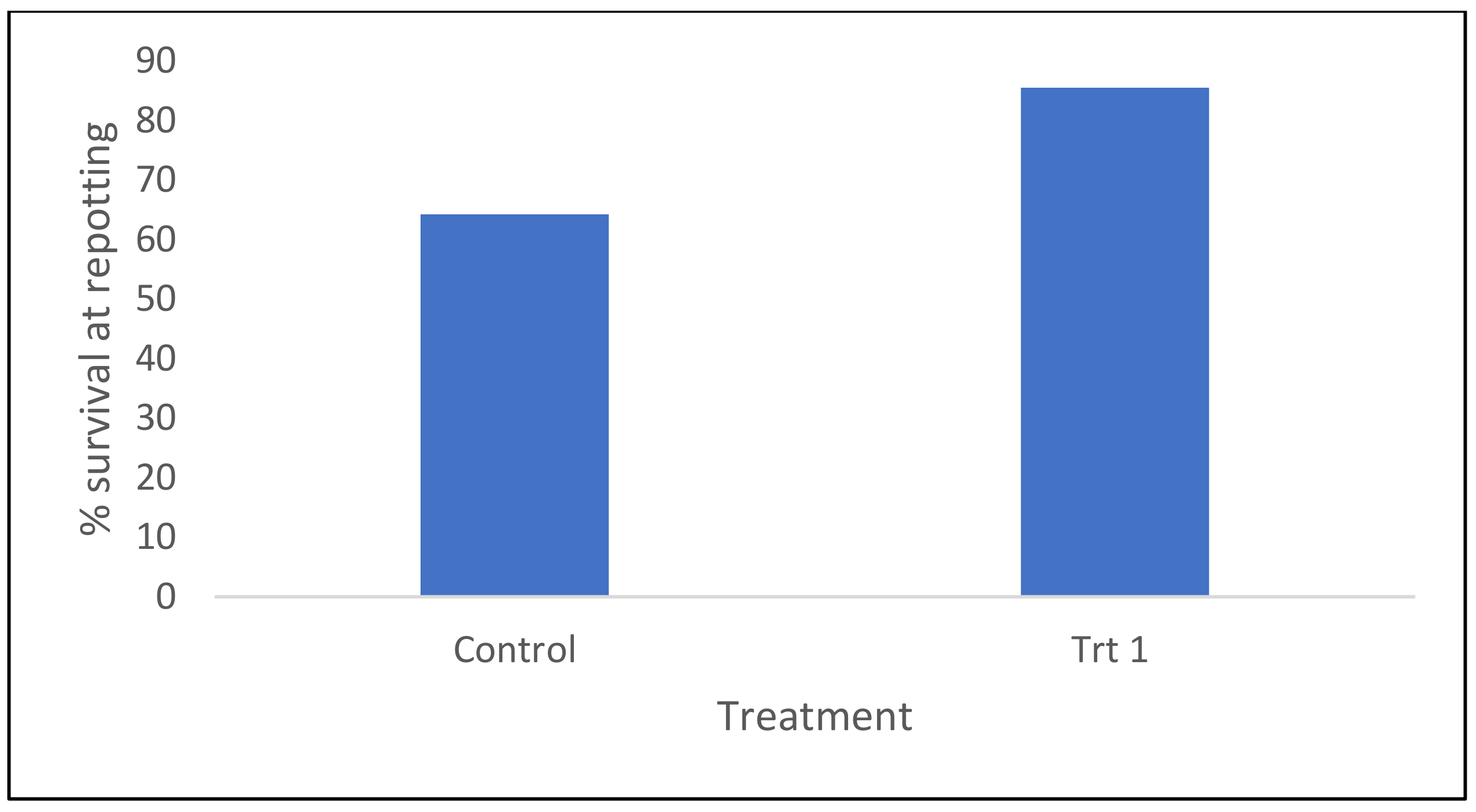
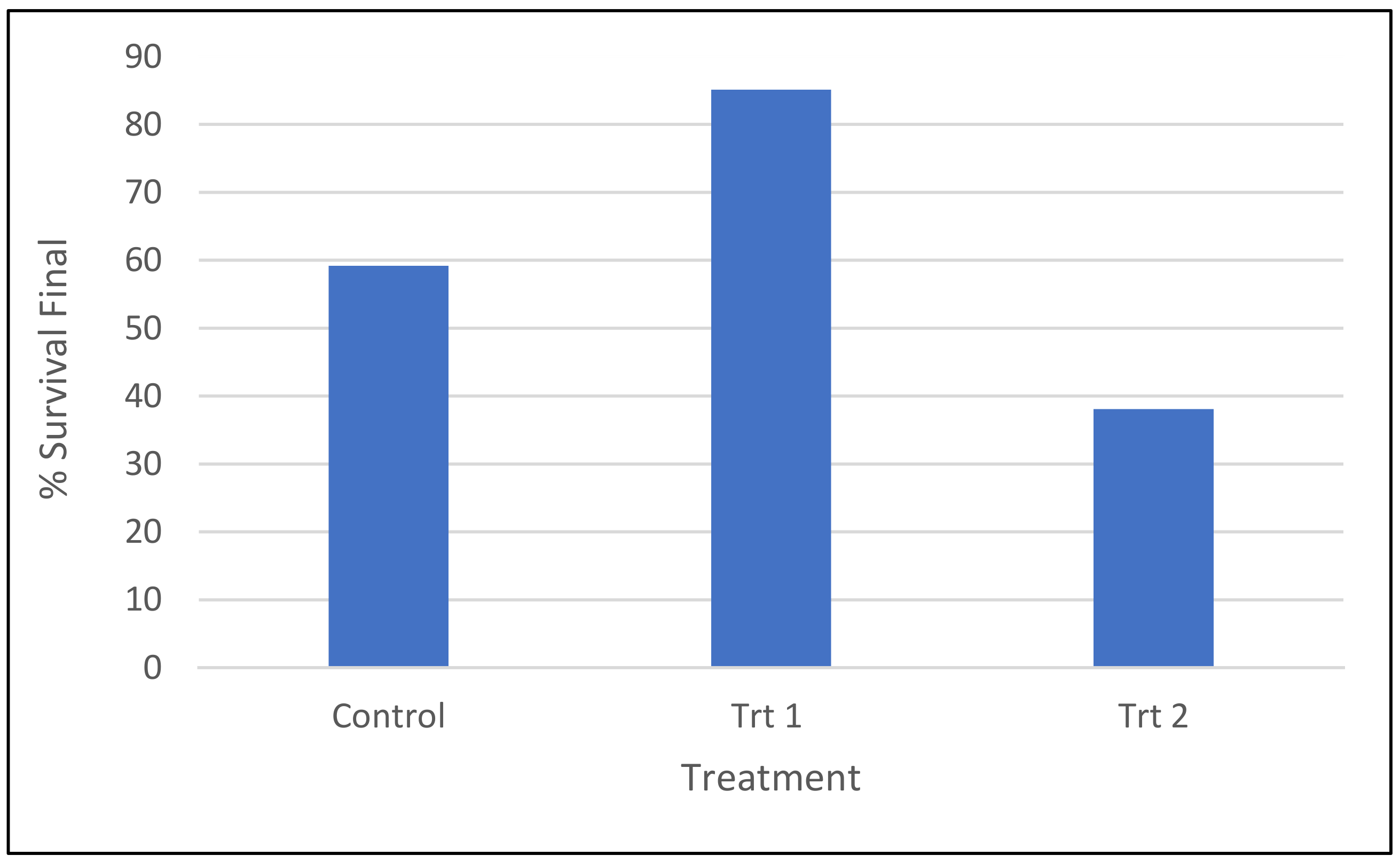
2.1.1. Survival of Somatic Emblings in Different Germination/Acclimatization Protocols
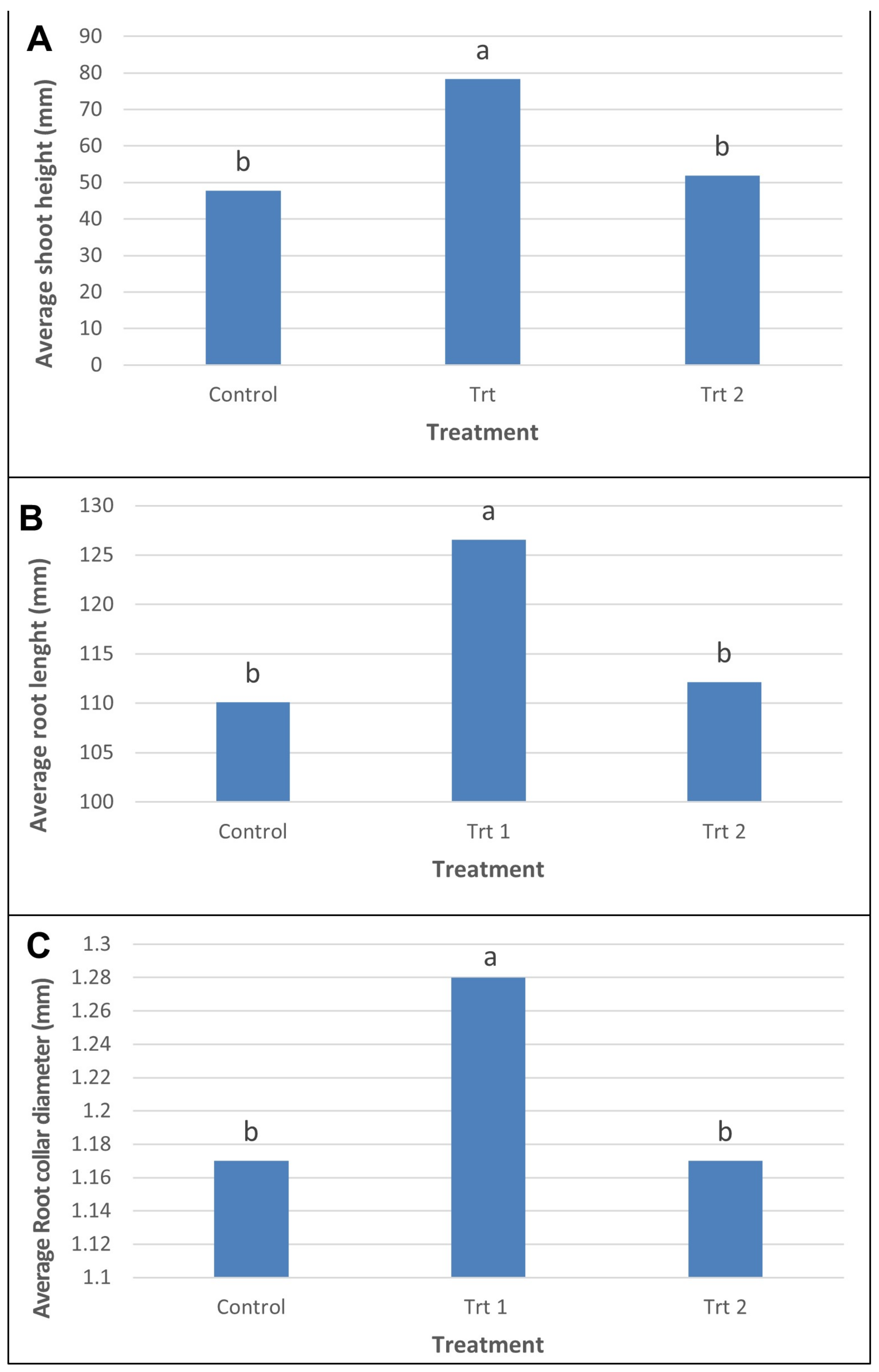
2.1.2. Growth Characteristics of Surviving Somatic Emblings
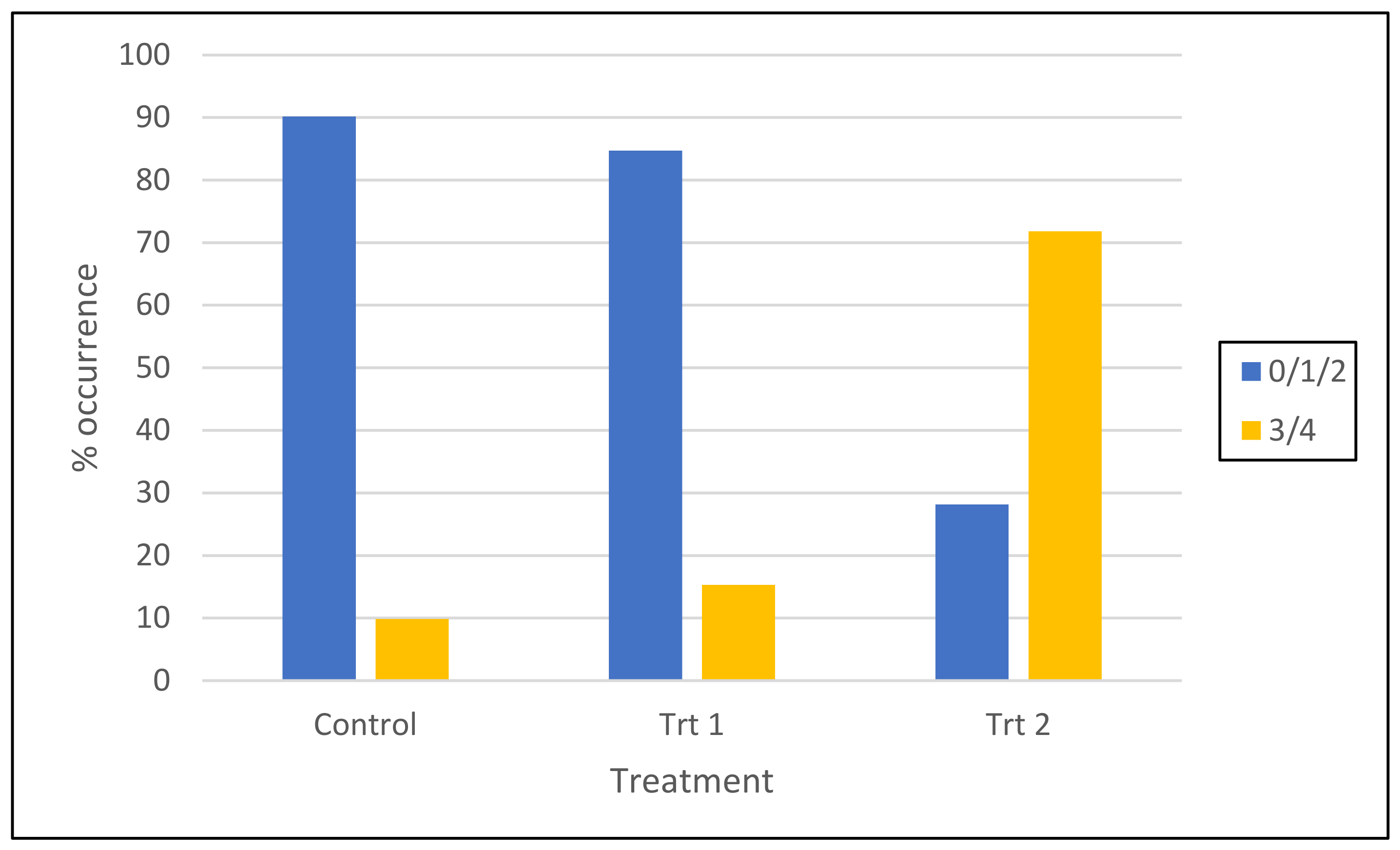
2.1.3. Balance of Root System (Root Quadrant Score) of Surviving Somatic Emblings
2.2. Pinus sylvestris
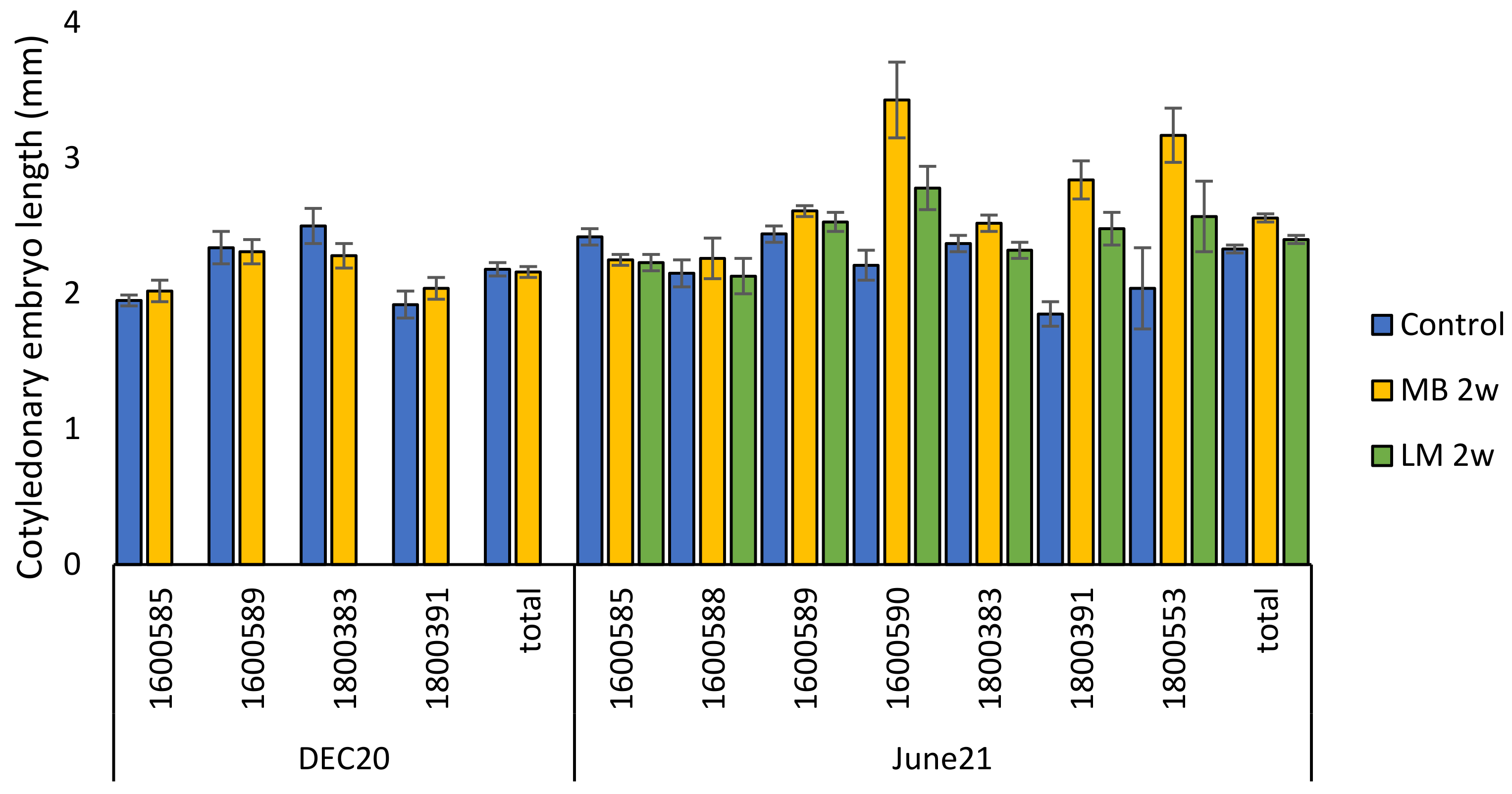
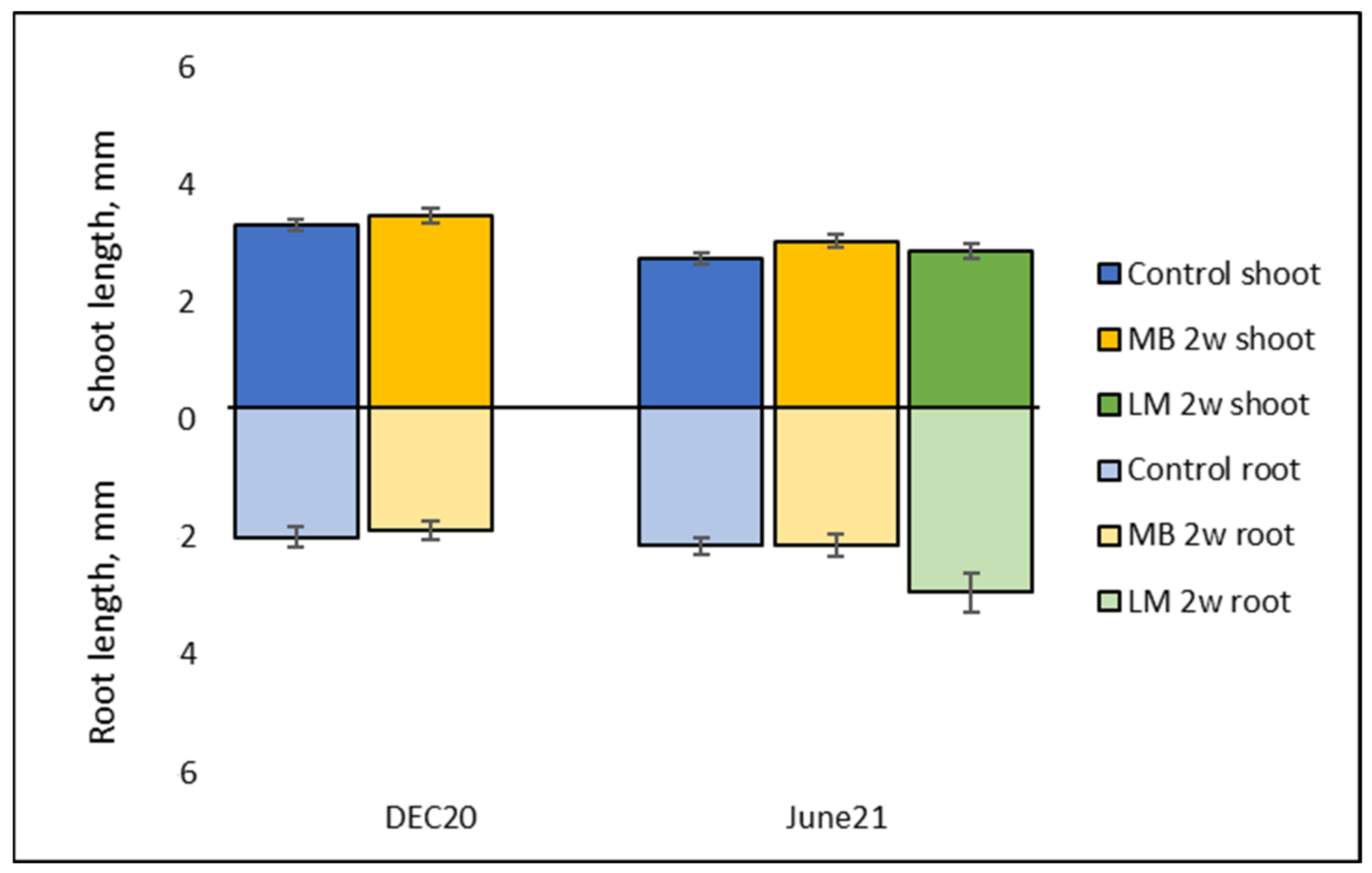
2.2.1. Embryo Size and Growth during Cold Storage and Germination
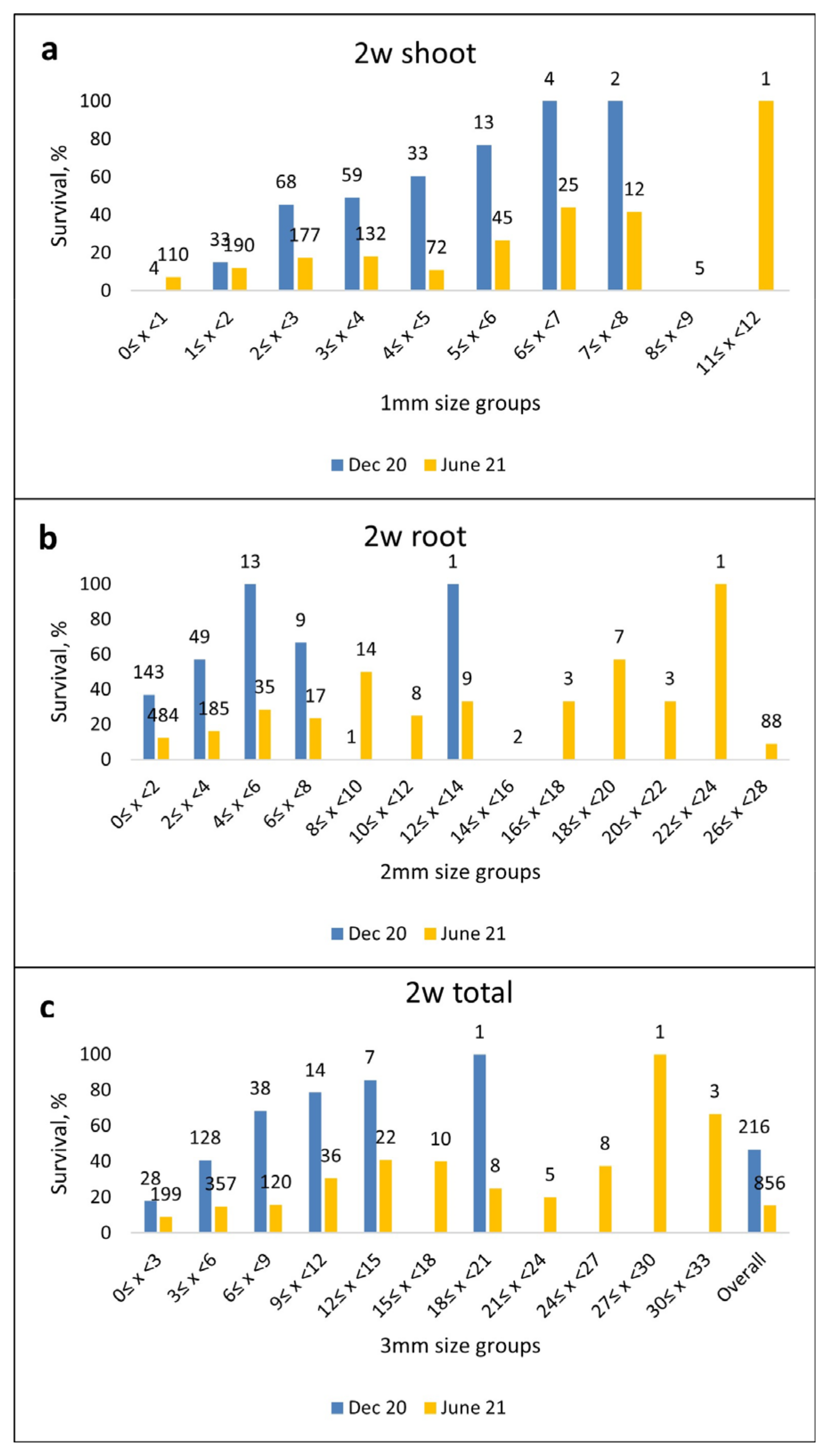
2.2.2. Survival of Somatic Emblings in Different Germination/Acclimatization Protocols
3. Discussion
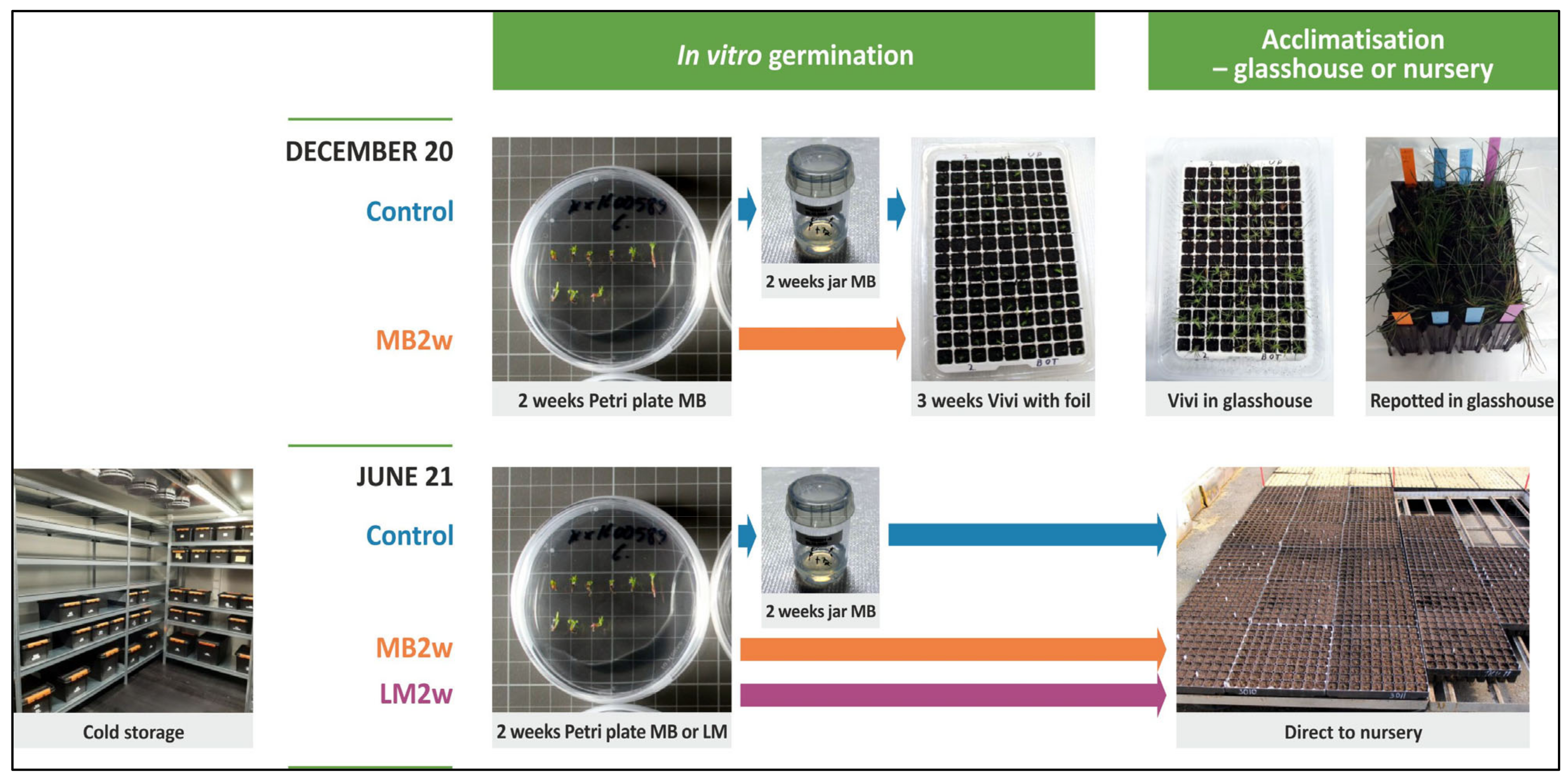
4. Materials and Methods
4.1. Pinus radiata
Plant Material
4.2. Methods
4.2.1. Maturation of Somatic Embryos
4.2.2. Germination Treatments and Further Growth of Emblings
4.2.3. Experimental Design and Statistical Evaluation
4.3. Pinus sylvestris
4.3.1. Plant Material
4.3.2. Methods
Production of Somatic Embryos and Cold Storage
Germination Treatments and Further Growth
Experimental Design and Statistical Evaluation
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forest Owners Association. Facts and Figures 2020/21. New Zealand Plantation Forest Industry. Available online: https://nzfoa.org.nz/images/FGT_3704_Facts_and_Figures_202021_Web_FA1.pdf (accessed on 25 March 2023).
- Aitken-Christie, J. Clonal propagation: Gymnosperms. Cell Cult. Somat. Cell Genet. Plants 1984, 1, 82–95. [Google Scholar]
- Smith, D.R.; Walter, C.; Warr, A.; Hargreaves, C.L.; Grace, L.J. Somatic Embryogenesis joins the plantation forestry revolution in New Zealand. In Proceedings of the TAPPI Biological Sciences Symposium, Minneapolis, MN, USA, 2–6 October 1994. [Google Scholar]
- Krakau, U.K.; Liesebach, M.; Aronen, T. Scots pine (Pinus sylvestris L.). In Forest Tree Breeding in Europe: Current State-of-the-Art and Perspectives; Paques, L.E., Ed.; Springer: Dordrecht, The Netherlands, 2013; Volume 25, pp. 267–323. [Google Scholar]
- Aronen, T. From lab to field-current state of somatic embryogenesis in Scots pine. In Vegetative Propagation of Forest Trees; Park, Y.-S., Bonga, J.M., Moon, H.-K., Eds.; National Institute of Forest Science: Seoul, Republic of Korea, 2016. [Google Scholar]
- Klimaszewska, K.; Hargreaves, C.; Lelu-Walter, M.-A.; Trontin, J.F. Advances in Conifer Somatic Embryogenesis Since Year 2000. Methods Mol. Biol. 2016, 1359, 131–166. [Google Scholar]
- Lelu-Walter, M.-A.; Klimaszewska, K.; Miguel, C.; Aronen, T.; Hargreaves, C.; Teyssier, C.; Trontin, J.-F. Somatic Embryogenesis for More Effective Breeding and Deployment of Improved Varieties in Pinus spp.: Bottlenecks and Recent Advances. In Somatic Embryogenesis: Fundamental Aspects and Applications; Springer Science and Business Media LLC: Cham, Switzerland, 2016; pp. 319–365. [Google Scholar]
- Egertsdotter, U. Plant physiological and genetical aspects of the somatic embryogenesis process in conifers. Scand. J. For. Res. 2018, 34, 360–369. [Google Scholar] [CrossRef]
- Stasolla, C.; Yeung, E. Recent advances in conifer somatic embryogenesis: Improving somatic embryo quality. Plant Cell Tissue Organ Cult. 2003, 74, 15–35. [Google Scholar] [CrossRef]
- Von Arnold, S.; Sabala, I.; Bozhkov, P.; Dyachok, J.; Filonova, L. Developmental pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult. 2002, 69, 233–249. [Google Scholar] [CrossRef]
- Tikkinen, M.; Varis, S.; Aronen, T. Development of somatic embryo maturation and growing techniques of Norway Spruce Emblings towards large-scale field testing. Forests 2018, 9, 325. [Google Scholar] [CrossRef]
- Aronen, T.; Pehkonen, T.; Ryynänen, L. Enhancement of somatic embryogenesis from immature zygotic embryos of Pinus sylvestris. Scand. J. For. Res. 2009, 24, 372–383. [Google Scholar] [CrossRef]
- Hay, E.I.; Charest, P.J. Somatic embryo germination and desiccation tolerance in conifers. In Somatic Embryogenesis in Woody Plants; Mohain Jain, S., Gupta, P.K., Newton, R.J., Eds.; Springer Science: Dordrecht, The Netherlands, 1999; pp. 61–96. [Google Scholar]
- Lelu-Walter, M.-A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Thompson, D. Challenges for the large-scale propagation of forest trees by somatic embryogenesis—A review. In Proceedings of the 3rd International Conference of the IUFRO Unit, Vitoria-Gasteiz, Spain, 12–18 September 2014. [Google Scholar]
- Tikkinen, M. Improved propagation efficiency in a laboratory-nursery interface for somatic embryogenesis in Norway spruce. Diss. For. 2018, 265, 35. [Google Scholar] [CrossRef]
- Pullman, G.S.; Johnson, S.; Peter, G.; Cairney, J.; Xu, N. Improving Loblolly Pine Somatic Embryo Maturation: Comparison of Somatic and Zygotic Embryo Morphology, Germination, and Gene Expression. Plant Cell Rep. 2003, 21, 747–758. [Google Scholar] [CrossRef]
- Lelu-Walter, M.A.; Bernier-Cardou, M.; Klimaszewska, K. Simplified and improved somatic embryogenesis for clonal propagation of Pinus pinaster (Ait.). Plant Cell Rep. 2006, 25, 767–776. [Google Scholar] [CrossRef]
- Maruyama, T.E.; Hosoi, Y. Progress in Somatic Embryogenesis of Japanese Pines. Front. Plant Sci. 2019, 10, 15. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Smith, D.R. Maturation of Somatic Embryos of Pinus strobus Is Promoted by a High Concentration of Gellan Gum. Physiol. Plant. 1997, 100, 949–957. [Google Scholar] [CrossRef]
- Montalbán, I.A.; Moncaleán, P. Rooting of Pinus radiata somatic embryos: Factors involved in the success of the process. J. For. Res. 2019, 30, 65–71. [Google Scholar] [CrossRef]
- Lara-Chavez, A.; Egertsdotter, U.; Flinn, B.S. Oocarpa Pine (Pinus oocarpa var. oocarpa Schiede). In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Mohan Jain, S., Gupta, P., Eds.; Springer: Cham, Switzerland, 2018; pp. 295–305. [Google Scholar]
- Jones, N.B.; Van Staden, J. Improved Somatic Embryo Production from Embryogenic Tissue of Pinus patula. Vitr. Cell. Dev. Biol. Plant 2001, 37, 543–549. [Google Scholar] [CrossRef]
- Tikkinen, M.; Varis, S.; Peltola, H.; Aronen, T. Improved germination conditions for Norway spruce somatic cotyledonary embryos increased survival and height growth of emblings. Trees 2018, 32, 1489–1504. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Montalbán, I.A.; Moncaleán, P. Multi-strategy approach towards optimization of maturation and germination in radiata pine somatic embryogenesis. Plant Cell Tissue Organ Cult. PCTOC 2023, 153, 173–190. [Google Scholar] [CrossRef]
- Stasolla, C.; Kong, L.; Yeung, E.C.; Thorpe, T.A. Maturation of somatic embryos in conifers: Morphogenesis, physiology, biochemistry, and molecular biology. Vitr. Cell. Dev. Biol. Plant 2002, 38, 93–105. [Google Scholar] [CrossRef]
- Llebrés, M.T.; Avila, C.; Cánovas, F.M.; Klimaszewska, K. Root growth of somatic plants of hybrid Pinus strobus (L.) and P-wallichiana (A. B. Jacks.) is affected by the nitrogen composition of the somatic embryo germination medium. Trees Struct. Funct. 2018, 32, 371–381. [Google Scholar]
- Kvaalen, H.; Appelgren, M. Light Quality Influences Germination, Root Growth and Hypocotyl Elongation in Somatic Embryos but Not in Seedlings of Norway Spruce. Vitr. Cell. Dev. Biol. Plant 1999, 35, 437–441. [Google Scholar] [CrossRef]
- Högberg, K.A.; Bozhkov, P.V.; Grönroos, R.; Arnold, S.V. Critical factors affecting ex vitro performance of somatic embryo plants of Picea abies. Scand. J. For. Res. 2001, 16, 295–304. [Google Scholar] [CrossRef]
- Hargreaves, C. The development and application of conifer tissue culture and somatic embryogenesis protocols in New Zealand: The Pinus radiata D. Don story. In Proceedings of the 4th International Conference of the IUFRO Unit 2.09.02 on “Development and Application of Vegetative Propagation Technologies in Plantation Forestry to Cope with a Changing Climate and Environment”, La Plata, Argentina, 19–23 September 2016; pp. 235–243. [Google Scholar]
- Bonga, J.M. A comparative evaluation of the application of somatic embryogenesis, rooting of cuttings, and organogenesis of conifers. Can. J. For. Res. 2015, 45, 379–383. [Google Scholar] [CrossRef]
- Denchev, P.; Grossnickle, S.C. Somatic embryogenesis for conifer seedling production: The biology of scaling. Reforesta 2019, 7, 109–137. [Google Scholar] [CrossRef]
- Wu, H.X. Benefits and risks of using clones in forestry—A review. Scand. J. For. Res. 2018, 34, 352–359. [Google Scholar] [CrossRef]
- Cervelli, R.; Senaratna, T. Economic aspects of somatic embryogenesis. In Automation and Environmental Control in Plant Tissue Culture; Aitken-Christie, J., Kozai, T., Smith, M.A.L., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 29–64. [Google Scholar]
- Carson, M.; Carson, S.; Riini, C.T. Successful varietal forestry with radiata pine in New Zealand. New Zealand J. For. 2015, 60, 8–11. [Google Scholar]
- Adams, G.W.; Kunze, H.A.; McCartney, A.; Millican, S.; Park, Y.S. An industrial perspective on the use of advanced reforestation stock technologies. In Vegetative Propagation Forest Trees; Park, Y.S., Bonga, J.M., Moon, H.-K., Eds.; National Institute of Forest Science: Seoul, Republic of Korea, 2016; pp. 323–334. [Google Scholar]
- Tikkinen, M.; Latvala, T.; Aronen, T. Interest in vegetatively propagated Norway spruce materials—A survey among Finnish forest owners and professionals. Silva Fenn. 2021, 55, 10506. [Google Scholar] [CrossRef]
- Egertsdotter, U.; Ahmad, I.; Clapham, D. Automation and Scale Up of Somatic Embryogenesis for Commercial Plant Production, with Emphasis on Conifers. Front. Plant Sci. 2019, 10, 13. [Google Scholar] [CrossRef]
- Sutton, B.C.; Attree, S.M.; El-Kassaby, Y.A.; Grossnickle, S.C.; Polonenko, D.R. Commercialisation of Somatic Embryogenesis for Plantation Forestry. In Plantation Forest Biotechnology for the 21st Century; Walter, C., Carson, M., Eds.; Research Signpost: Kerala, India, 2004; pp. 275–301. [Google Scholar]
- Grossnickle, S.C.; MacDonald, J.E. Seedling Quality: History, Application, and Plant Attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef]
- Varis, S.; Ahola, S.; Jaakola, L.; Aronen, T. Reliable and practical methods for cryopreservation of embryogenic cultures and cold storage of somatic embryos of Norway spruce. Cryobiology 2017, 76, 8–17. [Google Scholar] [CrossRef]
- Liao, Y.K.; Juan, I.P. Improving the germination of somatic embryos of Picea morrisonicola Hayata: Effects of cold storage and partial drying. J. For. Res. 2015, 20, 114–124. [Google Scholar] [CrossRef]
- Lipavská, H.; Konrádová, H. Somatic embryogenesis in conifers: The role of carbohydrate metabolism. Vitr. Cell. Dev. Biol. Plant 2004, 40, 23–30. [Google Scholar] [CrossRef]
- Välimäki, S.; Teyssier, C.; Tikkinen, M.; Delile, A.; Boizot, N.; Varis, S.; Lelu-Walter, M.-A.; Aronen, T. Norway spruce somatic embryogenesis benefits from proliferation of embryogenic tissues on filter discs and cold storage of cotyledonary embryos. Front. Plant Sci. 2022, 13, 1031686. [Google Scholar] [CrossRef]
- Carlsson, J.; Egertsdotter, U.; Ganeteg, U.; Svennerstam, H. Nitrogen utilization during germination of somatic embryos of Norway spruce: Revealing the importance of supplied glutamine for nitrogen metabolism. Trees Struct. Funct. 2019, 33, 383–394. [Google Scholar] [CrossRef]
- Lamhamedi, M.S.; Chamberland, H.; Tremblay, F.M. Epidermal transpiration, ultrastructural characteristics and net photosynthesis of white spruce somatic seedlings in response to in vitro acclimatization. Physiol. Plant. 2003, 118, 554–561. [Google Scholar] [CrossRef]
- Chandra, S.; Bandopadhyay, R.; Kumar, V.; Chandra, R. Acclimatization of tissue cultured plantlets: From laboratory to land. Biotechnol. Lett. 2010, 32, 1199–1205. [Google Scholar] [CrossRef]
- Landis, T.; Tinus, R.; Barnett, J. The Container Tree Nursery Manual: Seedling Propagation; US Department of Agriculture, Forest Service: Washington, DC, USA, 1998; Volume 6, p. 674. [Google Scholar]
- Harju, A.; Heiska, S.; Julkunen-Tiitto, R.; Venäläinen, M.; Aronen, T. Somatic Embryogenesis of Pinus sylvestris L. from Parent Genotypes with High-and Low Stilbene Content in Their Heartwood. Forests 2022, 13, 557. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Tissue culture of conifer seedlings-20 years on: Viewed through the lens of seedling quality. In National Proceedings: Forest and Conservation Nursery Associations—2010. Proc. RMRS-P-65; Riley, L.E., Haase, D.L., Pinto, J.R., Eds.; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; pp. 139–146. [Google Scholar]
- Haase, D.L. Understanding forest seedling quality: Measurements and interpretation. Tree Plant. Notes 2008, 52, 24–30. [Google Scholar]
- Grossnickle, S.C.; Ivetić, V. Root system development and field establishment: Effect of seedling quality. New For. 2022, 53, 1021–1067. [Google Scholar] [CrossRef]
- McGuinness, B.; Duke, M.; Au, C.K.; Lim, S.H. Measuring radiata pine seedling morphological features using a machine vision system. Comput. Electron. Agric. 2021, 189, 106355. [Google Scholar] [CrossRef]
- Landis, T.D.; Nisley, R.G. The Container Tree Nursery Manual: Seedling Processing, Storage, and Outplanting; US Department of Agriculture, Forest Service: Washington, DC, USA, 2010. [Google Scholar]
- Lamhamedi, M.S.; Chamberland, H.; Bernier, P.Y.; Tremblay, F.M. Clonal variation in morphology, growth, physiology, anatomy and ultrastructure of container-grown white spruce somatic plants. Tree Physiol. 2000, 20, 869–880. [Google Scholar] [CrossRef]
- Kozai, T. Acclimatization of micropropagated plants. Biotechnol. Agric. For. 1991, 17, 127–141. [Google Scholar]
- Carneros, E.; Toribio, M.; Celestino, C. Effect of ABA, the auxin antagonist PCIB and partial desiccation on stone pine somatic embryo maturation. Plant Cell Tissue Organ Cult. 2017, 131, 445–458. [Google Scholar] [CrossRef]
- Humánez, A.; Blasco, M.; Brisa, C.; Segura, J.; Arrillaga, I. Somatic embryogenesis from different tissues of Spanish populations of maritime pine. Plant Cell Tissue Organ Cult. 2012, 111, 373–383. [Google Scholar] [CrossRef]
- Hargreaves, C.L.; Reeves, C.B.; Gough, K.; Menzies, M.I.; Low, C.B.; Mullin, T.J. Overcoming the challenges of family and genotype representation and early cell line proliferation in somatic embryogenesis from control-pollinated seeds of Pinus radiata. N. Z. J. For. Sci. 2011, 41, 97–114. [Google Scholar]
- Litvay, J.; Verma, D.; Johnson, M. Influence of a loblolly pine (Pinus taeda L.) culture medium and its components on growth and somatic embryogenesis of the wild carrot (Daucus carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef]
- Hargreaves, C.L.; Reeves, C.B.; Find, J.I.; Gough, K.; Josekutty, P.; Skudder, D.B.; Van Der Maas, S.A.; Sigley, M.R.; Menzies, M.I.; Low, C.B.; et al. Improving initiation, genotype capture, and family representation in somatic embryogenesis of Pinus radiata by a combination of zygotic embryo maturity, media, and explant preparation. Can. J. For. Res. 2009, 39, 1566–1574. [Google Scholar] [CrossRef]
- Hargreaves, C.L.; Grace, L.J.; Holden, D.G. Nurse culture for efficient recovery of cryopreserved Pinus radiata D. Don embryogenic cell lines. Plant Cell Rep. 2002, 21, 40–45. [Google Scholar]
- Poovaiah, C.; Phillips, L.; Geddes, B.; Reeves, C.; Sorieul, M.; Thorlby, G. Genome editing with CRISPR/Cas9 in Pinus radiata (D. Don). BMC Plant Biol. 2021, 21, 363. [Google Scholar] [CrossRef]
- Boulay, M.P.; Gupta, P.K.; Krogstrup, P.; Durzan, D.J. Development of somatic embryos from cell suspension cultures of Norway spruce (Picea abies Karst.). Plant Cell Rep. 1988, 7, 134–137. [Google Scholar] [CrossRef]
- Walter, C.; Grace, L.J. Somatic embryogenesis and genetic transformation in Pinus radiata. In Protocol for Somatic Embryogenesis in Woody Plants; Mohan Jain, S., Gupta, P., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 11–24. [Google Scholar]
- Latutrie, M.; Aronen, T. Long-term cryopreservation of embryogenic Pinus sylvestris cultures. Scand. J. For. Res. 2013, 28, 103–109. [Google Scholar] [CrossRef]
- Gupta, P.K.; Durzan, D.J. Shoot multiplication from mature trees of Douglas-fir (Pseudotsuga menziesii) and sugar pine (Pinus lambertiana). Plant Cell Rep. 1985, 4, 177–179. [Google Scholar] [CrossRef]
- Becwar, M.R.; Nagmani, R.; Wann, S.R. Initiation of Embryogenic Cultures and Somatic Embryo Development in Loblolly-Pine (Pinus taeda). Can. J. For. Res. Rev. Can. Rech. For. 1990, 20, 810–817. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Shoot Height | Root Length | Root Collar Diameter | |||
|---|---|---|---|---|---|---|
| DF | F Value | DF | F Value | DF | F Value | |
| Treatment | 2 | 153.90 *** | 2 | 38.31 *** | 2 | 25.77 *** |
| Cell line (treatment) | 43 | 9.23 *** | 43 | 6.22 *** | 43 | 8.58 *** |
| Error | 731 | 731 | 731 | |||
| Trial | DEC20 | June21 | |||||
|---|---|---|---|---|---|---|---|
| Treatment | Control | MB 2 w | Total | Control | MB 2 w | LM 2 w | Total |
| Germination, % | 37.0 | 29.6 | 33.3 | 41.1 | 29.5 | 28.5 | 32.9 |
| GS1 Survival, % | 48.1 | 45.4 | 46.8 | 28.6 | 7.0 | 10.2 | 15.3 |
| 2 w shoot, mm | 3.83 (±0.18) | 4.20 (±0.27) | 3.99 (±0.16) | 3.82 (±0.15) | 4.47 (±0.19) | 3.84 (±0.19) | 4.03 (±0.1) |
| 2 w root, mm | 3.80 (±0.36) | 3.92 (±0.33) | 3.85 (±0.24) | 4.18 (±0.27) | 5.68 (±0.54) | 6.51 (±0.7) | 5.28 (±0.28) |
| 2 w total, mm | 7.64 (±0.49) | 8.12 (±0.51) | 7.85 (±0.35) | 8.00 (±0.37) | 10.15 (±0.66) | 10.35 (±0.81) | 9.31 (±0.34) |
| Component Group | Chemical | Modified Version [61] of Litvay Medium [60] Used for Proliferation Stage | Modified Version [61] of Litvay Medium [60] Used for Maturation Stage | Modified Version [63] of Boulay Medium [64] Used for Germination |
|---|---|---|---|---|
| [mg/L] | [mg/L] | [mg/L] | ||
| Macronutrient | KH2PO4 | 170 | 170 | 272 |
| KNO3 | 950 | 950 | 505.5 | |
| NH4NO3 | 825 | 825 | 360 | |
| MgSO4·7H2O | 925 | 925 | 493 | |
| CaCl2·2H2O | 11 | 11 | ||
| Ca(NO3)2·4H2O | 708.5 | |||
| KCl | 149 | |||
| Micronutrient | MnSO4·H2O | 21 | 21 | 4.23 |
| H3BO3 | 31 | 31 | 4.64 | |
| ZnSO4·7H2O | 43 | 43 | 1.44 | |
| KI | 4.15 | 4.15 | 0.083 | |
| Na2MoO4.2H2O | 1.25 | 1.25 | 0.12 | |
| CuSO4·5H2O | 0.5 | 0.5 | 0.25 | |
| CoCl2·6H2O | 1.25 | 1.25 | 0.012 | |
| AlCl3·6H2O | 0.024 | |||
| NiCl2·6H2O | 0.024 | |||
| Chelated iron | FeSO4·7H2O | 30 | 30 | 13.9 |
| Na2EDTA·2H2O | 40 | 40 | 18.625 | |
| Vitamin | Nicotinic acid | 5 | 5 | 0.5 |
| Pyridoxine-HCl | 0.5 | 0.5 | 0.5 | |
| Thiamine | 5 | 5 | 1 | |
| myo-inositol | 100 | 100 | 100 | |
| Amino acids | glycine | 2.0 | 2.0 | |
| glutamine | 450 | 550 | ||
| asparagine | 525 | |||
| arginine | 175 | |||
| citrulline | 19.75 | |||
| ornithine | 19 | |||
| lysine | 13.75 | |||
| alanine | 10 | |||
| proline | 8.75 |
| Component | Media Type | Amount |
|---|---|---|
| Besgro Precision No. 8 Bark (3–6 mm) | Composted pine bark | 30% |
| C.A.N Fines fine screened | Composted pine bark | 35% |
| Coco Fibre precision | Coir | 35% |
| Osmocote Exact 12/14 Protect DCT | Control release | 6 kg/m3 |
| Microplus (TE + Mg + Fe) | Micronutrients | 0.75 kg/m3 |
| Dolomite | Calcium magnesium carbonate | 1 kg/m3 |
| Gypsum fine | Calcium sulfate dihydrate | 2 kg/m3 |
| Hydraflo 2 Granular Wetting Agent | Wetting agent | 0.3 kg/m3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reeves, C.; Tikkinen, M.; Aronen, T.; Krajnakova, J. Application of Cold Storage and Short In Vitro Germination for Somatic Embryos of Pinus radiata and P. sylvestris. Plants 2023, 12, 2095. https://doi.org/10.3390/plants12112095
Reeves C, Tikkinen M, Aronen T, Krajnakova J. Application of Cold Storage and Short In Vitro Germination for Somatic Embryos of Pinus radiata and P. sylvestris. Plants. 2023; 12(11):2095. https://doi.org/10.3390/plants12112095
Chicago/Turabian StyleReeves, Cathie, Mikko Tikkinen, Tuija Aronen, and Jana Krajnakova. 2023. "Application of Cold Storage and Short In Vitro Germination for Somatic Embryos of Pinus radiata and P. sylvestris" Plants 12, no. 11: 2095. https://doi.org/10.3390/plants12112095