
Plants and Small Molecules: An Up-and-Coming Synergy
 ,
,  , ,
, , 
Abstract
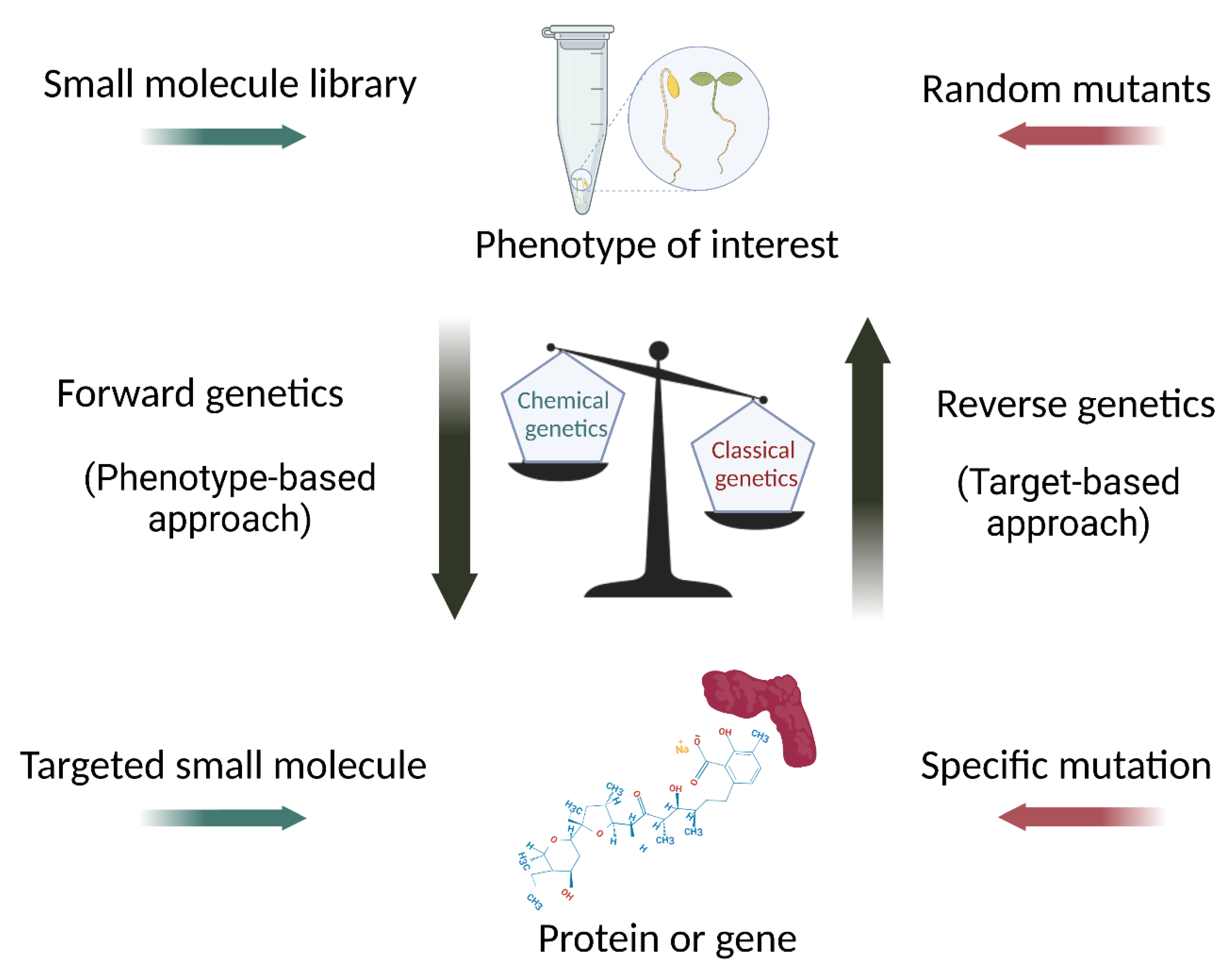
:1. Introduction

2. A Successful Case: Small Molecules to Study Auxin

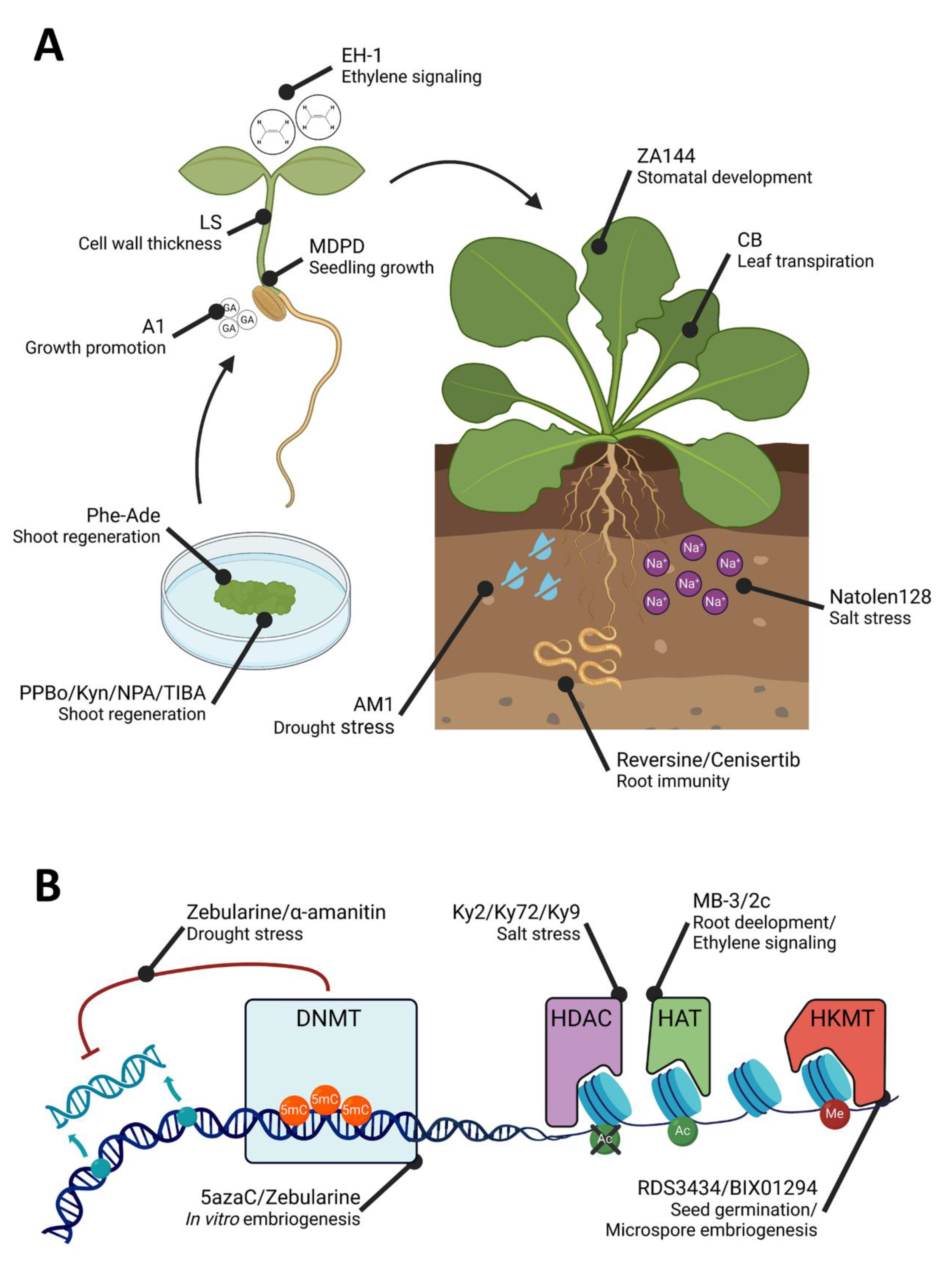
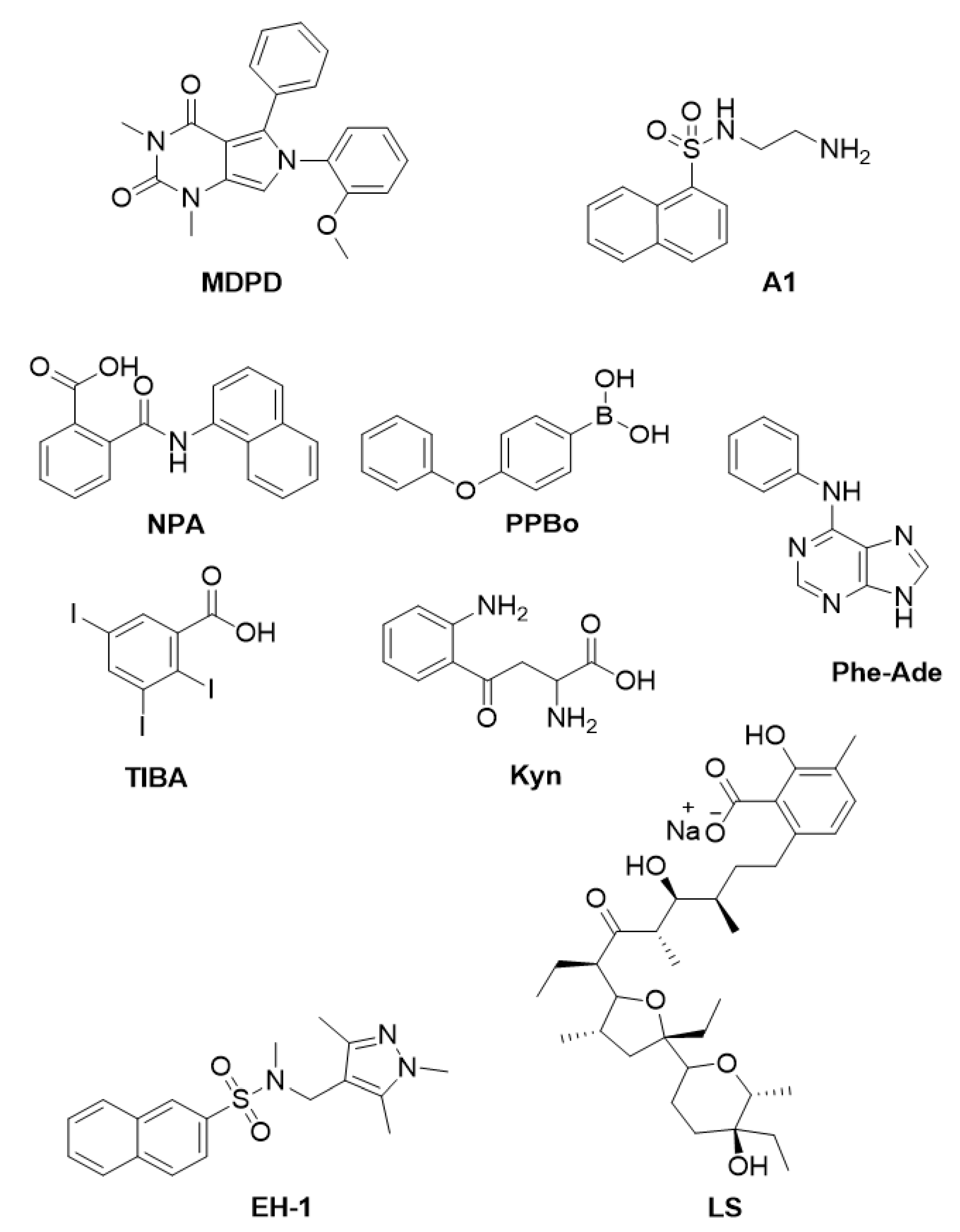
3. Small Molecules to Gain Insights into Plant Morphogenesis and Development

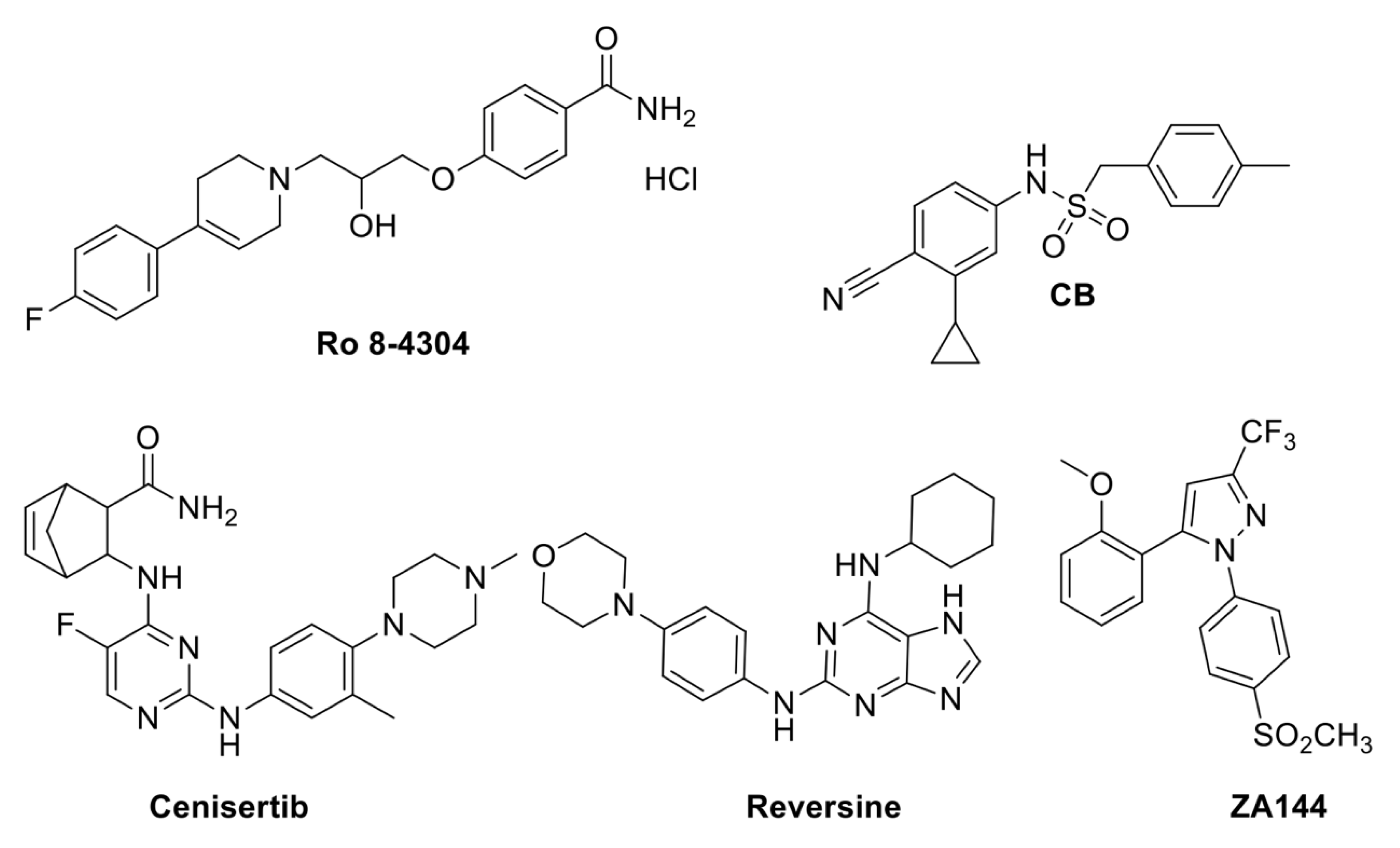
4. Small Molecules to Find Out How Plants Cope with Biotic and Abiotic Stresses

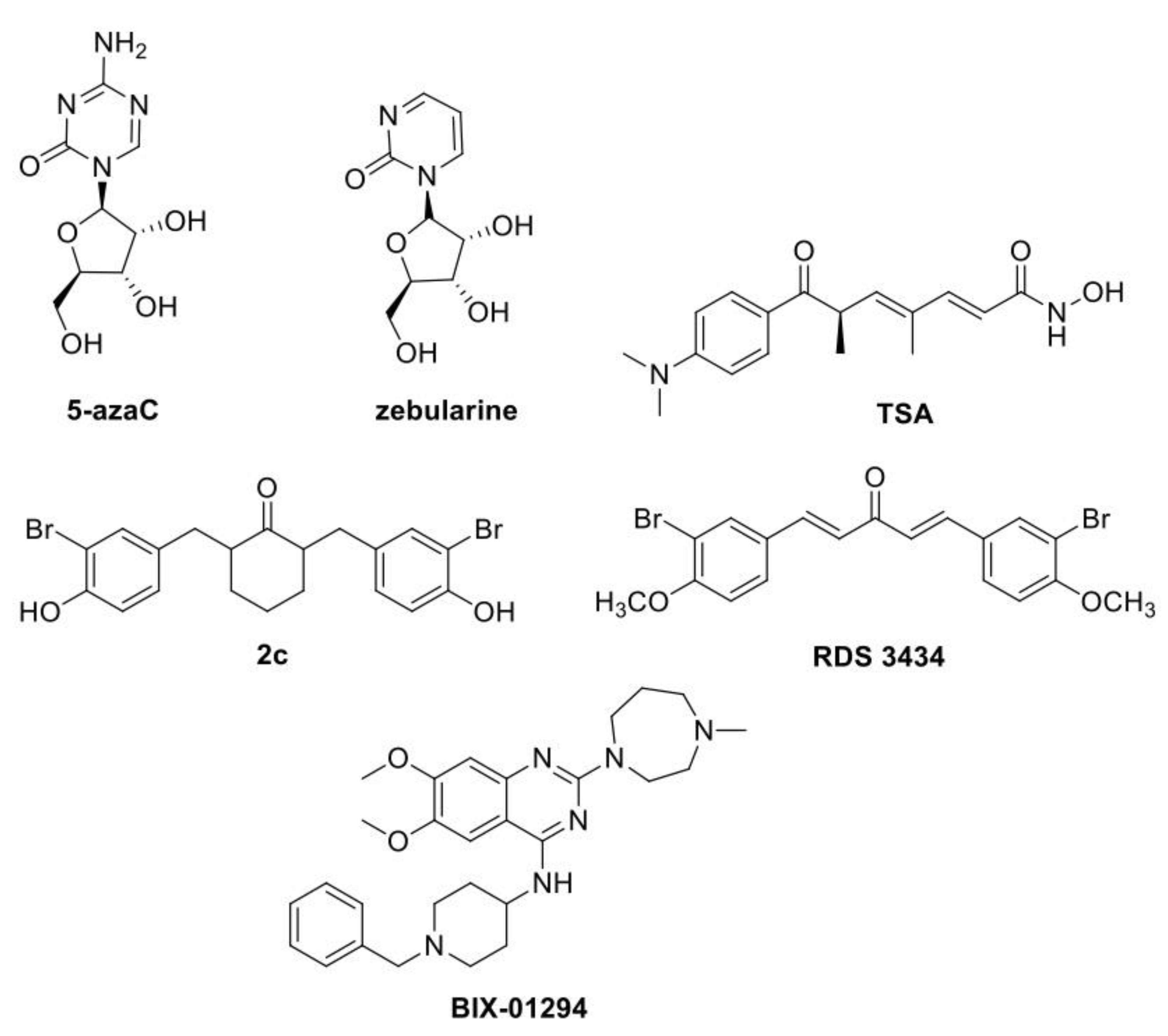
5. Small Molecules to Study Epigenetic-Controlled Processes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
6. Future Perspectives and Challenges
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.; Wang, B.; Duan, C.-G.; Zhu, J.-K. Chemical Probes in Plant Epigenetics Studies. Plant Signal. Behav. 2013, 8, e25364. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Kim, J.-Y.; Chang, Y.T.; Nam, H.G. Forward Chemical Genetic Screening. In Arabidopsis Protocols; Sanchez-Serrano, J.J., Salinas, J., Eds.; Humana Press: Totowa, NJ, USA, 2014; pp. 393–404. ISBN 978-1-62703-580-4. [Google Scholar]
- Kawasumi, M.; Nghiem, P. Chemical Genetics: Elucidating Biological Systems with Small-Molecule Compounds. J. Investig. Dermatol. 2007, 127, 1577–1584. [Google Scholar] [CrossRef]
- Robert, S.; Raikhel, N.V.; Hicks, G.R. Powerful Partners: Arabidopsis and Chemical Genomics. Am. Soc. Plant Biol. 2009, 7, e0109. [Google Scholar] [CrossRef]
- Norambuena, L.; Raikhel, N.V.; Hicks, G.R. Chemical Genomics Approaches in Plant Biology. Methods Mol. Biol. 2009, 553, 345–354. [Google Scholar] [CrossRef]
- Ma, Q.; Robert, S. Auxin Biology Revealed by Small Molecules. Physiol. Plant. 2014, 151, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Grones, P.; Robert, S. Auxin Signaling: A Big Question to be Addressed by Small Molecules. J. Exp. Bot. 2018, 69, 313–328. [Google Scholar] [CrossRef]
- Maher, E.P.; Martindale, S.J. Mutants of Arabidopsis Thaliana with Altered Responses to Auxins and Gravity. Biochem. Genet. 1980, 18, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Christian, M.; Hannah, W.B.; Lüthen, H.; Jones, A.M. Identification of Auxins by a Chemical Genomics Approach. J. Exp. Bot. 2008, 59, 2757–2767. [Google Scholar] [CrossRef]
- Savaldi-Goldstein, S.; Baiga, T.J.; Pojer, F.; Dabi, T.; Butterfield, C.; Parry, G.; Santner, A.; Dharmasiri, N.; Tao, Y.; Estelle, M.; et al. New Auxin Analogs with Growth-Promoting Effects in Intact Plants Reveal a Chemical Strategy to Improve Hormone Delivery. Proc. Natl. Acad. Sci. USA 2008, 105, 15190–15195. [Google Scholar] [CrossRef]
- Tan, X.; Calderon-Villalobos, L.I.A.; Sharon, M.; Zheng, C.; Robinson, C.V.; Estelle, M.; Zheng, N. Mechanism of Auxin Perception by the TIR1 Ubiquitin Ligase. Nature 2007, 446, 640–645. [Google Scholar] [CrossRef]
- Calderon-Villalobos, L.I.; Tan, X.; Zheng, N.; Estelle, M. Auxin Perception—Structural Insights. Cold Spring Harb. Perspect. Biol. 2010, 2, a005546. [Google Scholar] [CrossRef] [PubMed]
- Kleine-Vehn, J.; Friml, J. Polar Targeting and Endocytic Recycling in Auxin-Dependent Plant Development. Annu. Rev. Cell Dev. Biol. 2008, 24, 447–473. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, W.; Friml, J. The March of the PINs: Developmental Plasticity by Dynamic Polar Targeting in Plant Cells. EMBO J. 2010, 29, 2700–2714. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.L.; Casanova, J.E. Turning on ARF: The Sec7 Family of Guanine-Nucleotide-Exchange Factors. Trends Cell Biol. 2000, 10, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Anders, N.; Jürgens, G. Large ARF Guanine Nucleotide Exchange Factors in Membrane Trafficking. Cell. Mol. Life Sci. 2008, 65, 3433–3445. [Google Scholar] [CrossRef] [PubMed]
- Geldner, N.; Friml, J.; Stierhof, Y.D.; Jürgens, G.; Palme, K. Auxin Transport Inhibitors Block PIN1 Cycling and Vesicle Trafficking. Nature 2001, 413, 425–428. [Google Scholar] [CrossRef]
- Singh, M.K.; Jürgens, G. Specificity of Plant Membrane Trafficking—ARFs, Regulators and Coat Proteins. Semin. Cell Dev. Biol. 2018, 80, 85–93. [Google Scholar] [CrossRef]
- Dejonghe, W.; Russinova, E. Plant Chemical Genetics: From Phenotype-Based Screens to Synthetic Biology. Plant Physiol. 2017, 174, 5–20. [Google Scholar] [CrossRef]
- Rivera-Serrano, E.E.; Rodriguez-Welsh, M.F.; Hicks, G.R.; Rojas-Pierce, M. A Small Molecule Inhibitor Partitions Two Distinct Pathways for Trafficking of Tonoplast Intrinsic Proteins in Arabidopsis. PLoS ONE 2012, 7, e44735. [Google Scholar] [CrossRef]
- Dünser, K.; Schöller, M.; Rößling, A.-K.; Löfke, C.; Xiao, N.; Pařízková, B.; Melnik, S.; Rodriguez-Franco, M.; Stöger, E.; Novák, O.; et al. Endocytic Trafficking Promotes Vacuolar Enlargements for Fast Cell Expansion Rates in Plants. Elife 2022, 11, e75945. [Google Scholar] [CrossRef]
- Zouhar, J.; Hicks, G.R.; Raikhel, N.V. Sorting Inhibitors (Sortins): Chemical Compounds to Study Vacuolar Sorting in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 9497–9501. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.M.; Vain, T.; Robert, S. Small Molecules Unravel Complex Interplay between Auxin Biology and Endomembrane Trafficking. J. Exp. Bot. 2015, 66, 4971–4982. [Google Scholar] [CrossRef]
- Rodriguez-Furlán, C.; Rubilar-Hernández, C.; Norambuena, L. Chemical Genomics Translatability from Unicellular to Multicellular Models. Methods Mol. Biol. 2018, 1795, 189–201. [Google Scholar] [CrossRef]
- Ma, Q.; Chang, M.; Drakakaki, G.; Russinova, E. Selective Chemical Probes can Untangle the Complexity of the Plant Cell Endomembrane System. Curr. Opin. Plant Biol. 2022, 68, 102223. [Google Scholar] [CrossRef]
- Arsovski, A.A.; Galstyan, A.; Guseman, J.M.; Nemhauser, J.L. Photomorphogenesis. Am. Soc. Plant Biol. 2012, 10, e0147. [Google Scholar] [CrossRef]
- Chapman, K.D.; Venables, B.; Markovic, R.; RW, B.J.; Bettinger, C. N-Acylethanolamines in Seeds. Quantification of Molecular Species and their Degradation upon Imbibition. Plant Physiol. 1999, 120, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Blancaflor, E.B.; Hou, G.; Chapman, K.D. Elevated Levels of N-Lauroylethanolamine, an Endogenous Constituent of Desiccated Seeds, Disrupt Normal Root Development in Arabidopsis Thaliana Seedlings. Planta 2003, 217, 206–217. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.K.; Cravatt, B.F. Structure and Function of Fatty Acid Amide Hydrolase. Annu. Rev. Biochem. 2005, 74, 411–432. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Shrestha, R.; Kilaru, A.; Wiant, W.; Venables, B.J.; Chapman, K.D.; Blancaflor, E.B. Manipulation of Arabidopsis Fatty Acid Amide Hydrolase Expression Modifies Plant Growth and Sensitivity to N-Acylethanolamines. Proc. Natl. Acad. Sci. USA 2006, 103, 12197–12202. [Google Scholar] [CrossRef]
- Blankman, J.L.; Cravatt, B.F. Chemical Probes of Endocannabinoid Metabolism. Pharmacol. Rev. 2013, 65, 849–871. [Google Scholar] [CrossRef]
- Khan, B.R.; Faure, L.; Chapman, K.D.; Blancaflor, E.B. A Chemical Genetic Screen Uncovers a Small Molecule Enhancer of the N-Acylethanolamine Degrading Enzyme, Fatty Acid Amide Hydrolase, in Arabidopsis. Sci. Rep. 2017, 7, 41121. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Genschik, P. Releasing the Brakes of Plant Growth: How GAs Shutdown DELLA Proteins. J. Exp. Bot. 2009, 60, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Xu, H.; Gao, X.; Fu, X. New Insights into Gibberellin Signaling in Regulating Plant Growth-Metabolic Coordination. Curr. Opin. Plant Biol. 2021, 63, 102074. [Google Scholar] [CrossRef] [PubMed]
- de Lucas, M.; Davière, J.-M.; Rodríguez-Falcón, M.; Pontin, M.; Iglesias-Pedraz, J.M.; Lorrain, S.; Fankhauser, C.; Blázquez, M.A.; Titarenko, E.; Prat, S. A Molecular Framework for Light and Gibberellin Control of Cell Elongation. Nature 2008, 451, 480–484. [Google Scholar] [CrossRef]
- Fu, X.; Richards, D.E.; Fleck, B.; Xie, D.; Burton, N.; Harberd, N.P. The Arabidopsis Mutant Sleepy1gar2-1 Protein Promotes Plant Growth by Increasing the Affinity of the SCFSLY1 E3 Ubiquitin Ligase for DELLA Protein Substrates. Plant Cell 2004, 16, 1406–1418. [Google Scholar] [CrossRef]
- Sun, T. Gibberellin-GID1-DELLA: A Pivotal Regulatory Module for Plant Growth and Development. Plant Physiol. 2010, 154, 567–570. [Google Scholar] [CrossRef]
- Sukiran, N.A.; Pollastri, S.; Steel, P.G.; Knight, M.R. Plant Growth Promotion by the Interaction of a Novel Synthetic Small Molecule with GA-DELLA Function. Plant Direct 2022, 6, e398. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.; Davydov, O.; Knight, H.; Galon, Y.; Knight, M.R.; Fluhr, R.; Fromm, H. Rapid Transcriptome Changes Induced by Cytosolic Ca2+ Transients Reveal ABRE-Related Sequences as Ca2+-Responsive Cis Elements in Arabidopsis. Plant Cell 2006, 18, 2733–2748. [Google Scholar] [CrossRef]
- Feng, S.; Martinez, C.; Gusmaroli, G.; Wang, Y.; Zhou, J.; Wang, F.; Chen, L.; Yu, L.; Iglesias-Pedraz, J.M.; Kircher, S.; et al. Coordinated Regulation of Arabidopsis Thaliana Development by Light and Gibberellins. Nature 2008, 451, 475–479. [Google Scholar] [CrossRef]
- Koornneef, M.; van der Veen, J.H. Induction and Analysis of Gibberellin Sensitive Mutants in Arabidopsis Thaliana (L.) heynh. Theor. Appl. Genet. 1980, 58, 257–263. [Google Scholar] [CrossRef]
- Fridborg, I.; Kuusk, S.; Moritz, T.; Sundberg, E. The Arabidopsis Dwarf Mutant Shi Exhibits Reduced Gibberellin Responses Conferred by Overexpression of a New Putative Zinc Finger Protein. Plant Cell 1999, 11, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Verdeil, J.-L.; Alemanno, L.; Niemenak, N.; Tranbarger, T.J. Pluripotent versus Totipotent Plant Stem Cells: Dependence versus Autonomy? Trends Plant Sci. 2007, 12, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Duclercq, J.; Sangwan-Norreel, B.; Catterou, M.; Sangwan, R.S. De Novo Shoot Organogenesis: From Art to Science. Trends Plant Sci. 2011, 16, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Cary, A.J.; Che, P.; Howell, S.H. Developmental Events and Shoot Apical Meristem Gene Expression Patterns during Shoot Development in Arabidopsis Thaliana. Plant J. 2002, 32, 867–877. [Google Scholar] [CrossRef]
- Pernisová, M.; Klíma, P.; Horák, J.; Válková, M.; Malbeck, J.; Soucek, P.; Reichman, P.; Hoyerová, K.; Dubová, J.; Friml, J.; et al. Cytokinins Modulate Auxin-Induced Organogenesis in Plants via Regulation of the Auxin Efflux. Proc. Natl. Acad. Sci. USA 2009, 106, 3609–3614. [Google Scholar] [CrossRef]
- Gordon, S.P.; Chickarmane, V.S.; Ohno, C.; Meyerowitz, E.M. Multiple Feedback Loops through Cytokinin Signaling Control Stem Cell Number within the Arabidopsis Shoot Meristem. Proc. Natl. Acad. Sci. USA 2009, 106, 16529–16534. [Google Scholar] [CrossRef]
- Motte, H.; Galuszka, P.; Spíchal, L.; Tarkowski, P.; Plíhal, O.; Šmehilová, M.; Jaworek, P.; Vereecke, D.; Werbrouck, S.; Geelen, D. Phenyl-Adenine, Identified in a Light-Dependent Short Hypocotyls4-Assisted Chemical Screen, is a Potent Compound for Shoot Regeneration through the Inhibition of Cytokinin Oxidase/Dehydrogenase Activity. Plant Physiol. 2013, 161, 1229–1241. [Google Scholar] [CrossRef]
- Haseloff, J. GFP Variants for Multispectral Imaging of Living Cells. Methods Cell Biol. 1999, 58, 139–151. [Google Scholar] [CrossRef]
- Miller, C.O. Kinetin and Related Compounds in Plant Growth. Annu. Rev. Plant Physiol. 1961, 12, 395–408. [Google Scholar] [CrossRef]
- Motte, H.; Vereecke, D.; Geelen, D.; Werbrouck, S. The Molecular Path to in vitro Shoot Regeneration. Biotechnol. Adv. 2014, 32, 107–121. [Google Scholar] [CrossRef]
- Ulmasov, T.; Murfett, J.; Hagen, G.; Guilfoyle, T.J. Aux/IAA Proteins Repress Expression of Reporter Genes Containing Natural and Highly Active Synthetic Auxin Response Elements. Plant Cell 1997, 9, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hu, X.; Qin, P.; Prasad, K.; Hu, Y.; Xu, L. The WOX11-LBD16 Pathway Promotes Pluripotency Acquisition in Callus Cells during De Novo Shoot Regeneration in Tissue Culture. Plant Cell Physiol. 2018, 59, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Ohbayashi, I.; Sakamoto, Y.; Kuwae, H.; Kasahara, H.; Sugiyama, M. Enhancement of Shoot Regeneration by Treatment with Inhibitors of Auxin Biosynthesis and Transport during Callus Induction in Tissue Culture of Arabidopsis Thaliana. Plant Biotechnol. 2022, 39, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Abramson, M.; Shoseyov, O.; Shani, Z. Plant Cell Wall Reconstruction toward Improved Lignocellulosic Production and Processability. Plant Sci. 2010, 178, 61–72. [Google Scholar] [CrossRef]
- Okubo-Kurihara, E.; Ohtani, M.; Kurihara, Y.; Kakegawa, K.; Kobayashi, M.; Nagata, N.; Komatsu, T.; Kikuchi, J.; Cutler, S.; Demura, T.; et al. Modification of Plant Cell Wall Structure Accompanied by Enhancement of Saccharification Efficiency Using a Chemical, Lasalocid Sodium. Sci. Rep. 2016, 6, 34602. [Google Scholar] [CrossRef]
- Patel, D.J.; Shen, C. Structural and Kinetic Studies of Lasalocid A (X537A) and its Silver, Sodium, and Barium Salts in Nonpolar Solvents. Proc. Natl. Acad. Sci. USA 1976, 73, 1786–1790. [Google Scholar] [CrossRef]
- Chiang, C.C.; Paul, I.C. Monomeric Forms of the Acid Ionophore Lasalocid A (X-537A) from Polar Solvents. Science 1977, 196, 1441–1443. [Google Scholar] [CrossRef]
- Hamann, T. Plant Cell Wall Integrity Maintenance as an Essential Component of Biotic Stress Response Mechanisms. Front. Plant Sci. 2012, 3, 77. [Google Scholar] [CrossRef]
- Shigeto, J.; Tsutsumi, Y. Diverse Functions and Reactions of Class III Peroxidases. New Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef]
- Denness, L.; McKenna, J.F.; Segonzac, C.; Wormit, A.; Madhou, P.; Bennett, M.; Mansfield, J.; Zipfel, C.; Hamann, T. Cell Wall Damage-Induced Lignin Biosynthesis is Regulated by a Reactive Oxygen Species- and Jasmonic Acid-Dependent Process in Arabidopsis. Plant Physiol. 2011, 156, 1364–1374. [Google Scholar] [CrossRef]
- Goeschl, J.D.; Pratt, H.K.; Bonner, B.A. An Effect of Light on the Production of Ethylene and the Growth of the Plumular Portion of Etiolated Pea Seedlings. Plant Physiol. 1967, 42, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Baur, A.H.; Yang, S.F. Precursors of Ethylene. Plant Physiol. 1969, 44, 1347–1349. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, G.; Mazzeo, A.; Matarrese, A.M.S.; Pacucci, C.; Trani, A.; Fidelibus, M.W.; Gambacorta, G. Ethephon as a Potential Abscission Agent for Table Grapes: Effects on Pre-Harvest Abscission, Fruit Quality, and Residue. Front. Plant Sci. 2016, 7, 620. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Hoshi, T.; Tomio, S.; Ueda, K.; Hara, K. A Chemical Genetics Strategy that Identifies Small Molecules Which Induce the Triple Response in Arabidopsis. Molecules 2017, 22, 2270. [Google Scholar] [CrossRef]
- United Nations Department of Economic and Social Affairs. World Population Prospects 2022: Summary of Results 2022; UN DESA/POP/2022/TR/NO. 3; United Nations Department of Economic and Social Affairs: New York, NY, USA, 2022. [Google Scholar]
- FAO; IFAD; UNICEF. The State of Food Security and Nutrition in the World 2022; FAO: Rome, Italy, 2022; ISBN 978-92-5-136499-4. [Google Scholar]
- Huang, S.; Balgi, A.; Pan, Y.; Li, M.; Zhang, X.; Du, L.; Zhou, M.; Roberge, M.; Li, X. Identification of Methylosome Components as Negative Regulators of Plant Immunity Using Chemical Genetics. Mol. Plant 2016, 9, 1620–1633. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-B.; Li, X.; Cai, J.; Jiang, L.-L.; Zhang, X.; Wu, D.; Wang, L.; Yang, A.; Guo, C.; Chen, J.; et al. A Screening of Inhibitors Targeting the Receptor Kinase FERONIA Reveals Small Molecules that Enhance Plant Root Immunity. Plant Biotechnol. J. 2023, 21, 63–77. [Google Scholar] [CrossRef]
- Bi, D.; Johnson, K.C.M.; Zhu, Z.; Huang, Y.; Chen, F.; Zhang, Y.; Li, X. Mutations in an Atypical TIR-NB-LRR-LIM Resistance Protein Confer Autoimmunity. Front. Plant Sci. 2011, 2, 71. [Google Scholar] [CrossRef]
- Friesen, W.J.; Paushkin, S.; Wyce, A.; Massenet, S.; Pesiridis, G.S.; Van Duyne, G.; Rappsilber, J.; Mann, M.; Dreyfuss, G. The Methylosome, a 20S Complex Containing JBP1 and PICln, Produces Dimethylarginine-Modified Sm Proteins. Mol. Cell. Biol. 2001, 21, 8289–8300. [Google Scholar] [CrossRef]
- Shih, H.-W.; Miller, N.D.; Dai, C.; Spalding, E.P.; Monshausen, G.B. The Receptor-like Kinase FERONIA Is Required for Mechanical Signal Transduction in Arabidopsis Seedlings. Curr. Biol. 2014, 24, 1887–1892. [Google Scholar] [CrossRef]
- Chen, J.; Yu, F.; Liu, Y.; Du, C.; Li, X.; Zhu, S.; Wang, X.; Lan, W.; Rodriguez, P.L.; Liu, X.; et al. FERONIA Interacts with ABI2-Type Phosphatases to Facilitate Signaling Cross-Talk between Abscisic Acid and RALF Peptide in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, E5519-27. [Google Scholar] [CrossRef]
- Masachis, S.; Segorbe, D.; Turrà, D.; Leon-Ruiz, M.; Fürst, U.; El Ghalid, M.; Leonard, G.; López-Berges, M.S.; Richards, T.A.; Felix, G.; et al. A Fungal Pathogen Secretes Plant Alkalinizing Peptides to Increase Infection. Nat. Microbiol. 2016, 1, 16043. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Kita, D.; Peaucelle, A.; Cartwright, H.N.; Doan, V.; Duan, Q.; Liu, M.-C.; Maman, J.; Steinhorst, L.; Schmitz-Thom, I.; et al. The FERONIA Receptor Kinase Maintains Cell-Wall Integrity during Salt Stress through Ca(2+) Signaling. Curr. Biol. 2018, 28, 666–675.e5. [Google Scholar] [CrossRef]
- Guo, H.; Nolan, T.M.; Song, G.; Liu, S.; Xie, Z.; Chen, J.; Schnable, P.S.; Walley, J.W.; Yin, Y. FERONIA Receptor Kinase Contributes to Plant Immunity by Suppressing Jasmonic Acid Signaling in Arabidopsis Thaliana. Curr. Biol. 2018, 28, 3316–3324.e6. [Google Scholar] [CrossRef]
- Dong, Q.; Zhang, Z.; Liu, Y.; Tao, L.-Z.; Liu, H. FERONIA Regulates Auxin-Mediated Lateral Root Development and Primary Root Gravitropism. FEBS Lett. 2019, 593, 97–106. [Google Scholar] [CrossRef]
- Höfte, H. The Yin and Yang of Cell Wall Integrity Control: Brassinosteroid and FERONIA Signaling. Plant Cell Physiol. 2015, 56, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Verstraeten, I.; Roosjen, M.; Takahashi, K.; Rodriguez, L.; Merrin, J.; Chen, J.; Shabala, L.; Smet, W.; Ren, H.; et al. Cell Surface and Intracellular Auxin Signalling for H(+) Fluxes in Root Growth. Nature 2021, 599, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, S.; Zheng, C. Growth or Stress Responses: TMK-FER Balancing Act. Trends Plant Sci. 2023, 28, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Onoda, G.; Steinhardt, P.J.; DiVincenzo, D.; Socolar, J.E. Onoda Etal. Reply. Phys. Rev. Lett. 1989, 62, 1210. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of Protein Phosphatases by Microcystis and Nodularin Associated with Hepatotoxicity. J. Cancer Res. Clin. Oncol. 1990, 116, 609–614. [Google Scholar] [CrossRef]
- Ewart-Toland, A.; Dai, Q.; Gao, Y.-T.; Nagase, H.; Dunlop, M.G.; Farrington, S.M.; Barnetson, R.A.; Anton-Culver, H.; Peel, D.; Ziogas, A.; et al. Aurora-A/STK15 T+91A Is a General Low Penetrance Cancer Susceptibility Gene: A Meta-Analysis of Multiple Cancer Types. Carcinogenesis 2005, 26, 1368–1373. [Google Scholar] [CrossRef]
- Chen, S.; Takanashi, S.; Zhang, Q.; Xiong, W.; Zhu, S.; Peters, E.C.; Ding, S.; Schultz, P.G. Reversine Increases the Plasticity of Lineage-Committed Mammalian Cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10482–10487. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jia, H.; Wang, X.; Shi, C.; Wang, X.; Ma, P.; Wang, J.; Ren, M.; Li, J. Hydrogen Sulfide Positively Regulates Abscisic Acid Signaling through Persulfidation of SnRK2.6 in Guard Cells. Mol. Plant 2020, 13, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Melcher, K.; Ng, L.-M.; Zhou, X.E.; Soon, F.-F.; Xu, Y.; Suino-Powell, K.M.; Park, S.-Y.; Weiner, J.J.; Fujii, H.; Chinnusamy, V.; et al. A Gate-Latch-Lock Mechanism for Hormone Signalling by Abscisic Acid Receptors. Nature 2009, 462, 602–608. [Google Scholar] [CrossRef]
- Miyazono, K.-I.; Miyakawa, T.; Sawano, Y.; Kubota, K.; Kang, H.-J.; Asano, A.; Miyauchi, Y.; Takahashi, M.; Zhi, Y.; Fujita, Y.; et al. Structural Basis of Abscisic Acid Signalling. Nature 2009, 462, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, N.; Hitomi, K.; Arvai, A.S.; Rambo, R.P.; Hitomi, C.; Cutler, S.R.; Schroeder, J.I.; Getzoff, E.D. Structural Mechanism of Abscisic Acid Binding and Signaling by Dimeric PYR1. Science 2009, 326, 1373–1379. [Google Scholar] [CrossRef]
- Santiago, J.; Dupeux, F.; Round, A.; Antoni, R.; Park, S.-Y.; Jamin, M.; Cutler, S.R.; Rodriguez, P.L.; Márquez, J.A. The Abscisic Acid Receptor PYR1 in Complex with Abscisic Acid. Nature 2009, 462, 665–668. [Google Scholar] [CrossRef]
- Yin, P.; Fan, H.; Hao, Q.; Yuan, X.; Wu, D.; Pang, Y.; Yan, C.; Li, W.; Wang, J.; Yan, N. Structural Insights into the Mechanism of Abscisic Acid Signaling by PYL Proteins. Nat. Struct. Mol. Biol. 2009, 16, 1230–1236. [Google Scholar] [CrossRef]
- Cao, M.; Liu, X.; Zhang, Y.; Xue, X.; Zhou, X.E.; Melcher, K.; Gao, P.; Wang, F.; Zeng, L.; Zhao, Y.; et al. An ABA-Mimicking Ligand that Reduces Water Loss and Promotes Drought Resistance in Plants. Cell Res. 2013, 23, 1043–1054. [Google Scholar] [CrossRef]
- Vaidya, A.S.; Peterson, F.C.; Yarmolinsky, D.; Merilo, E.; Verstraeten, I.; Park, S.-Y.; Elzinga, D.; Kaundal, A.; Helander, J.; Lozano-Juste, J.; et al. A Rationally Designed Agonist Defines Subfamily IIIA Abscisic Acid Receptors as Critical Targets for Manipulating Transpiration. ACS Chem. Biol. 2017, 12, 2842–2848. [Google Scholar] [CrossRef]
- Lake, J.A.; Quick, W.P.; Beerling, D.J.; Woodward, F.I. Plant Development. Signals from Mature to New Leaves. Nature 2001, 411, 154. [Google Scholar] [CrossRef]
- Franks, P.J.; Beerling, D.J. Maximum Leaf Conductance Driven by CO2 Effects on Stomatal Size and Density over Geologic Time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of Stomatal Density and Morphology on Water-Use Efficiency in a Changing World. Front. Plant Sci. 2019, 10, 225. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Torii, K.U. Stomatal Development and Perspectives toward Agricultural Improvement. Cold Spring Harb. Perspect. Biol. 2019, 11, 34660. [Google Scholar] [CrossRef]
- De Rybel, B.; Audenaert, D.; Vert, G.; Rozhon, W.; Mayerhofer, J.; Peelman, F.; Coutuer, S.; Denayer, T.; Jansen, L.; Nguyen, L.; et al. Chemical Inhibition of a Subset of Arabidopsis Thaliana GSK3-like Kinases Activates Brassinosteroid Signaling. Chem. Biol. 2009, 16, 594–604. [Google Scholar] [CrossRef]
- Sakai, Y.; Sugano, S.S.; Kawase, T.; Shirakawa, M.; Imai, Y.; Kawamoto, Y.; Sugiyama, H.; Nakagawa, T.; Hara-Nishimura, I.; Shimada, T. The Chemical Compound Bubblin Induces Stomatal Mispatterning in Arabidopsis by Disrupting the Intrinsic Polarity of Stomatal Lineage Cells. Development 2017, 144, 499–506. [Google Scholar] [CrossRef]
- Ziadi, A.; Uchida, N.; Kato, H.; Hisamatsu, R.; Sato, A.; Hagihara, S.; Itami, K.; Torii, K.U. Discovery of Synthetic Small Molecules that Enhance the Number of Stomata: C-H Functionalization Chemistry for Plant Biology. Chem. Commun. 2017, 53, 9632–9635. [Google Scholar] [CrossRef]
- Sako, K.; Van Ha, C.; Matsui, A.; Tanaka, M.; Sato, A.; Seki, M. Transcriptome Analysis of Arabidopsis Thaliana Plants Treated with a New Compound Natolen128, Enhancing Salt Stress Tolerance. Plants 2021, 10, 978. [Google Scholar] [CrossRef]
- Sako, K.; Nguyen, H.M.; Seki, M. Advances in Chemical Priming to Enhance Abiotic Stress Tolerance in Plants. Plant Cell Physiol. 2021, 61, 1995–2003. [Google Scholar] [CrossRef]
- Tiso, M.; Tejero, J.; Kenney, C.; Frizzell, S.; Gladwin, M.T. Nitrite Reductase Activity of Nonsymbiotic Hemoglobins from Arabidopsis Thaliana. Biochemistry 2012, 51, 5285–5292. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A Master Regulator of Salinity Stress Tolerance in Plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef]
- Sako, K.; Futamura, Y.; Shimizu, T.; Matsui, A.; Hirano, H.; Kondoh, Y.; Muroi, M.; Aono, H.; Tanaka, M.; Honda, K.; et al. Inhibition of Mitochondrial Complex I by the Novel Compound FSL0260 Enhances High Salinity-Stress Tolerance in Arabidopsis Thaliana. Sci. Rep. 2020, 10, 8691. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. Reduced DNA Methylation in Arabidopsis Thaliana Results in Abnormal Plant Development. Proc. Natl. Acad. Sci. USA 1996, 93, 8449–8454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and Function of DNA Methylation in Plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-I. A Stochastic Simulation Study on Speciation by Sexual Selection. Evolution 1985, 39, 66–82. [Google Scholar] [CrossRef] [PubMed]
- Pila Quinga, L.A.; Pacheco de Freitas Fraga, H.; do Nascimento Vieira, L.; Guerra, M.P. Epigenetics of Long-Term Somatic Embryogenesis in Theobroma cacao, L.: DNA Methylation and Recovery of Embryogenic Potential. Plant Cell Tissue Organ Cult. 2017, 131, 295–305. [Google Scholar] [CrossRef]
- Zhao, Q.; Du, Y.; Wang, H.; Rogers, H.J.; Yu, C.; Liu, W.; Zhao, M.; Xie, F. 5-Azacytidine Promotes Shoot Regeneration during Agrobacterium-Mediated Soybean Transformation. Plant Physiol. Biochem. PPB 2019, 141, 40–50. [Google Scholar] [CrossRef]
- Nowicka, A.; Juzoń, K.; Krzewska, M.; Dziurka, M.; Dubas, E.; Kopeć, P.; Zieliński, K.; Żur, I. Chemically-Induced DNA de-Methylation Alters the Effectiveness of Microspore Embryogenesis in Triticale. Plant Sci. 2019, 287, 110189. [Google Scholar] [CrossRef]
- Chen, R.; Chen, X.; Huo, W.; Zheng, S.; Lin, Y.; Lai, Z. Transcriptome Analysis of Azacitidine (5-AzaC)-Treatment Affecting the Development of Early Somatic Embryogenesis in Longan. J. Hortic. Sci. Biotechnol. 2021, 96, 311–323. [Google Scholar] [CrossRef]
- De-la-Peña, C.; Nic-Can, G.I.; Galaz-Ávalos, R.M.; Avilez-Montalvo, R.; Loyola-Vargas, V.M. The Role of Chromatin Modifications in Somatic Embryogenesis in Plants. Front. Plant Sci. 2015, 6, 635. [Google Scholar] [CrossRef]
- Solís, M.-T.; El-Tantawy, A.-A.; Cano, V.; Risueño, M.C.; Testillano, P.S. 5-Azacytidine Promotes Microspore Embryogenesis Initiation by Decreasing Global DNA Methylation, but Prevents Subsequent Embryo Development in Rapeseed and Barley. Front. Plant Sci. 2015, 6, 472. [Google Scholar] [CrossRef]
- Nic-Can, G.I.; López-Torres, A.; Barredo-Pool, F.; Wrobel, K.; Loyola-Vargas, V.M.; Rojas-Herrera, R.; De-la-Peña, C. New Insights into Somatic Embryogenesis: Leafy Cotyledon1, Baby Boom1 and WUSCHEL-Related Homeobox4 are Epigenetically Regulated in Coffea Canephora. PLoS ONE 2013, 8, e72160. [Google Scholar] [CrossRef] [PubMed]
- Grzybkowska, D.; Morończyk, J.; Wójcikowska, B.; Gaj, M.D. Azacitidine (5-AzaC)-Treatment and Mutations in DNA Methylase Genes Affect Embryogenic Response and Expression of the Genes that are Involved in Somatic Embryogenesis in Arabidopsis. Plant Growth Regul. 2018, 85, 243–256. [Google Scholar] [CrossRef]
- Chang, S.; Pikaard, C.S. Transcript Profiling in Arabidopsis Reveals Complex Responses to Global Inhibition of DNA Methylation and Histone Deacetylation. J. Biol. Chem. 2005, 280, 796–804. [Google Scholar] [CrossRef] [PubMed]
- McClintock, B. The Origin and Behavior of Mutable Loci in Maize. Proc. Natl. Acad. Sci. USA 1950, 36, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Lisch, D. How Important are Transposons for Plant Evolution? Nat. Rev. Genet. 2013, 14, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Negi, P.; Rai, A.N.; Suprasanna, P. Moving through the Stressed Genome: Emerging Regulatory Roles for Transposons in Plant Stress Response. Front. Plant Sci. 2016, 7, 1448. [Google Scholar] [CrossRef]
- Nishimura, H.; Himi, E.; Eun, C.-H.; Takahashi, H.; Qian, Q.; Tsugane, K.; Maekawa, M. Transgenerational Activation of an Autonomous DNA Transposon, Dart1-24, by 5-AzaC Treatment in Rice. Theor. Appl. Genet. 2019, 132, 3347–3355. [Google Scholar] [CrossRef]
- Boonjing, P.; Masuta, Y.; Nozawa, K.; Kato, A.; Ito, H. The Effect of Zebularine on the Heat-Activated Retrotransposon ONSEN in Arabidopsis Thaliana and Vigna Angularis. Genes Genet. Syst. 2020, 95, 165–172. [Google Scholar] [CrossRef]
- Konečná, K.; Sováková, P.P.; Anteková, K.; Fajkus, J.; Fojtová, M. Distinct Responses of Arabidopsis Telomeres and Transposable Elements to Zebularine Exposure. Int. J. Mol. Sci. 2021, 22, 468. [Google Scholar] [CrossRef]
- Hayashi, Y.; Takehira, K.; Nozawa, K.; Suzuki, T.; Masuta, Y.; Kato, A.; Ito, H. ONSEN Shows Different Transposition Activities in RdDM Pathway Mutants. Genes Genet. Syst. 2020, 95, 183–190. [Google Scholar] [CrossRef]
- Ito, H.; Gaubert, H.; Bucher, E.; Mirouze, M.; Vaillant, I.; Paszkowski, J. An SiRNA Pathway Prevents Transgenerational Retrotransposition in Plants Subjected to Stress. Nature 2011, 472, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Thieme, M.; Lanciano, S.; Balzergue, S.; Daccord, N.; Mirouze, M.; Bucher, E. Inhibition of RNA Polymerase II Allows Controlled Mobilisation of Retrotransposons for Plant Breeding. Genome Biol. 2017, 18, 134. [Google Scholar] [CrossRef] [PubMed]
- Thieme, M.; Brêchet, A.; Bourgeois, Y.; Keller, B.; Bucher, E.; Roulin, A.C. Experimentally Heat-Induced Transposition Increases Drought Tolerance in Arabidopsis Thaliana. New Phytol. 2022, 236, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Bie, X.M.; Dong, L.; Li, X.H.; Wang, H.; Gao, X.-Q.; Li, X.G. Trichostatin A and Sodium Butyrate Promotes Plant Regeneration in Common Wheat. Plant Signal. Behav. 2020, 15, 1820681. [Google Scholar] [CrossRef]
- Castillo, A.M.; Valero-Rubira, I.; Burrell, M.Á.; Allué, S.; Costar, M.A.; Vallés, M.P. Trichostatin A Affects Developmental Reprogramming of Bread Wheat Microspores towards an Embryogenic Route. Plants 2020, 9, 1442. [Google Scholar] [CrossRef]
- Sako, K.; Kim, J.-M.; Matsui, A.; Nakamura, K.; Tanaka, M.; Kobayashi, M.; Saito, K.; Nishino, N.; Kusano, M.; Taji, T.; et al. Ky-2, a Histone Deacetylase Inhibitor, Enhances High-Salinity Stress Tolerance in Arabidopsis Thaliana. Plant Cell Physiol. 2016, 57, 776–783. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Sako, K.; Matsui, A.; Ueda, M.; Tanaka, M.; Ito, A.; Nishino, N.; Yoshida, M.; Seki, M. Transcriptomic Analysis of Arabidopsis Thaliana Plants Treated with the Ky-9 and Ky-72 Histone Deacetylase Inhibitors. Plant Signal. Behav. 2018, 13, e1448333. [Google Scholar] [CrossRef]
- Gehring, C.A.; Irving, H.R. Natriuretic Peptides—A Class of Heterologous Molecules in Plants. Int. J. Biochem. Cell Biol. 2003, 35, 1318–1322. [Google Scholar] [CrossRef]
- Ludidi, N.; Morse, M.; Sayed, M.; Wherrett, T.; Shabala, S.; Gehring, C. A Recombinant Plant Natriuretic Peptide Causes Rapid and Spatially Differentiated K+, Na+ and H+ Flux Changes in Arabidopsis Thaliana Roots. Plant Cell Physiol. 2004, 45, 1093–1098. [Google Scholar] [CrossRef]
- Binzel, M.L.; Hasegawa, P.M.; Handa, A.K.; Bressan, R.A. Adaptation of Tobacco Cells to NaCl. Plant Physiol. 1985, 79, 118–125. [Google Scholar] [CrossRef]
- Biel, M.; Kretsovali, A.; Karatzali, E.; Papamatheakis, J.; Giannis, A. Design, Synthesis, and Biological Evaluation of a Small-Molecule Inhibitor of the Histone Acetyltransferase Gcn5. Angew. Chem. Int. Ed. Engl. 2004, 43, 3974–3976. [Google Scholar] [CrossRef] [PubMed]
- Aquea, F.; Timmermann, T.; Herrera-Vásquez, A. Chemical Inhibition of the Histone Acetyltransferase Activity in Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2017, 483, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Weiste, C.; Dröge-Laser, W. The Arabidopsis Transcription Factor BZIP11 Activates Auxin-Mediated Transcription by Recruiting the Histone Acetylation Machinery. Nat. Commun. 2014, 5, 3883. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.G.; Lee, K.; Jang, K.; Seo, P.J. Circadian Expression Profiles of Chromatin Remodeling Factor Genes in Arabidopsis. J. Plant Res. 2015, 128, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Z.; Liu, D.; Zhang, K.; Li, A.; Mao, L. Squamosa Promoter-Binding Protein-like Transcription Factors: Star Players for Plant Growth and Development. J. Integr. Plant Biol. 2010, 52, 946–951. [Google Scholar] [CrossRef]
- Costi, R.; Di Santo, R.; Artico, M.; Miele, G.; Valentini, P.; Novellino, E.; Cereseto, A. Cinnamoyl Compounds as Simple Molecules that Inhibit P300 Histone Acetyltransferase. J. Med. Chem. 2007, 50, 1973–1977. [Google Scholar] [CrossRef] [PubMed]
- Madia, V.N.; Benedetti, R.; Barreca, M.L.; Ngo, L.; Pescatori, L.; Messore, A.; Pupo, G.; Saccoliti, F.; Valente, S.; Mai, A.; et al. Structure-Activity Relationships on Cinnamoyl Derivatives as Inhibitors of P300 Histone Acetyltransferase. Chemmedchem 2017, 12, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Longo, C.; Lepri, A.; Paciolla, A.; Messore, A.; De Vita, D.; Bonaccorsi di Patti, M.C.; Amadei, M.; Madia, V.N.; Ialongo, D.; Di Santo, R.; et al. New Inhibitors of the Human P300/CBP Acetyltransferase are Selectively Active against the Arabidopsis HAC Proteins. Int. J. Mol. Sci. 2022, 23, 10446. [Google Scholar] [CrossRef]
- Liu, C.; Lu, F.; Cui, X.; Cao, X. Histone Methylation in Higher Plants. Annu. Rev. Plant Biol. 2010, 61, 395–420. [Google Scholar] [CrossRef]
- Pontvianne, F.; Blevins, T.; Pikaard, C.S. Arabidopsis Histone Lysine Methyltransferases. Adv. Bot. Res. 2010, 53, 1–22. [Google Scholar] [CrossRef]
- Ruta, V.; Longo, C.; Boccaccini, A.; Madia, V.N.; Saccoliti, F.; Tudino, V.; Di Santo, R.; Lorrai, R.; Dello Ioio, R.; Sabatini, S.; et al. Inhibition of Polycomb Repressive Complex 2 Activity Reduces Trimethylation of H3K27 and Affects Development in Arabidopsis Seedlings. BMC Plant Biol. 2019, 19, 429. [Google Scholar] [CrossRef] [PubMed]
- Berenguer, E.; Bárány, I.; Solís, M.-T.; Pérez-Pérez, Y.; Risueño, M.C.; Testillano, P.S. Inhibition of Histone H3K9 Methylation by BIX-01294 Promotes Stress-Induced Microspore Totipotency and Enhances Embryogenesis Initiation. Front. Plant Sci. 2017, 8, 1161. [Google Scholar] [CrossRef] [PubMed]
- Köhler, C.; Villar, C.B.R. Programming of Gene Expression by Polycomb Group Proteins. Trends Cell Biol. 2008, 18, 236–243. [Google Scholar] [CrossRef]
- Müller, J.; Verrijzer, P. Biochemical Mechanisms of Gene Regulation by Polycomb Group Protein Complexes. Curr. Opin. Genet. Dev. 2009, 19, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, D.; Roudier, F.; Heese, M.; Andersen, E.D.; Gey, D.; Nowack, M.K.; Goodrich, J.; Renou, J.-P.; Grini, P.E.; Colot, V.; et al. Polycomb Repressive Complex 2 Controls the Embryo-to-Seedling Phase Transition. PLoS Genet. 2011, 7, e1002014. [Google Scholar] [CrossRef]
- de Lucas, M.; Pu, L.; Turco, G.; Gaudinier, A.; Morao, A.K.; Harashima, H.; Kim, D.; Ron, M.; Sugimoto, K.; Roudier, F.; et al. Transcriptional Regulation of Arabidopsis Polycomb Repressive Complex 2 Coordinates Cell-Type Proliferation and Differentiation. Plant Cell 2016, 28, 2616–2631. [Google Scholar] [CrossRef]
- Tachibana, M.; Sugimoto, K.; Nozaki, M.; Ueda, J.; Ohta, T.; Ohki, M.; Fukuda, M.; Takeda, N.; Niida, H.; Kato, H.; et al. G9a Histone Methyltransferase Plays a Dominant Role in Euchromatic Histone H3 Lysine 9 Methylation and is Essential for Early Embryogenesis. Genes Dev. 2002, 16, 1779–1791. [Google Scholar] [CrossRef]
- Kubicek, S.; O’Sullivan, R.J.; August, E.M.; Hickey, E.R.; Zhang, Q.; Teodoro, M.L.; Rea, S.; Mechtler, K.; Kowalski, J.A.; Homon, C.A.; et al. Reversal of H3K9me2 by a Small-Molecule Inhibitor for the G9a Histone Methyltransferase. Mol. Cell 2007, 25, 473–481. [Google Scholar] [CrossRef]
- Shi, Y.; Desponts, C.; Do, J.T.; Hahm, H.S.; Schöler, H.R.; Ding, S. Induction of Pluripotent Stem Cells from Mouse Embryonic Fibroblasts by Oct4 and Klf4 with Small-Molecule Compounds. Cell Stem Cell 2008, 3, 568–574. [Google Scholar] [CrossRef]
- Shi, Y.; Do, J.T.; Desponts, C.; Hahm, H.S.; Schöler, H.R.; Ding, S. A Combined Chemical and Genetic Approach for the Generation of Induced Pluripotent Stem Cells. Cell Stem Cell 2008, 2, 525–528. [Google Scholar] [CrossRef]
- Chen, P.; Yao, J.-F.; Huang, R.-F.; Zheng, F.-F.; Jiang, X.-H.; Chen, X.; Chen, J.; Li, M.; Huang, H.-F.; Jiang, Y.-P.; et al. Effect of BIX-01294 on H3K9me2 Levels and the Imprinted Gene Snrpn in Mouse Embryonic Fibroblast Cells. Biosci. Rep. 2015, 35, e00257. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Wu, S. Reprogramming with Small Molecules Instead of Exogenous Transcription Factors. Stem Cells Int. 2015, 2015, 794632. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, H.; Yao, J.; Qin, G.; Wang, F.; Wang, X.; Luo, A.; Zheng, Q.; Cao, C.; Zhao, J. BIX-01294 Increases Pig Cloning Efficiency by Improving Epigenetic Reprogramming of Somatic Cell Nuclei. Reproduction 2016, 151, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Tóth, R.; van der Hoorn, R.A.L. Emerging Principles in Plant Chemical Genetics. Trends Plant Sci. 2010, 15, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Martinez Molina, D.; Jafari, R.; Ignatushchenko, M.; Seki, T.; Larsson, E.A.; Dan, C.; Sreekumar, L.; Cao, Y.; Nordlund, P. Monitoring Drug Target Engagement in Cells and Tissues Using the Cellular Thermal Shift Assay. Science 2013, 341, 84–87. [Google Scholar] [CrossRef]
- Savitski, M.M.; Reinhard, F.B.M.; Franken, H.; Werner, T.; Savitski, M.F.; Eberhard, D.; Martinez Molina, D.; Jafari, R.; Dovega, R.B.; Klaeger, S.; et al. Tracking Cancer Drugs in Living Cells by Thermal Profiling of the Proteome. Science 2014, 346, 1255784. [Google Scholar] [CrossRef]
- Becher, I.; Andrés-Pons, A.; Romanov, N.; Stein, F.; Schramm, M.; Baudin, F.; Helm, D.; Kurzawa, N.; Mateus, A.; Mackmull, M.-T.; et al. Pervasive Protein Thermal Stability Variation during the Cell Cycle. Cell 2018, 173, 1495–1507.e18. [Google Scholar] [CrossRef]
- Ziegler, S.; Pries, V.; Hedberg, C.; Waldmann, H. Target Identification for Small Bioactive Molecules: Finding the Needle in the Haystack. Angew. Chem. Int. Ed. Engl. 2013, 52, 2744–2792. [Google Scholar] [CrossRef]
- Dejonghe, W.; Russinova, E. Target Identification Strategies in Plant Chemical Biology. Front. Plant Sci. 2014, 5, 352. [Google Scholar] [CrossRef]
- Speers, A.E.; Cravatt, B.F. Profiling Enzyme Activities in vivo Using Click Chemistry Methods. Chem. Biol. 2004, 11, 535–546. [Google Scholar] [CrossRef]
- Kaschani, F.; Gu, C.; van der Hoorn, R.A.L. Activity-Based Protein Profiling of Infected Plants. Methods Mol. Biol. 2012, 835, 47–59. [Google Scholar] [CrossRef]
- Dai, X.; Hayashi, K.; Nozaki, H.; Cheng, Y.; Zhao, Y. Genetic and Chemical Analyses of the Action Mechanisms of Sirtinol in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 3129–3134. [Google Scholar] [CrossRef] [PubMed]
- Sandermann, H.J. Plant Metabolism of Xenobiotics. Trends Biochem. Sci. 1992, 17, 82–84. [Google Scholar] [CrossRef]
- Gachon, C.M.M.; Langlois-Meurinne, M.; Henry, Y.; Saindrenan, P. Transcriptional Co-Regulation of Secondary Metabolism Enzymes in Arabidopsis: Functional and Evolutionary Implications. Plant Mol. Biol. 2005, 58, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Schlaich, N.L. Flavin-Containing Monooxygenases in Plants: Looking beyond Detox. Trends Plant Sci. 2007, 12, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chow, T.F.; Puckrin, R.S.; Alfred, S.E.; Korir, A.K.; Larive, C.K.; Cutler, S.R. Chemical Genetic Interrogation of Natural Variation Uncovers a Molecule that is Glycoactivated. Nat. Chem. Biol. 2007, 3, 716–721. [Google Scholar] [CrossRef]
- Halpin, C. Gene Stacking in Transgenic Plants—The Challenge for 21st Century Plant Biotechnology. Plant Biotechnol. J. 2005, 3, 141–155. [Google Scholar] [CrossRef]
- Song, Z.; Tian, J.; Fu, W.; Li, L.; Lu, L.; Zhou, L.; Shan, Z.; Tang, G.; Shou, H. Screening Chinese Soybean Genotypes for Agrobacterium-Mediated Genetic Transformation Suitability. J. Zhejiang Univ. Sci. B 2013, 14, 289–298. [Google Scholar] [CrossRef]
- Backman, T.W.H.; Cao, Y.; Girke, T. ChemMine Tools: An Online Service for Analyzing and Clustering Small Molecules. Nucleic Acids Res. 2011, 39, W486–W491. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lepri, A.; Longo, C.; Messore, A.; Kazmi, H.; Madia, V.N.; Di Santo, R.; Costi, R.; Vittorioso, P. Plants and Small Molecules: An Up-and-Coming Synergy. Plants 2023, 12, 1729. https://doi.org/10.3390/plants12081729
Lepri A, Longo C, Messore A, Kazmi H, Madia VN, Di Santo R, Costi R, Vittorioso P. Plants and Small Molecules: An Up-and-Coming Synergy. Plants. 2023; 12(8):1729. https://doi.org/10.3390/plants12081729
Chicago/Turabian StyleLepri, A., C. Longo, A. Messore, H. Kazmi, V. N. Madia, R. Di Santo, R. Costi, and P. Vittorioso. 2023. "Plants and Small Molecules: An Up-and-Coming Synergy" Plants 12, no. 8: 1729. https://doi.org/10.3390/plants12081729