
New Growth-Related Features of Wheat Grain Pericarp Revealed by Synchrotron-Based X-ray Micro-Tomography and 3D Reconstruction
 , , and
, , and
Abstract
:1. Introduction
2. Results
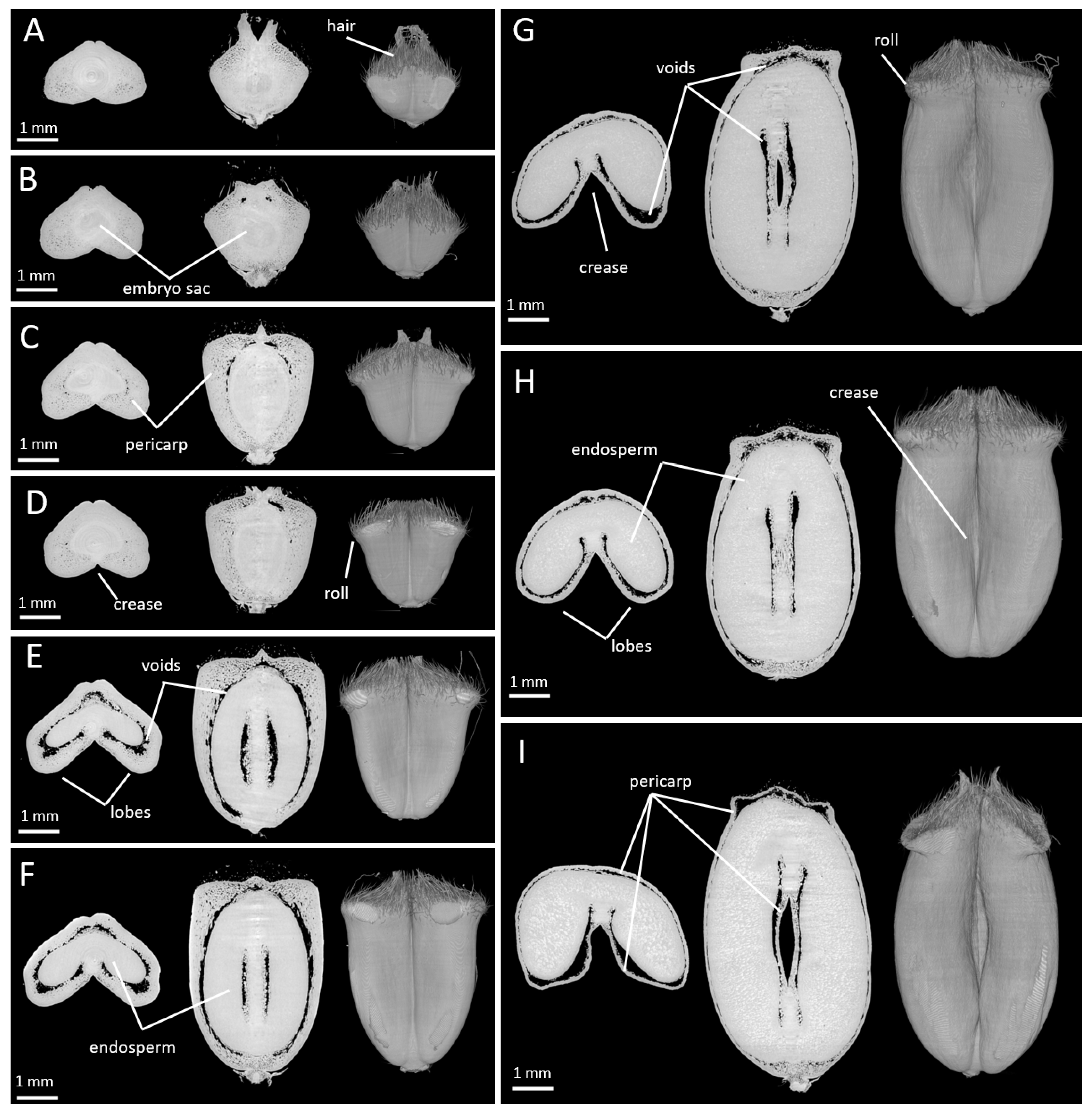
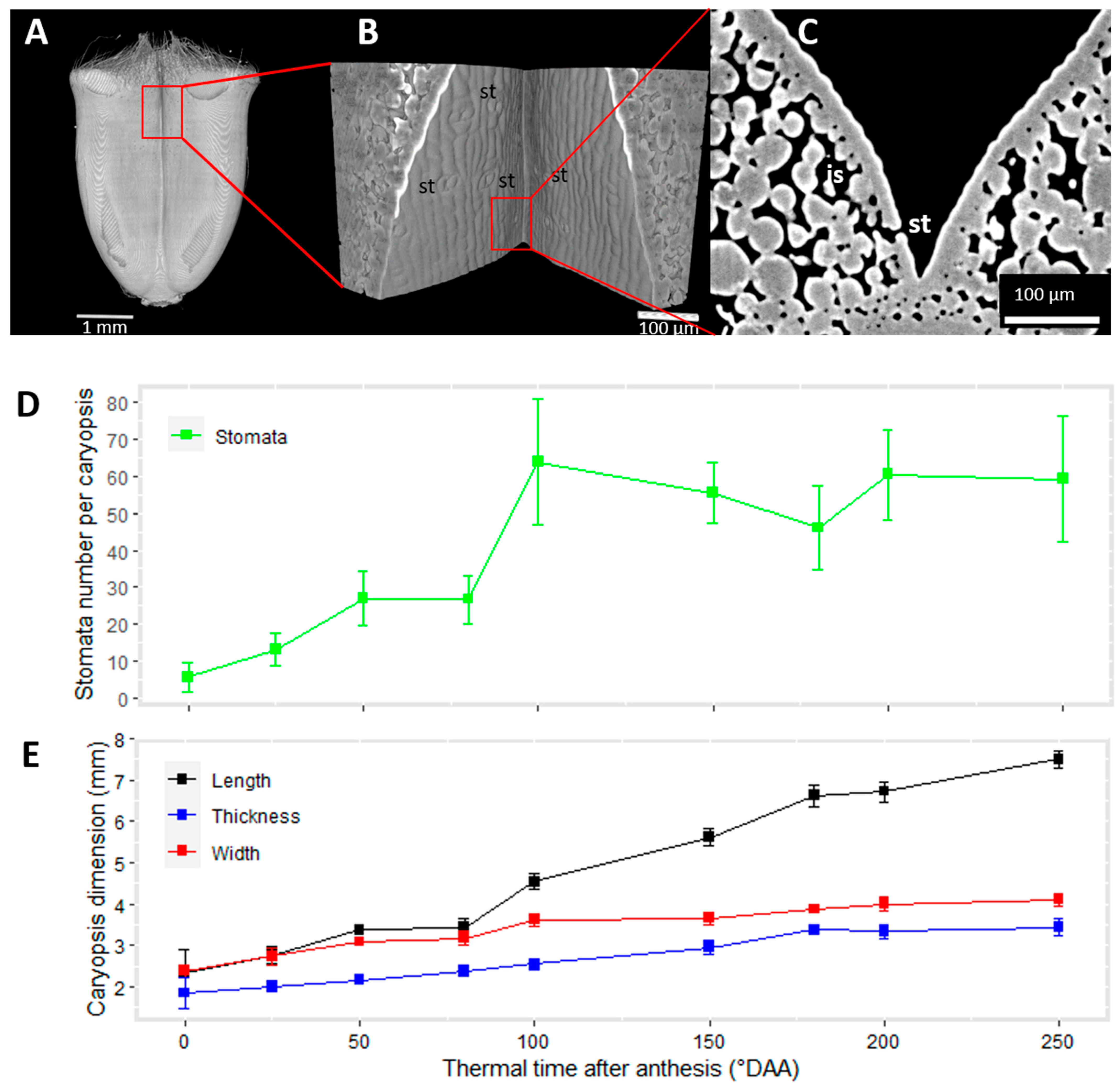
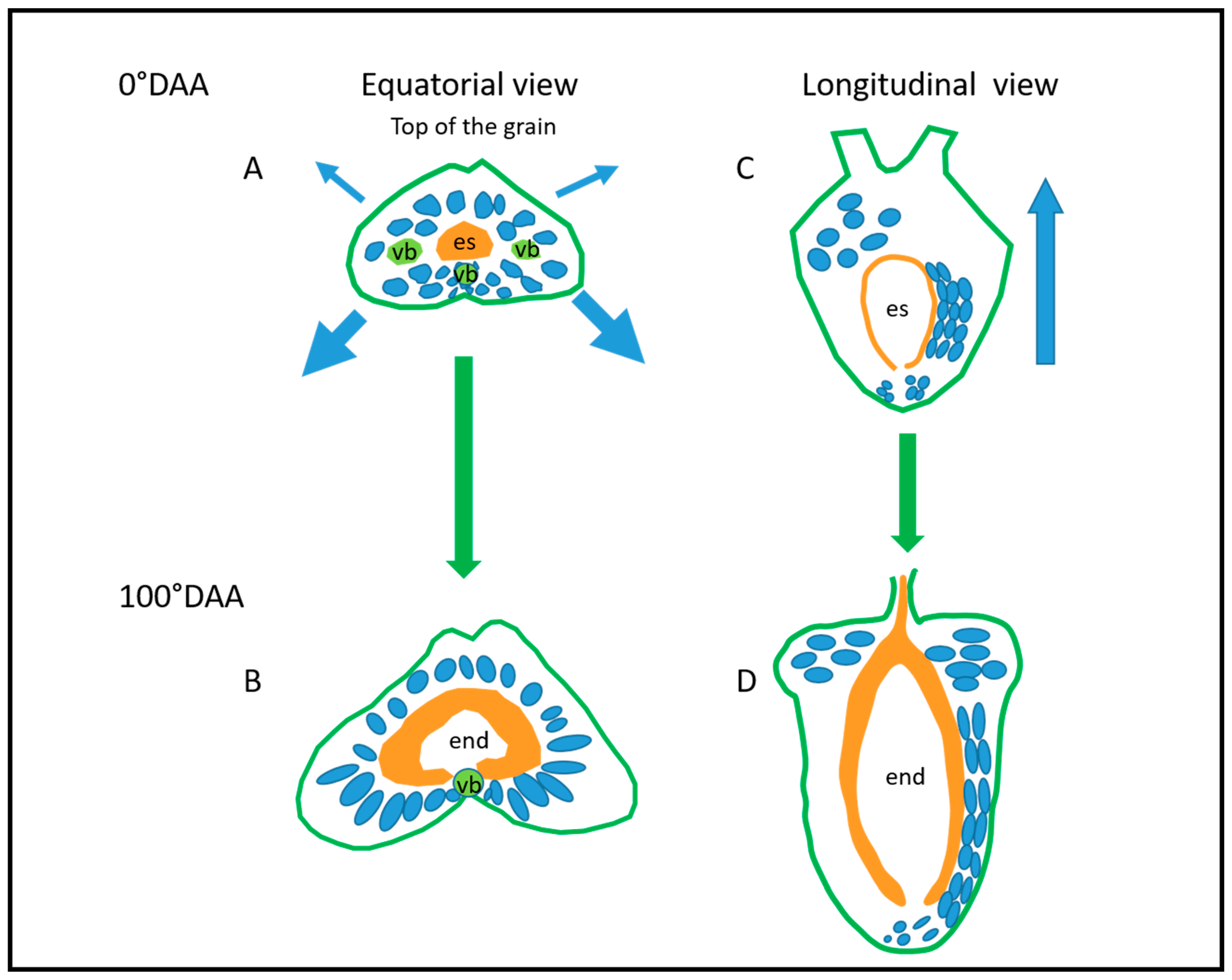
2.1. Changes in Caryopsis Shape with Development
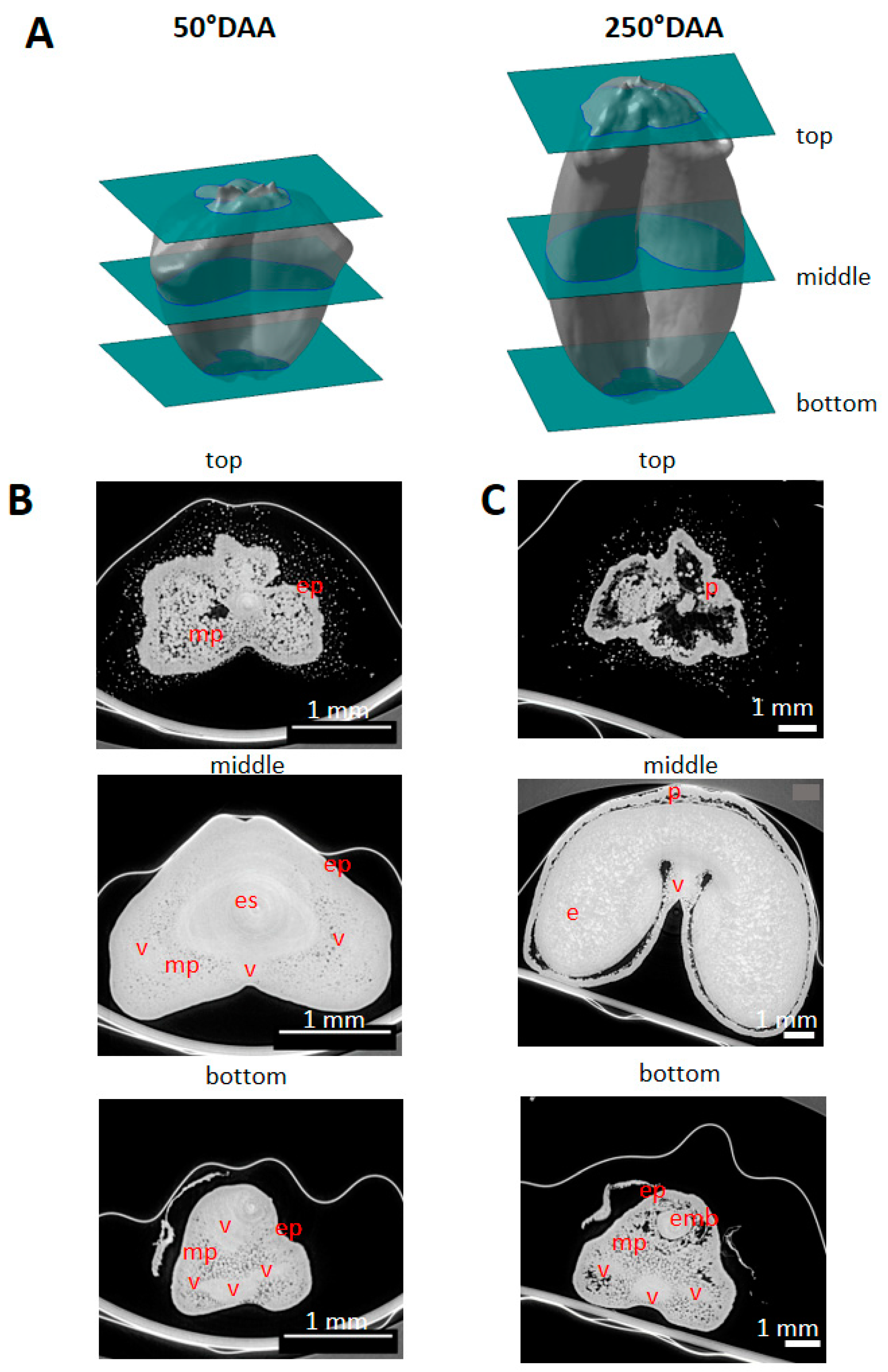
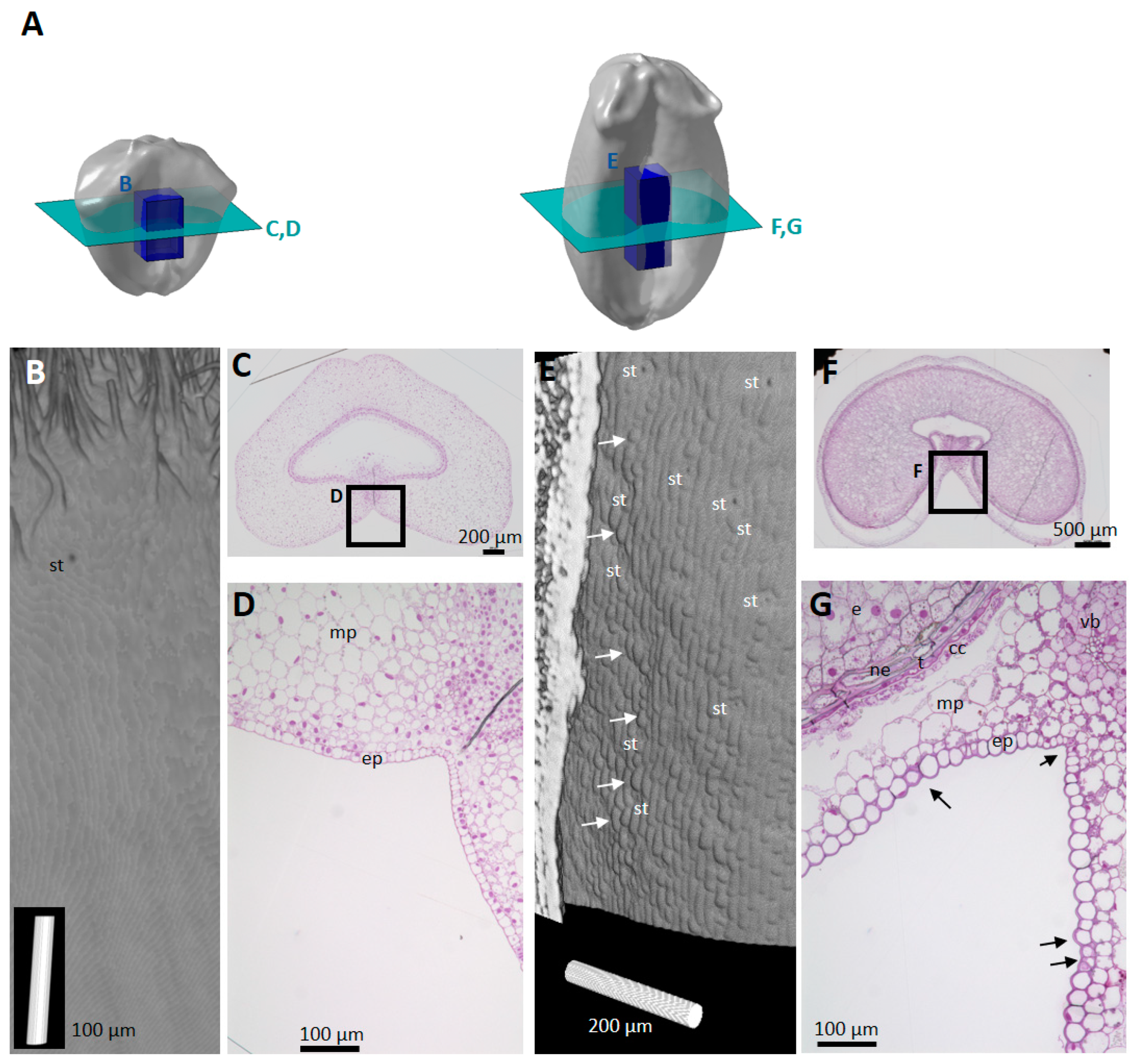
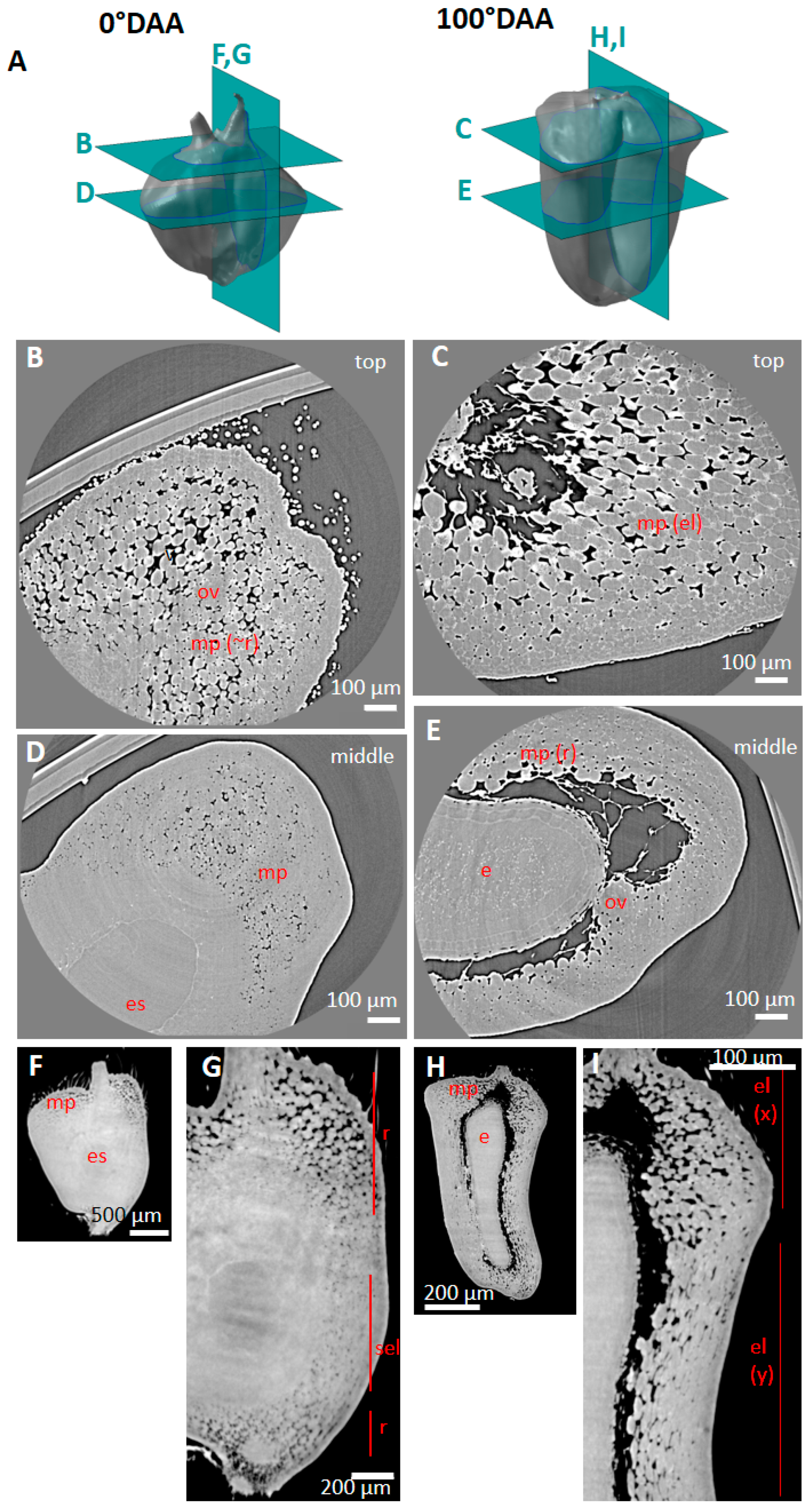
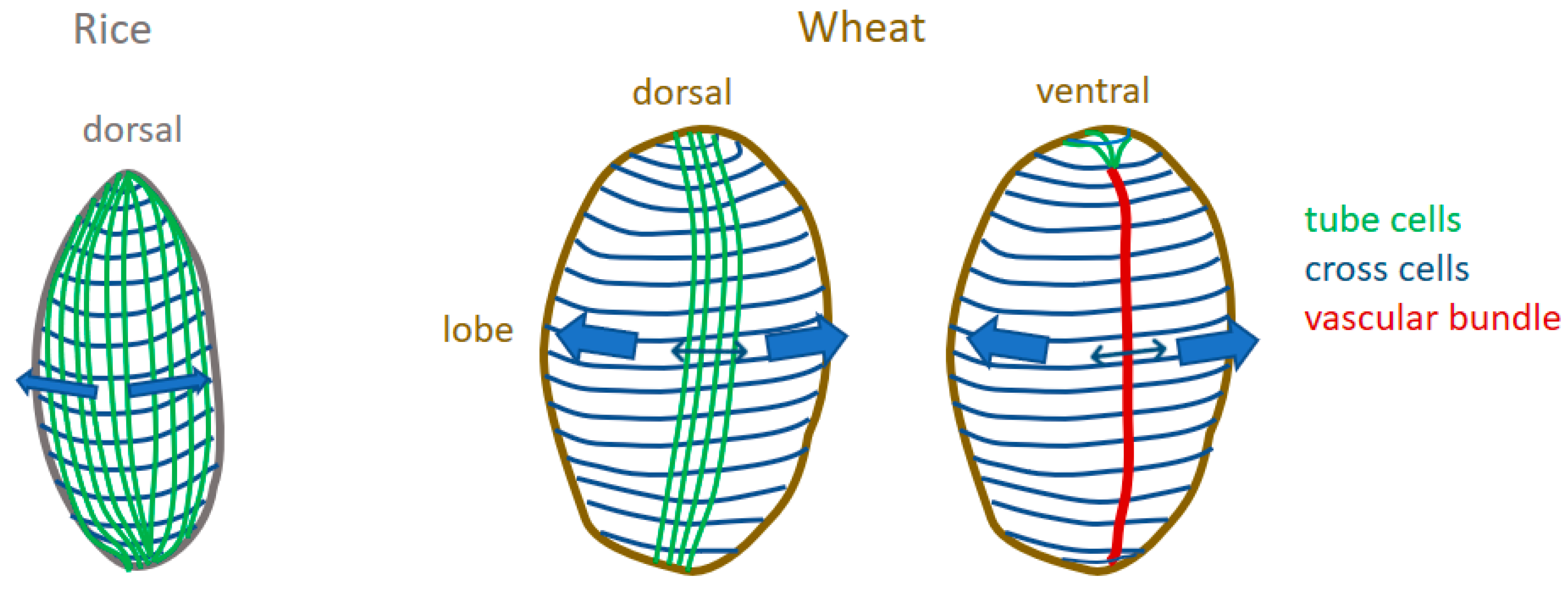
2.2. Spatio-Temporal Variability of Tissues within the Whole Caryopsis
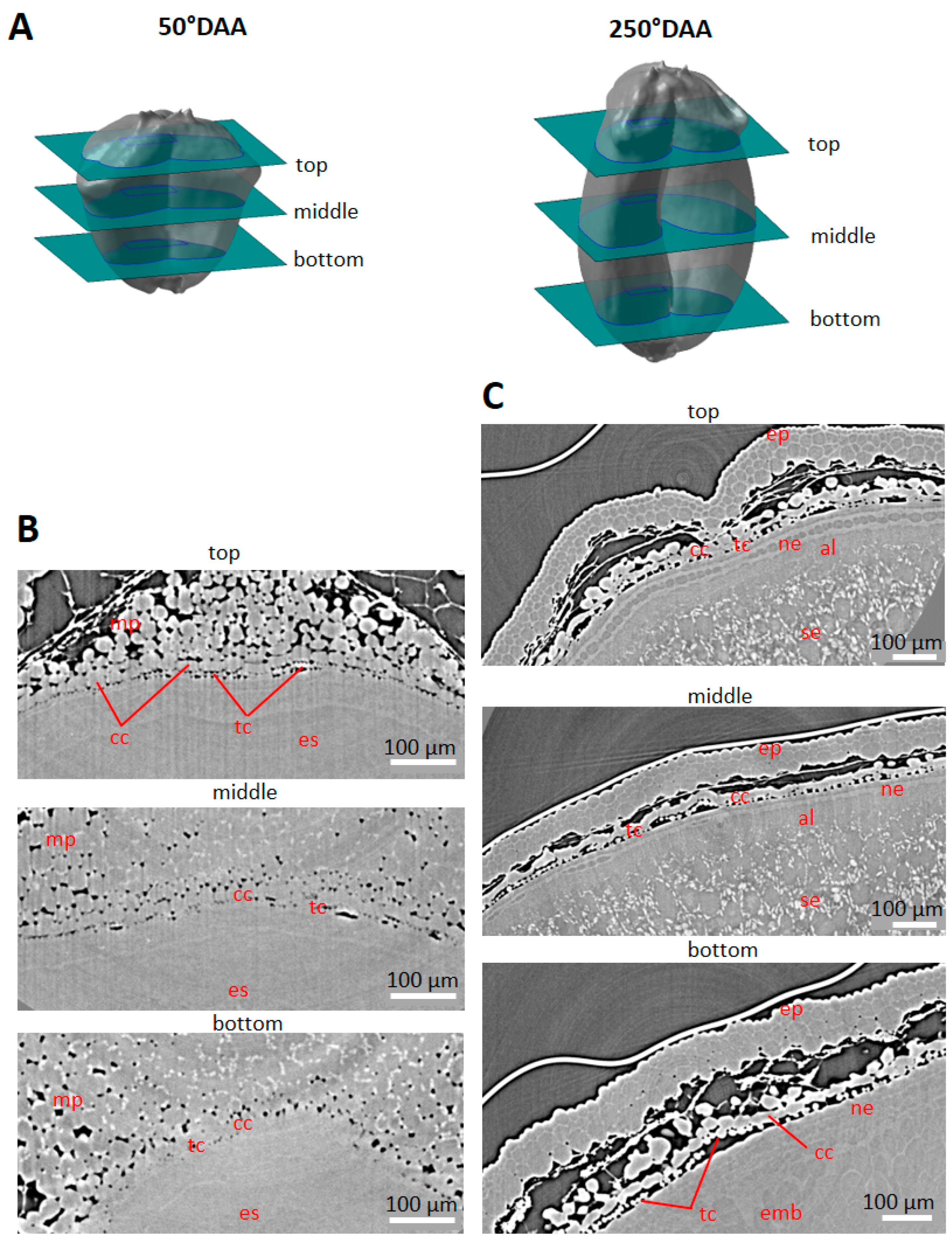
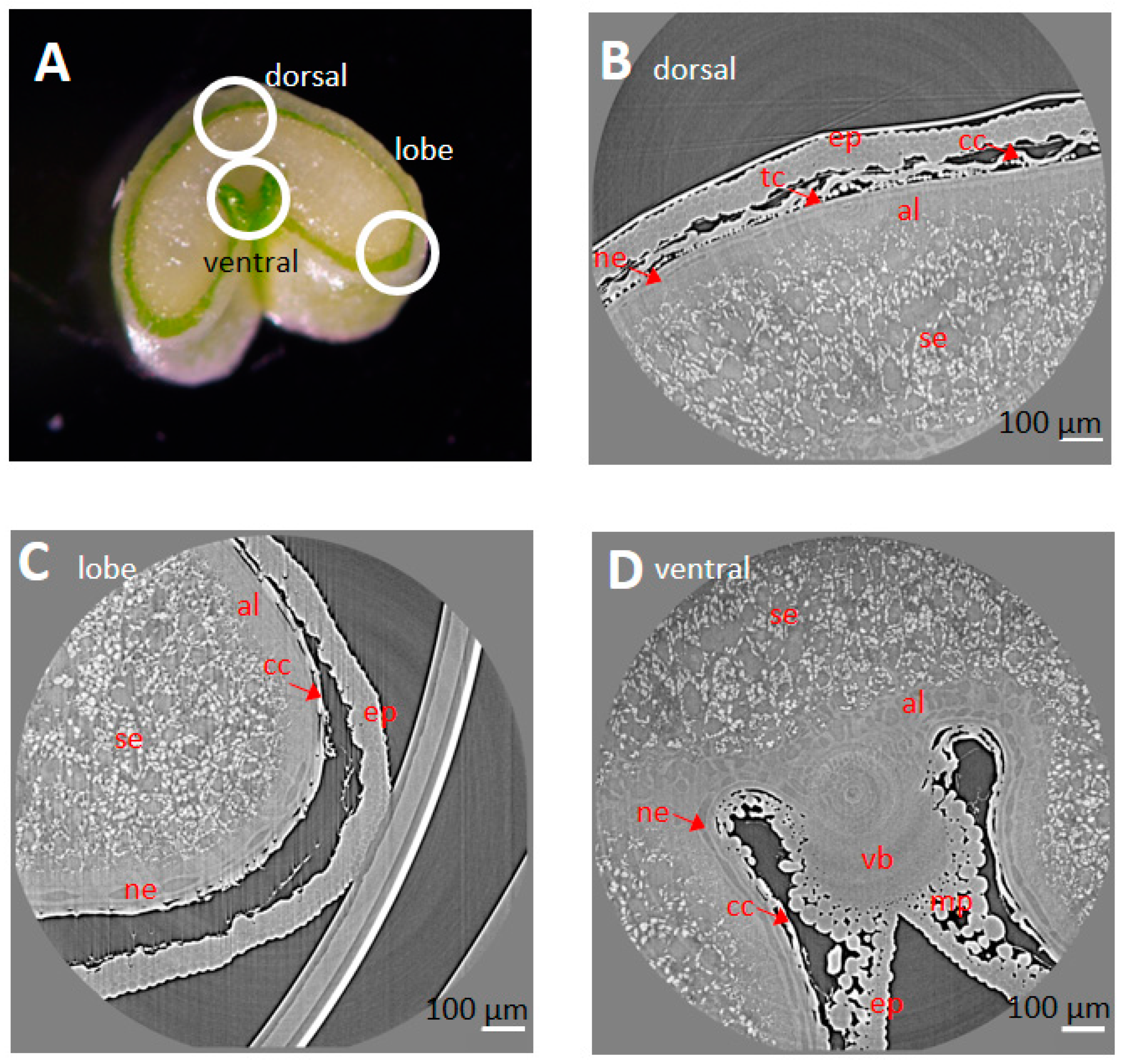
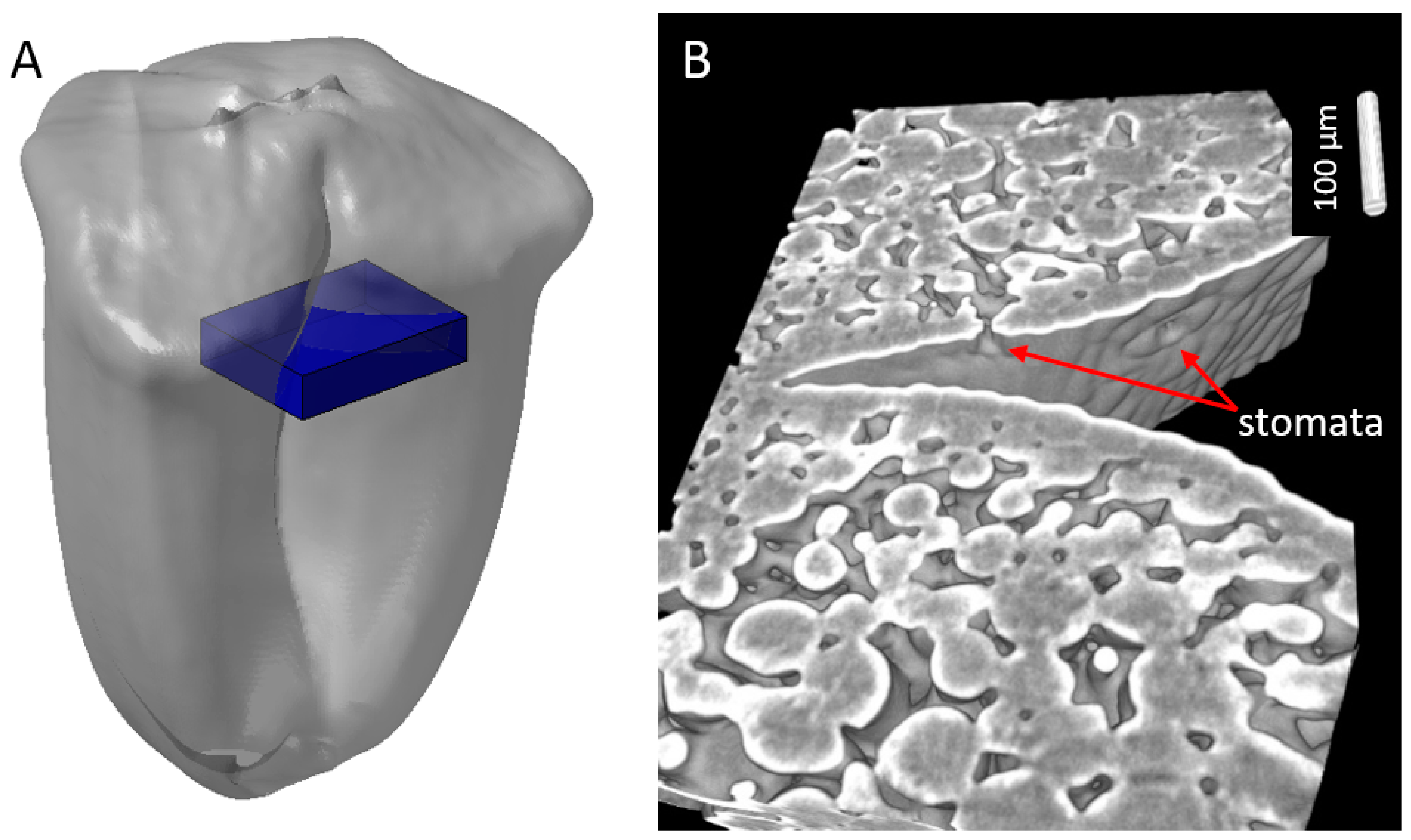
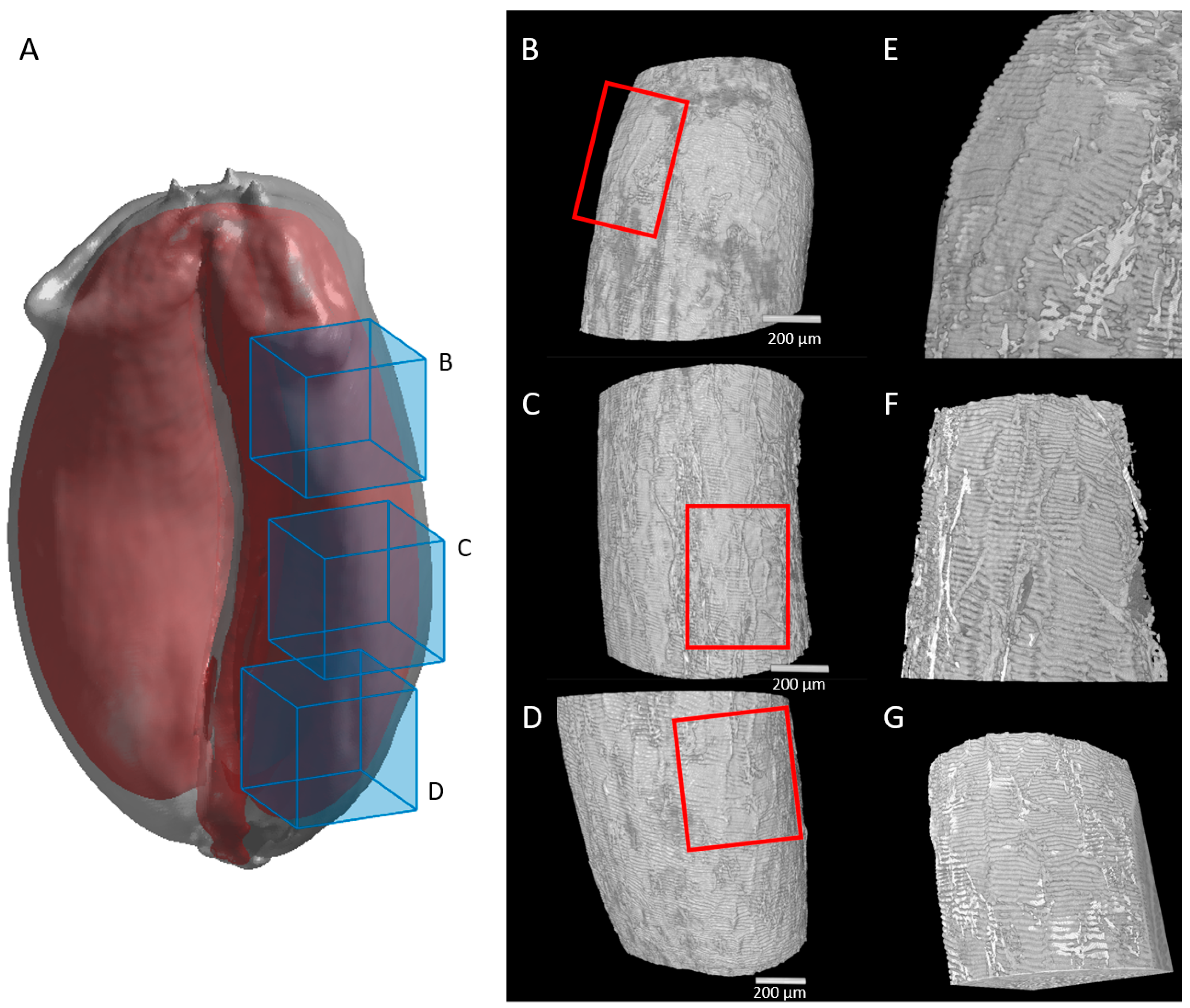
2.3. Cell Morphology within Epicarp
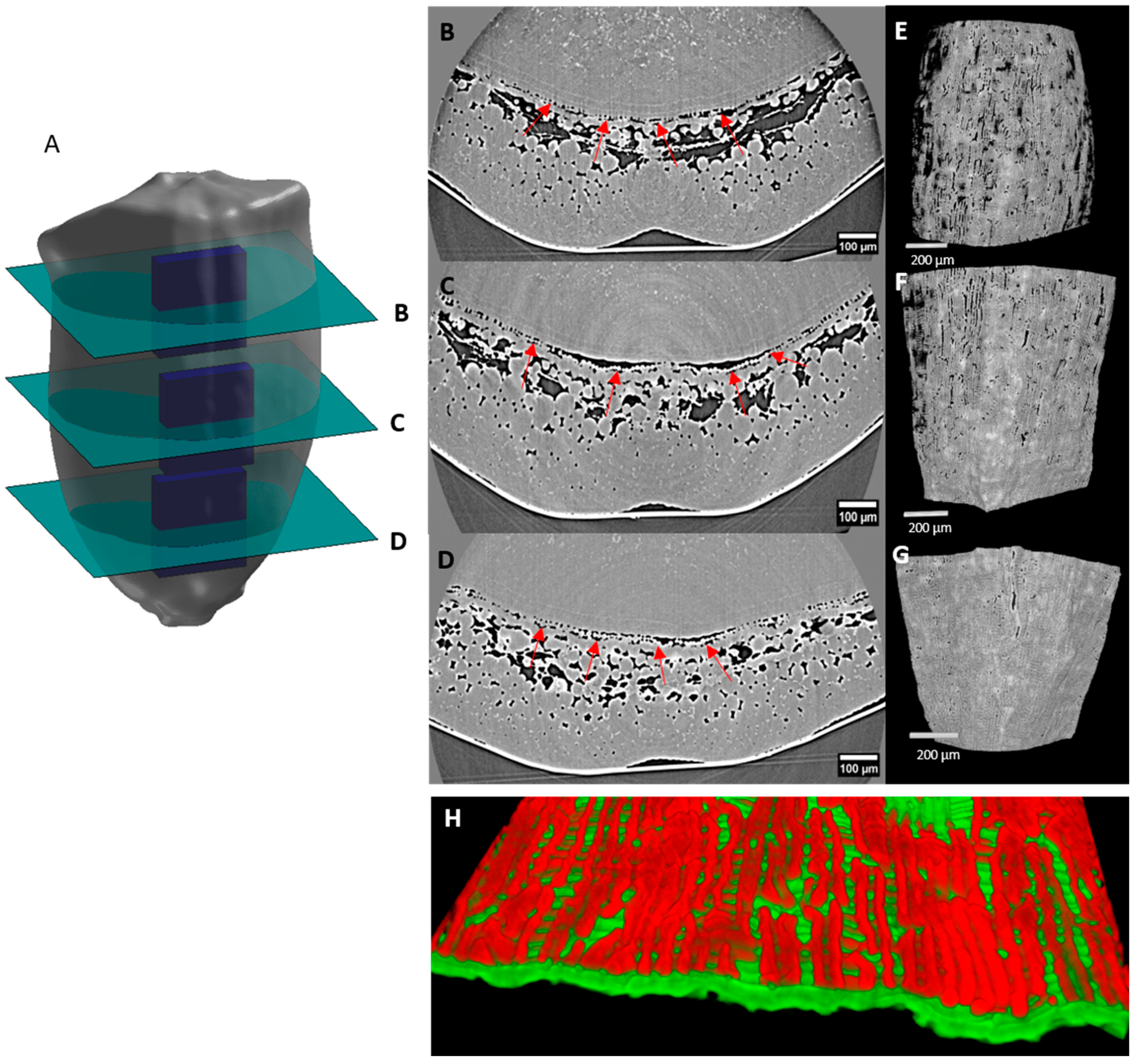
2.4. Porosity and Cell Orientation of Mesocarp
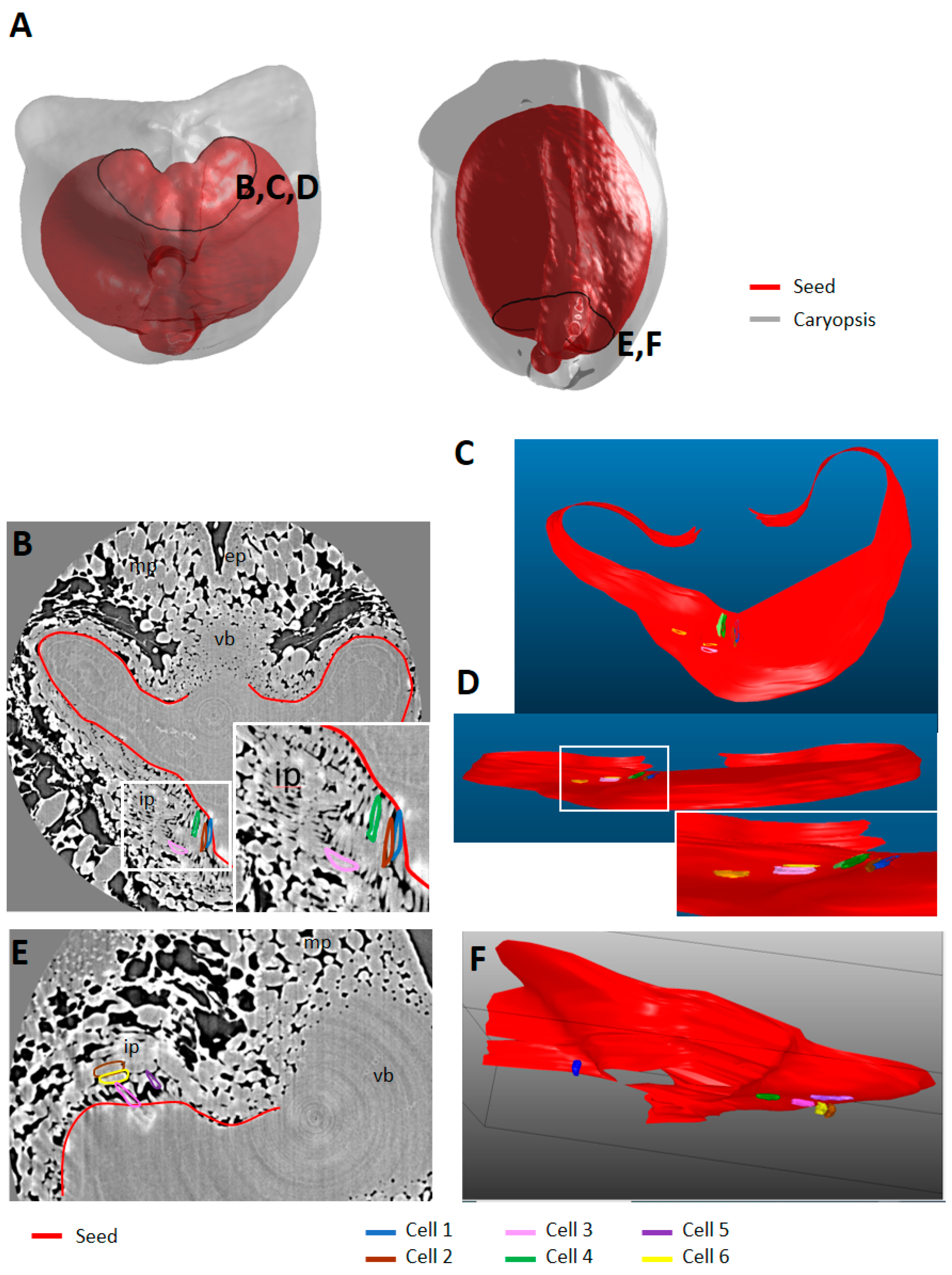
2.5. Cell Morphology and Organisation of Endocarp
3. Discussion
3.1. New 3D Anatomical Features Revealed in Epicarp and Mesocarp
3.2. Caryopsis Elongation, Radial Expansion, and Physical Constraints
3.2.1. Mesocarp Cells and Growth
3.2.2. Putative Role of the Endocarp
3.2.3. Putative Role of the Pericarp in Caryopsis Photosynthesis
3.3. Towards a 3D Model of Caryopsis Morphogenesis
4. Material and Methods
4.1. Plant Material and Preparation of Samples
4.2. Image Processing and Analysis
4.2.1. Image Preprocessing
4.2.2. Caryopsis Segmentation
4.2.3. Alignment
4.2.4. Visualization
4.2.5. Morphometry
4.2.6. Pericarp Porosity
4.2.7. Three-D Visualization of the Organization of the Inner Tissues
4.2.8. Visualization of the 3D Epicarp (RotCrop3D)
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The Contribution of Wheat to Human Diet and Health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Quintero, A.; Molero, G.; Reynolds, M.P.; Calderini, D.F. Trade-off between Grain Weight and Grain Number in Wheat Depends on GxE Interaction: A Case Study of an Elite CIMMYT Panel (CIMCOG). Eur. J. Agron. 2018, 92, 17–29. [Google Scholar] [CrossRef]
- Marshall, D.R.; Mares, D.J.; Moss, H.J.; Ellison, F.W. Effects of Grain Shape and Size on Milling Yields in Wheat. II. Experimental Studies. Aust. J. Agric. Res. 1986, 37, 331–342. [Google Scholar] [CrossRef]
- Evers, T.; Millar, S. Cereal Grain Structure and Development: Some Implications for Quality. J. Cereal Sci. 2002, 36, 261–284. [Google Scholar] [CrossRef]
- Bommert, P.; Werr, W. Gene Expression Patterns in the Maize Caryopsis: Clues to Decisions in Embryo and Endosperm Development. Gene 2001, 271, 131–142. [Google Scholar] [CrossRef]
- Wegel, E.; Pilling, E.; Calder, G.; Drea, S.; Doonan, J.; Dolan, L.; Shaw, P. Three-Dimensional Modelling of Wheat Endosperm Development. New Phytol. 2005, 168, 253–262. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z. Contrast Observation and Investigation of Wheat Endosperm Transfer Cells and Nucellar Projection Transfer Cells. Plant Cell Rep. 2011, 30, 1281–1288. [Google Scholar] [CrossRef]
- Thiel, J. Development of Endosperm Transfer Cells in Barley. Front. Plant Sci. 2014, 5, 108. [Google Scholar] [CrossRef] [Green Version]
- Xiang, D.; Quilichini, T.D.; Liu, Z.; Gao, P.; Pan, Y.; Li, Q.; Nilsen, K.T.; Venglat, P.; Esteban, E.; Pasha, A.; et al. The Transcriptional Landscape of Polyploid Wheats and Their Diploid Ancestors during Embryogenesis and Grain Development. Plant Cell 2019, 31, 2888–2911. [Google Scholar] [CrossRef] [Green Version]
- Lizana, X.C.; Riegel, R.; Gomez, L.D.; Herrera, J.; Isla, A.; McQueen-Mason, S.J.; Calderini, D.F. Expansins Expression Is Associated with Grain Size Dynamics in Wheat (Triticum aestivum L.). J. Exp. Bot. 2010, 61, 1147–1157. [Google Scholar] [CrossRef]
- Le, T.D.Q.; Alvarado, C.; Girousse, C.; Legland, D.; Chateigner-Boutin, A.-L. Use of X-Ray Micro Computed Tomography Imaging to Analyze the Morphology of Wheat Grain through Its Development. Plant Methods 2019, 15, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderini, D.F.; Abeledo, L.G.; Savin, R.; Slafer, G.A. Effect of Temperature and Carpel Size during Pre-Anthesis on Potential Grain Weight in Wheat. J. Agric. Sci. 1999, 132, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Brinton, J.; Simmonds, J.; Minter, F.; Leverington-Waite, M.; Snape, J.; Uauy, C. Increased Pericarp Cell Length Underlies a Major Quantitative Trait Locus for Grain Weight in Hexaploid Wheat. New Phytol. 2017, 215, 1026–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chateigner-Boutin, A.-L.; Lapierre, C.; Alvarado, C.; Yoshinaga, A.; Barron, C.; Bouchet, B.; Bakan, B.; Saulnier, L.; Devaux, M.-F.; Girousse, C.; et al. Ferulate and Lignin Cross-Links Increase in Cell Walls of Wheat Grain Outer Layers during Late Development. Plant Sci. 2018, 276, 199–207. [Google Scholar] [CrossRef]
- Domínguez, F.; Cejudo, F.J. Programmed Cell Death (PCD): An Essential Process of Cereal Seed Development and Germination. Front. Plant Sci. 2014, 5, 366. [Google Scholar] [CrossRef] [Green Version]
- Herrera, J.; Calderini, D.F. Pericarp Growth Dynamics Associate with Final Grain Weight in Wheat under Contrasting Plant Densities and Increased Night Temperature. Ann. Bot. 2020, 126, 1063–1076. [Google Scholar] [CrossRef]
- Xiong, F.; Yu, X.R.; Zhou, L.; Wang, F.; Xiong, A. Structural and Physiological Characterization during Wheat Pericarp Development. Plant Cell Rep. 2013, 32, 1309–1320. [Google Scholar] [CrossRef]
- Xu, A.; Wei, C. Comprehensive Comparison and Applications of Different Sections in Investigating the Microstructure and Histochemistry of Cereal Kernels. Plant Methods 2020, 16, 8. [Google Scholar] [CrossRef]
- Xurun, Y.; Xinyu, C.; Liang, Z.; Jing, Z.; Heng, Y.; Shanshan, S.; Fei, X.; Zhong, W. Structural Development of Wheat Nutrient Transfer Tissues and Their Relationships with Filial Tissues Development. Protoplasma 2015, 252, 605–617. [Google Scholar] [CrossRef]
- Morrison, I.N. The Structure of the Chlorophyll-Containing Cross Cells and Tube Cells of the Inner Pericarp of Wheat During Grain Development. Bot. Gaz. 1976, 137, 85–93. [Google Scholar] [CrossRef]
- Lee, D.R.; Atkey, P.T. Water Loss from the Developing Caryopsis of Wheat (Triticum aestivum). Can. J. Bot. 1984, 62, 1319–1326. [Google Scholar] [CrossRef]
- Evers, A.D.; Bechtel, D.B. Microscopic Structure of the Wheat Grain. In Wheat: Chemistry and Technology; Pomeranz, Y., Ed.; American Association of Cereal Chemists: St. Paul, MN, USA, 1988; Volume 1, pp. 47–95. [Google Scholar]
- Gubatz, S.; Dercksen, V.J.; Brüß, C.; Weschke, W.; Wobus, U. Analysis of Barley (Hordeum vulgare) Grain Development Using Three-Dimensional Digital Models. Plant J. 2007, 52, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Kittelmann, M.; Hawes, C.; Hughes, L. Serial Block Face Scanning Electron Microscopy and the Reconstruction of Plant Cell Membrane Systems. J. Microsc. 2016, 263, 200–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.D.; Kolesik, P.; Høj, P.B.; Coombe, B.G. Confocal Measurement of the Three-Dimensional Size and Shape of Plant Parenchyma Cells in a Developing Fruit Tissue. Plant J. 1999, 19, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Truernit, E.; Bauby, H.; Dubreucq, B.; Grandjean, O.; Runions, J.; Barthélémy, J.; Palauqui, J.-C. High-Resolution Whole-Mount Imaging of Three-Dimensional Tissue Organization and Gene Expression Enables the Study of Phloem Development and Structure in Arabidopsis. Plant Cell 2008, 20, 1494–1503. [Google Scholar] [CrossRef] [Green Version]
- Wuyts, N.; Palauqui, J.-C.; Conejero, G.; Verdeil, J.-L.; Granier, C.; Massonnet, C. High-Contrast Three-Dimensional Imaging of the Arabidopsis Leaf Enables the Analysis of Cell Dimensions in the Epidermis and Mesophyll. Plant Methods 2010, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Stuppy, W.H.; Maisano, J.A.; Colbert, M.W.; Rudall, P.J.; Rowe, T.B. Three-Dimensional Analysis of Plant Structure Using High-Resolution X-Ray Computed Tomography. Trends Plant Sci. 2003, 8, 2–6. [Google Scholar] [CrossRef]
- Rousseau, D.; Widiez, T.; Di Tommaso, S.; Rositi, H.; Adrien, J.; Maire, E.; Langer, M.; Olivier, C.; Peyrin, F.; Rogowsky, P. Fast Virtual Histology Using X-Ray in-Line Phase Tomography: Application to the 3D Anatomy of Maize Developing Seeds. Plant Methods 2015, 11, 55. [Google Scholar] [CrossRef]
- Kunishima, N.; Takeda, Y.; Hirose, R.; Kalasová, D.; Šalplachta, J.; Omote, K. Visualization of Internal 3D Structure of Small Live Seed on Germination by Laboratory-Based X-Ray Microscopy with Phase Contrast Computed Tomography. Plant Methods 2020, 16, 7. [Google Scholar] [CrossRef]
- Legland, D.; Alvarado, C.; Badel, E.; Guillon, F.; King, A.; Le, T.D.Q.; Rivard, C.; Paré, L.; Chateigner-Boutin, A.-L.; Girousse, C. Synchrotron Based X-Ray Microtomography Reveals Cellular Morphological Features of Developing Wheat Grain. Appl. Sci. 2022, 12, 3454. [Google Scholar] [CrossRef]
- Indore, N.S.; Karunakaran, C.; Jayas, D.S. Synchrotron Tomography Applications in Agriculture and Food Sciences Research: A Review. Plant Methods 2022, 18, 101. [Google Scholar] [CrossRef] [PubMed]
- Chaban, I.A.; Gulevich, A.A.; Smirnova, E.A.; Baranova, E.N. Morphological and Ultrastructural Features of Formation of the Skin of Wheat (Triticum aestivum L.) Kernel. Plants 2021, 10, 2538. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, M.; Duffus, C. Morphology and Ultrastructure of Immature Cereal-Grains in Relation to Transport. Ann. Bot. 1979, 44, 67–72. [Google Scholar] [CrossRef]
- Li, W.; Huang, Z.; Han, M.; Ren, Y.; Zhang, X. Development and Structure of Four Different Stamens in Clematis macropetala (Ranunculaceae): Particular Emphasis on Staminodes and Staminal Nectary. Protoplasma 2022, 259, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Waddington, S.R.; Cartwright, P.M.; Wall, P.C. A Quantitative Scale of Spike Initial and Pistil Development in Barley and Wheat. Ann. Bot. 1983, 51, 119–130. [Google Scholar] [CrossRef]
- Bradbury, D.; Macmasters, M.; Cull, I. Structure of the Mature Wheat Kernel. 2. Microscopic Structure of Pericarp, Seed Coat, and Other Coverings of the Endosperm and Germ of Hard Red Winter Wheat. Cereal Chem. 1956, 33, 342–360. [Google Scholar]
- Okada, T.; Jayasinghe, J.E.A.R.M.; Nansamba, M.; Baes, M.; Warner, P.; Kouidri, A.; Correia, D.; Nguyen, V.; Whitford, R.; Baumann, U. Unfertilized Ovary Pushes Wheat Flower Open for Cross-Pollination. J. Exp. Bot. 2018, 69, 399–412. [Google Scholar] [CrossRef]
- Krishnan, S.; Dayanandan, P. Structural and Histochemical Studies on Grain-Filling in the Caryopsis of Rice (Oryza sativa L.). J. Biosci. 2003, 28, 455–469. [Google Scholar] [CrossRef]
- Kutschera, U.; Niklas, K.J. The Epidermal-Growth-Control Theory of Stem Elongation: An Old and a New Perspective. J. Plant Physiol. 2007, 164, 1395–1409. [Google Scholar] [CrossRef]
- Zhou, L.; Du, F.; Feng, S.; Hu, J.; Lu, S.; Long, M.; Jiao, Y. Epidermal Restriction Confers Robustness to Organ Shapes. J. Integr. Plant Biol. 2020, 62, 1853–1867. [Google Scholar] [CrossRef]
- Gegas, V.C.; Nazari, A.; Griffiths, S.; Simmonds, J.; Fish, L.; Orford, S.; Sayers, L.; Doonan, J.H.; Snape, J.W. A Genetic Framework for Grain Size and Shape Variation in Wheat. Plant Cell 2010, 22, 1046–1056. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, M.R.; Mathers, A.; Baillie, A.L.; Dunn, J.; Wilson, M.J.; Hunt, L.; Pajor, R.; Fradera-Soler, M.; Rolfe, S.; Osborne, C.P.; et al. Mesophyll Porosity Is Modulated by the Presence of Functional Stomata. Nat. Commun. 2019, 10, 2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolletschek, H.; Grafahrend-Belau, E.; Munz, E.; Radchuk, V.; Kartäusch, R.; Tschiersch, H.; Melkus, G.; Schreiber, F.; Jakob, P.M.; Borisjuk, L. Metabolic Architecture of the Cereal Grain and Its Relevance to Maximize Carbon Use Efficiency. Plant Physiol. 2015, 169, 1698–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangan, P.; Furtado, A.; Henry, R.J. New Evidence for Grain Specific C 4 Photosynthesis in Wheat. Sci. Rep. 2016, 6, 31721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zitová, B.; Flusser, J. Image Registration Methods: A Survey. Image Vis. Comput. 2003, 21, 977–1000. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, D.; Chéné, Y.; Belin, E.; Semaan, G.; Trigui, G.; Boudehri, K.; Franconi, F.; Chapeau-Blondeau, F. Multiscale Imaging of Plants: Current Approaches and Challenges. Plant Methods 2015, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Legland, D.; Devaux, M.-F.; Guillon, F. Quantitative Imaging of Plants: Multi-Scale Data for Better Plant Anatomy. J. Exp. Bot. 2018, 69, 343–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friston, K.J.; Ashburner, J.; Frith, C.D.; Poline, J.-B.; Heather, J.D.; Frackowiak, R.S.J. Spatial Registration and Normalization of Images. Hum. Brain Mapp. 1995, 3, 165–189. [Google Scholar] [CrossRef]
- Grenander, U.; Miller, M.I. Computational Anatomy: An Emerging Discipline. Q. Appl. Math. 1998, 56, 617–694. [Google Scholar] [CrossRef] [Green Version]
- Young, A.A.; Frangi, A.F. Computational Cardiac Atlases: From Patient to Population and Back. Exp. Physiol. 2009, 94, 578–596. [Google Scholar] [CrossRef] [Green Version]
- Chateigner-Boutin, A.-L.; Bouchet, B.; Alvarado, C.; Bakan, B.; Guillon, F. The Wheat Grain Contains Pectic Domains Exhibiting Specific Spatial and Development-Associated Distribution. PLoS ONE 2014, 9, e89620. [Google Scholar] [CrossRef] [PubMed]
- Chateigner-Boutin, A.-L.; Alvarado, C.; Devaux, M.-F.; Durand, S.; Foucat, L.; Geairon, A.; Grélard, F.; Jamme, F.; Rogniaux, H.; Saulnier, L.; et al. The Endosperm Cavity of Wheat Grains Contains a Highly Hydrated Gel of Arabinoxylan. Plant Sci. 2021, 306, 110845. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, H.; Kedem, Z.M.; Uselton, S.P. Optimal Surface Reconstruction from Planar Contours. Commun. ACM 1977, 20, 693–702. [Google Scholar] [CrossRef]
- Andrey, P.; Maurin, Y. Free-D: An Integrated Environment for Three-Dimensional Reconstruction from Serial Sections. J. Neurosci. Methods 2005, 145, 233–244. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage (°DAA) | 0 | 25 | 50 | 80 | 100 | 150 | 180 | 200 | 250 |
|---|---|---|---|---|---|---|---|---|---|
| Approximate stage (DAA) | 0 | 1.5 | 3 | 5 | 6 | 9 | 11 | 12.5 | 15.5 |
| Low Resolution | 5 | 5 | 5 | 5 | 9 | 10 | 4 | 5 | 5 |
| High Resolution | 2 | 2 | 2 | 2 | 2 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legland, D.; Le, T.D.Q.; Alvarado, C.; Girousse, C.; Chateigner-Boutin, A.-L. New Growth-Related Features of Wheat Grain Pericarp Revealed by Synchrotron-Based X-ray Micro-Tomography and 3D Reconstruction. Plants 2023, 12, 1038. https://doi.org/10.3390/plants12051038
Legland D, Le TDQ, Alvarado C, Girousse C, Chateigner-Boutin A-L. New Growth-Related Features of Wheat Grain Pericarp Revealed by Synchrotron-Based X-ray Micro-Tomography and 3D Reconstruction. Plants. 2023; 12(5):1038. https://doi.org/10.3390/plants12051038
Chicago/Turabian StyleLegland, David, Thang Duong Quoc Le, Camille Alvarado, Christine Girousse, and Anne-Laure Chateigner-Boutin. 2023. "New Growth-Related Features of Wheat Grain Pericarp Revealed by Synchrotron-Based X-ray Micro-Tomography and 3D Reconstruction" Plants 12, no. 5: 1038. https://doi.org/10.3390/plants12051038