
Labellum Features and Chemical Composition of Floral Scent in Bulbophyllum carunculatum Garay, Hamer & Siegrist (Section Lepidorhiza Schltr., Bulbophyllinae Schltr., Orchidaceae Juss.)


Abstract
:1. Introduction
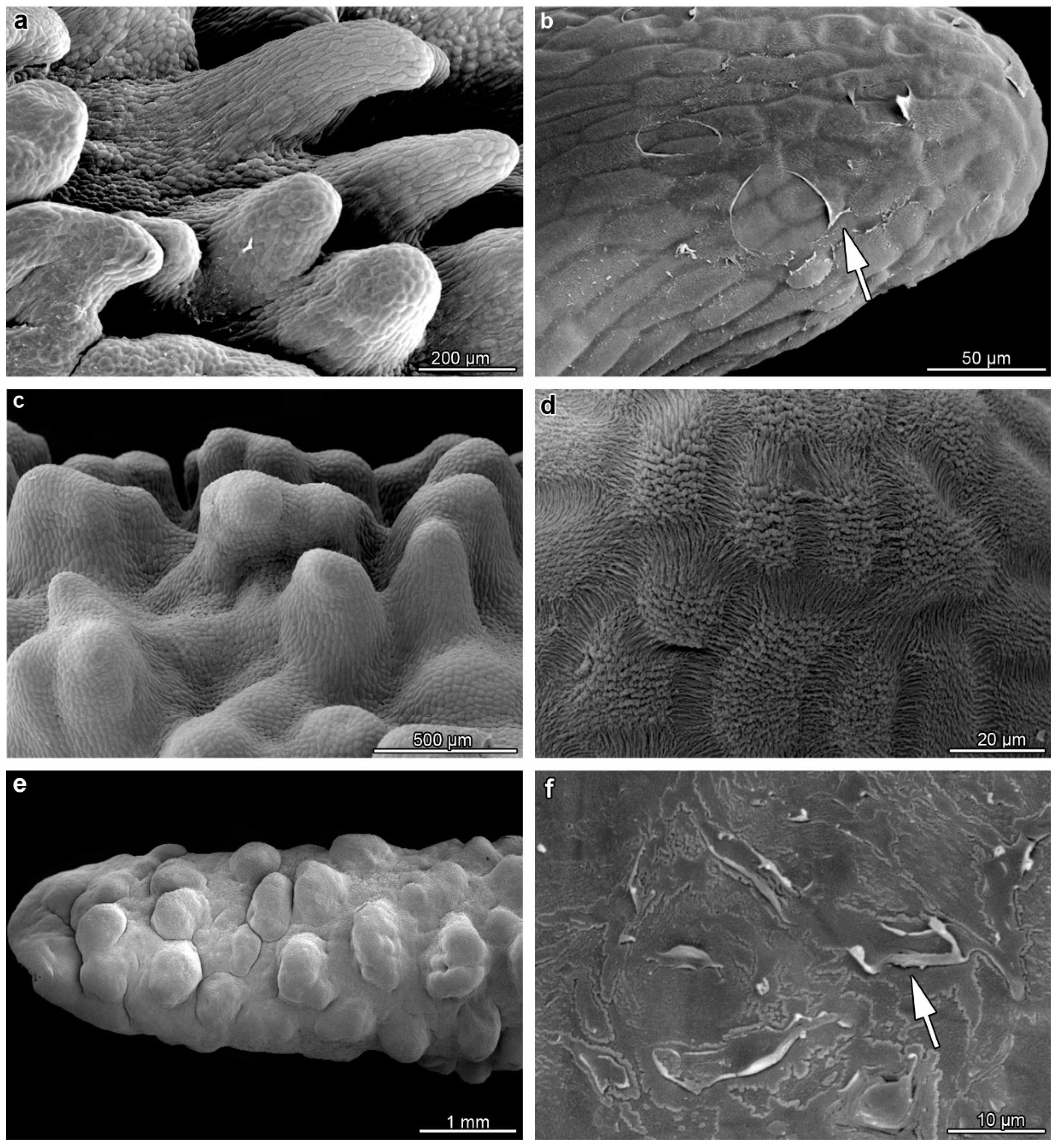
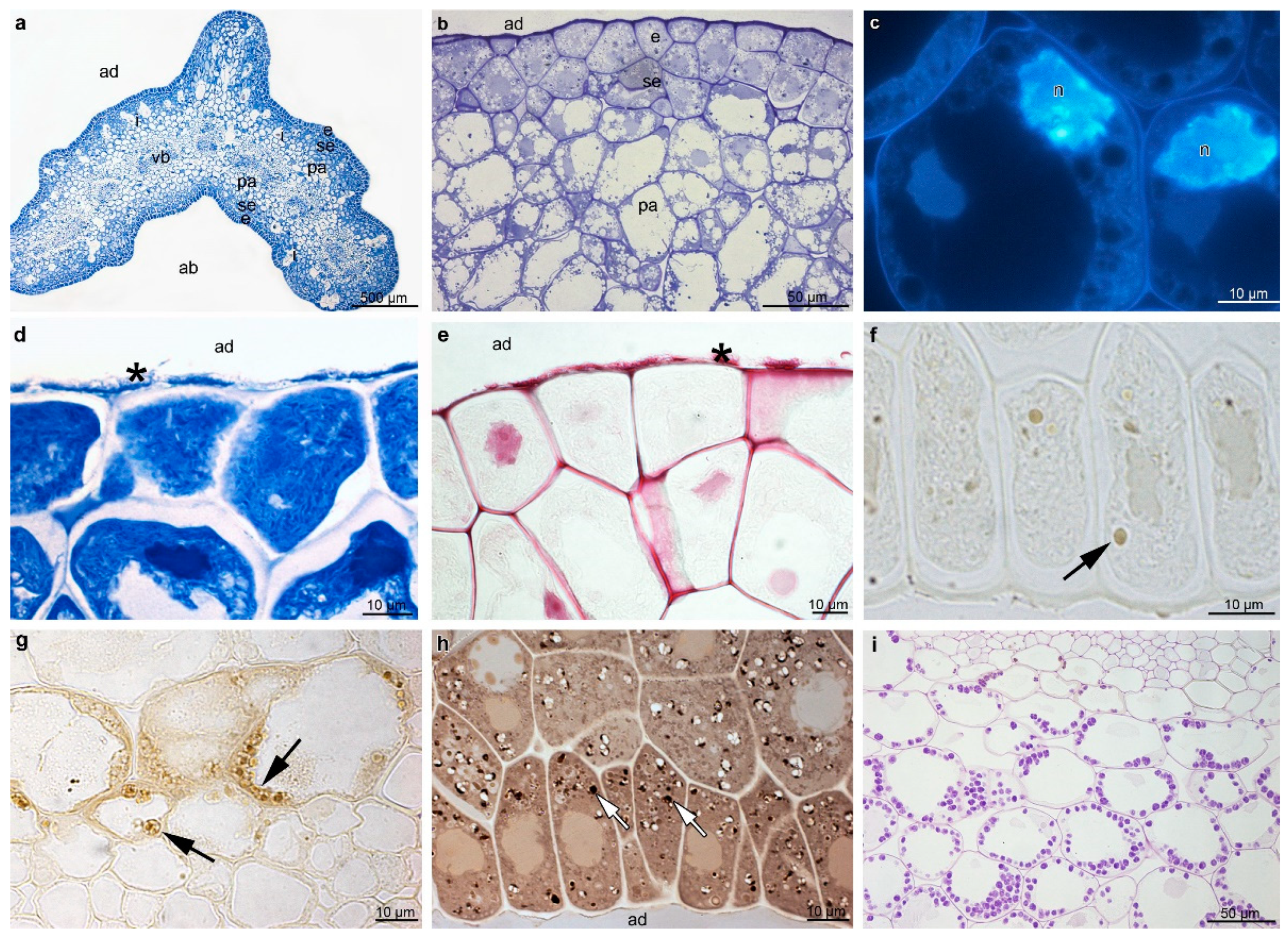
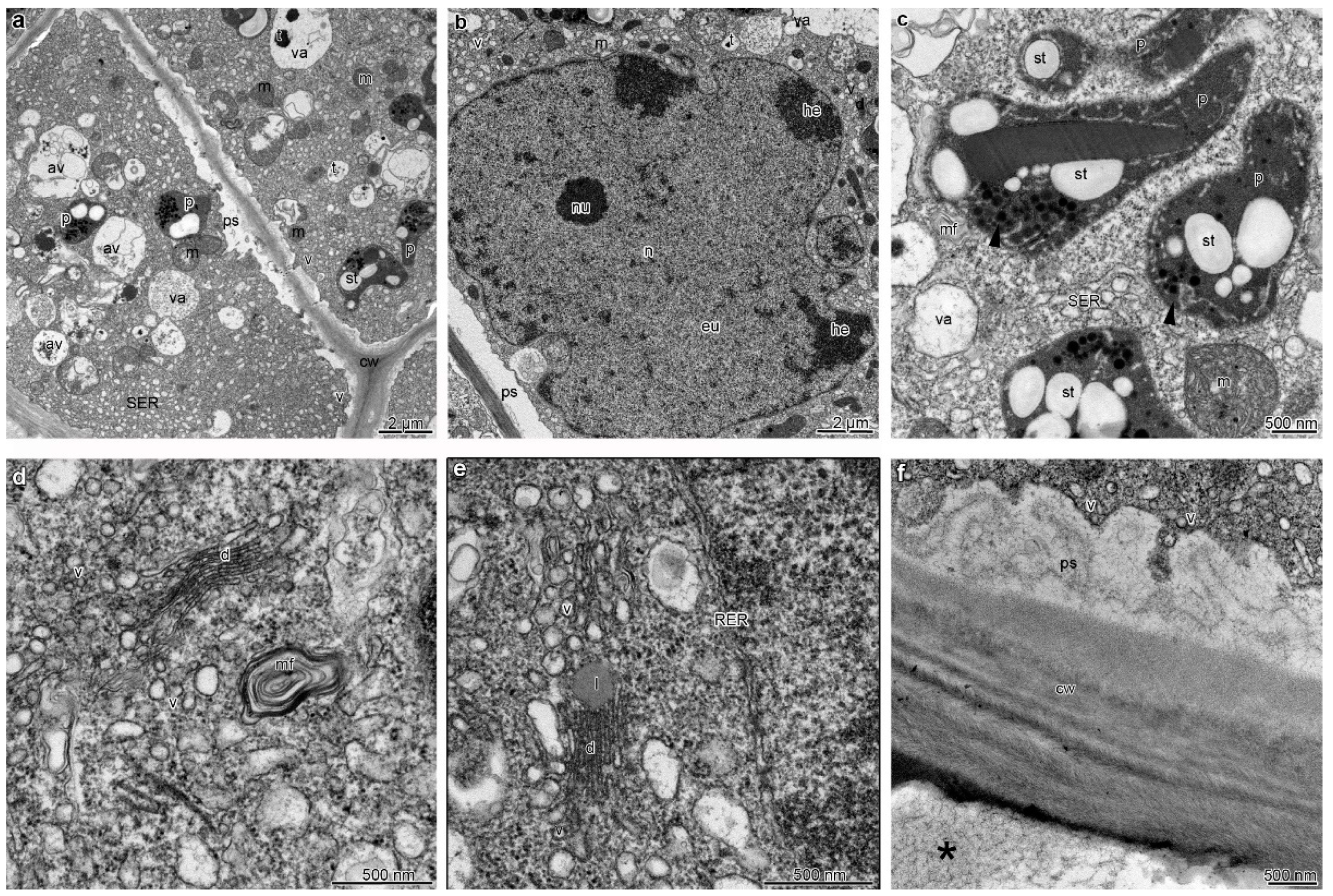
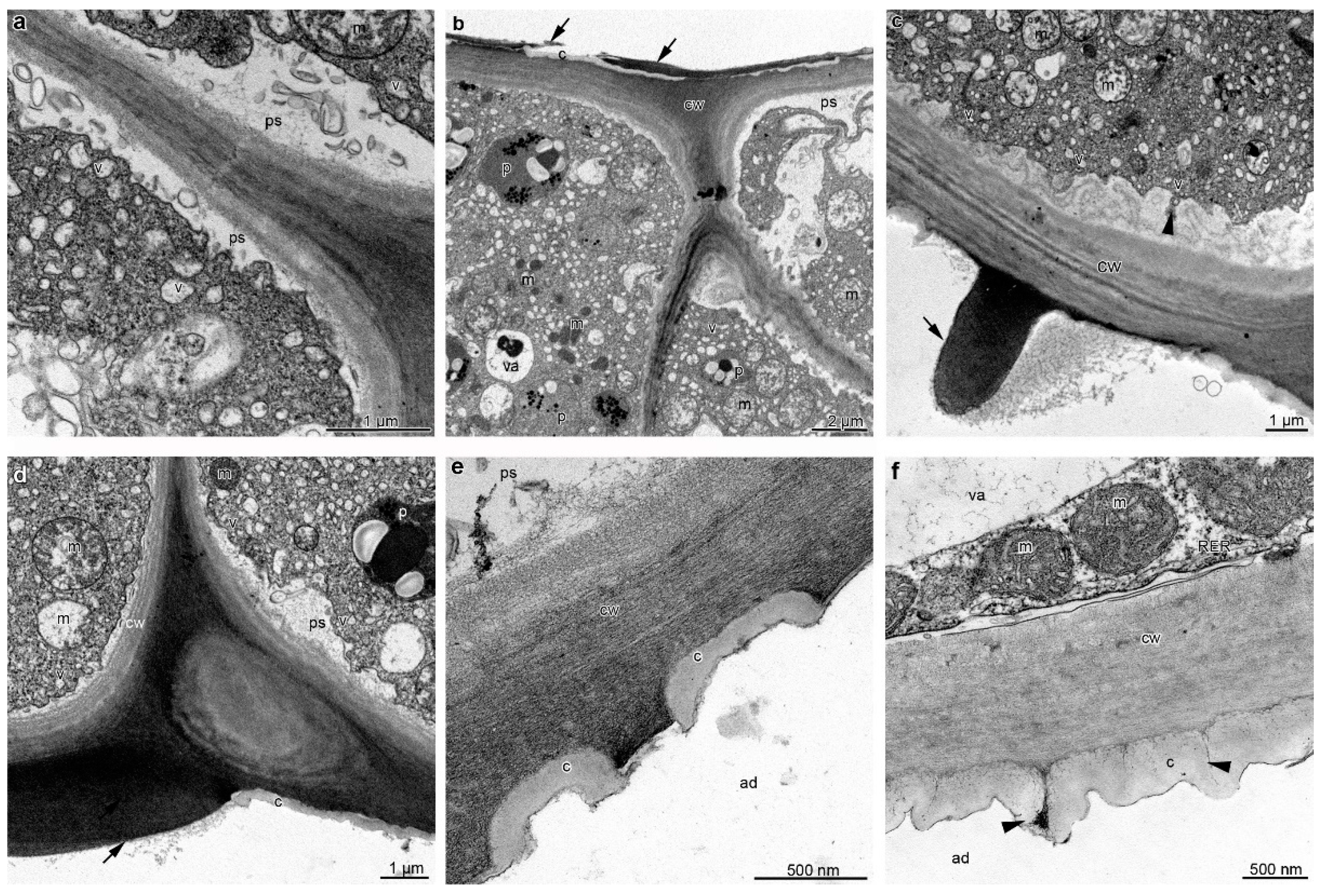
2. Results
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rupp, T.; Oelschlägel, B.; Rabitsch, K.; Mahfoud, H.; Wenke, T.; Disney, R.H.L.; Neinhuis, C.; Wanke, S.; Dötterl, S. Flowers of Deceptive Aristolochia microstoma Are Pollinated by Phorid Flies and Emit Volatiles Known From Invertebrate Carrion. Front. Ecol. Evol. 2021, 9, 658441. [Google Scholar] [CrossRef]
- Brodmann, J.; Twele, R.; Francke, W.; Hölzler, G.; Zhang, Q.H.; Ayasse, M. Orchids mimic Green-Leaf volatiles to attract prey-hunting wasps for pollination. Curr. Biol. 2008, 18, 740–744. [Google Scholar] [CrossRef] [Green Version]
- Urru, I.; Stensmyr, M.C.; Hansson, B.S. Pollination by brood-site deception. Phytochemistry 2011, 72, 1655–1666. [Google Scholar] [CrossRef]
- Heiduk, A.; Brake, I.; von Tschirnhaus, M.; Göhl, M.; Jürgens, A.; Johnson, S.D.; Meve, U.; Dötterl, S. Ceropegia sandersonii mimics attacked honeybees to attract kleptoparasitic flies for pollination. Curr. Biol. 2016, 26, 2787–2793. [Google Scholar] [CrossRef]
- Jürgens, A.; Wee, S.L.; Shuttleworth, A.; Johnson, S.D. Chemical mimicry of insect oviposition sites: A global analysis of convergence in angiosperms. Ecol. Lett. 2013, 16, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewska, N.; Lipińska, M.M.; Gołębiowski, M.; Kowalkowska, A.K. Labellum structure of Bulbophyllum echinolabium J.J. Sm. (section Lepidorhiza Schltr., Bulbophyllinae Schltr., Orchidaceae Juss.). Protoplasma 2019, 256, 1185–1203. [Google Scholar] [CrossRef] [Green Version]
- Martel, C.; Cairampoma, L.; Stauffer, F.W.; Ayasse, M. Telipogon peruvianus (Orchidaceae) flowers elicit pre-mating behaviour in Eudejeania (Tachinidae) males for pollination. PLoS ONE 2016, 11, e0165896. [Google Scholar] [CrossRef]
- Tan, K.H.; Nishida, R. Mutual reproductive benefits between a wild orchid, Bulbophyllum patens, and Bactrocera fruit flies via a floral synomone. J. Chem. Ecol. 2000, 26, 533–546. [Google Scholar] [CrossRef]
- Tan, K.H.; Nishida, R.; Toong, Y.C. Floral synomone of a wild orchid, Bulbophyllum cheiri, lures Bactrocera fruit flies for pollination. J. Chem. Ecol. 2002, 28, 1161–1172. [Google Scholar] [CrossRef]
- Davies, K.L.; Stpiczyńska, M. Labellar anatomy and secretion in Bulbophyllum Thouars (Orchidaceae:Bulbophyllinae) sect. Racemosae Benth & Hook f. Ann. Bot. 2014, 114, 889–911. [Google Scholar]
- Stensmyr, M.C.; Urru, I.; Collu, I.; Celander, M.; Hansson, B.S.; Angioy, A.M. Rotting smell of dead-horse arum florets–these blooms chemically fool flies into pollinating them. Nature 2002, 420, 625–626. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.D.; Jürgens, A. Convergent evolution of carrion and faecal scent mimicry in fly-pollinated angiosperm flowers and a stinkhorn fungus. S. Afr. J. Bot. 2010, 76, 796–807. [Google Scholar] [CrossRef] [Green Version]
- Kite, G.C.; Hetterscheid, W.L.A. Phylogenetic trends in the evolution of inflorescence odours in Amorphophallus. Phytochemistry 2017, 142, 126–142. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Biochemical and molecular genetic aspects of floral scents. Plant Physiol. 2000, 122, 627–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnepp, J.; Dudareva, N. Floral scent: Biosynthesis, regulation and genetic modification. In Annual Plant Reviews Volume 20: Flowering and Its Manipulation; Ainsworth, C., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2006; Volume 20, pp. 240–257. [Google Scholar]
- Ong, P.; Tan, K.H. Fly pollination of four Malaysian species of Bulbophyllum (section Sestochilus)—B. lasianthum, B. lobbii, B. subumbellatum and B. virescens. Males. Orchid J. 2011, 8, 103–110. [Google Scholar]
- Ong, P.; Tan, K.H. Three species of Bulbophyllum section Racemosae pollinated by Drosophila flies. Males. Orchid J. 2012, 9, 45–50. [Google Scholar]
- Tan, K.H.; Nishida, R. Zingerone in the floral synomone of Bulbophyllum baileyi (Orchidaceae) attracts Bactrocera fruit flies during pollination. Biochem. Syst. Ecol. 2007, 35, 334–341. [Google Scholar] [CrossRef]
- Pridgeon, A.M.; Cribb, J.P.; Chase, M.W.; Rasmussen, F.N. Genera Orchidacearum Volume 6 Epidendroidae (Part Three); Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Teixeira, S.D.P.; Borba, E.L.; Semir, J. Lip anatomy and its implications for the pollination mechanisms of Bulbophyllum species(Orchidaceae). Ann. Bot. 2004, 93, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, E.L.P.; Smidt, E.C.; Stützel, T.; Coan, A.I. What do floral anatomy and micromorphology tell us about Neotropical Bulbophyllum section Didactyle (Orchidaceae: Bulbophyllinae)? Bot. J. Linn. Soc. 2014, 175, 438–452. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewska, N.; Kowalkowska, A.K.; Kozieradzka-Kiszkurno, M.; Krawczyńska, A.T.; Bohdanowicz, J. Floral features of two species of Bulbophyllum section Lepidorhiza Schltr.: B. levanae Ames and B. nymphopolitanum Kraenzl. (Bulbophyllinae Schltr., Orchidaceae). Protoplasma 2018, 255, 485–499. [Google Scholar] [CrossRef]
- Gikonyo, N.K.; Hassanali, A.; Njagi, P.G.N.; Gitu, P.M.; Midiwo, J.O. Odor composition of preferred (buffalo and ox) and nonpreferred (waterbuck) hosts of some savanna tsetse flies. J. Chem. Ecol. 2002, 28, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Lipińska, M.M.; Wiśniewska, N.; Gołębiowski, M.; Narajczyk, M.; Kowalkowska, A.K. Floral micromorphology, histochemistry, ultrastructure and chemical composition of floral secretions in three Neotropical Maxillariella species (Orchidaceae). Bot. J. Linn. Soc. 2020, 196, 53–80. [Google Scholar] [CrossRef]
- Ramarathnam, N.; Rubin, L.J. The flavour of cured meat. In Flavor of Meat and Meat Products; Shahidi, F., Ed.; Springer: Boston, MA, USA, 1994; pp. 174–198. [Google Scholar]
- Martin, K.R.; Moré, M.; Hipólito, J.; Charlemagne, S.; Schlumpberger, B.O.; Raguso, R.A. Spatial and temporal variation in volatile composition suggests olfactory division of labor within the trap flowers of Aristolochia gigantea. Flora 2017, 232, 153–168. [Google Scholar] [CrossRef] [Green Version]
- The Pherobase: Database of Pheromones and Semiochemicals. Available online: http://www.pherobase.com (accessed on 1 January 2023).
- Diaz, A.; Kite, G.C. A comparison of the pollination ecology of Arum maculatum and A. italicum in England. Watsonia 2002, 24, 171–181. [Google Scholar]
- Hadacek, F.; Weber, M. Club-shaped organs as additional osmophores within the Sauromatum inflorescence: Odour analysis, ultrastructural changes and pollination aspects. Plant Biol. 2002, 4, 367–383. [Google Scholar] [CrossRef]
- Humeau, L.; Micheneau, C.; Jacquemyn, H.; Gauvin-Bialecki, A.; Fournel, J.; Pailler, T. Sapromyiophily in the native orchid, Bulbophyllum variegatum, on Réunion (Mascarene Archipelago, Indian Ocean). J. Trop. Ecol. 2011, 27, 591–599. [Google Scholar] [CrossRef]
- Formisano, C.; Senatore, F.; Della Porta, G.; Scognamiglio, M.; Bruno, M.; Maggio, A.; Rosselli, S.; Zito, P.; Sajeva, M. Headspace volatile composition of the flowers of Caralluma europaea N.E.Br. (Apocynaceae). Molecules 2009, 14, 4597–4613. [Google Scholar] [CrossRef]
- O’Donoghue, J.L. Neurotoxicity of Industrial and Commercial Chemicals; CRC Press: Boca Raton, FL, USA, 1984; Volume 1. [Google Scholar]
- Gopal, J.V.; Kannabiran, K. Interaction of 2, 5-di-tert-butyl-1, 4-Benzoquinone with Selected Antibacterial Drug Target Enzymes by In silico Molecular Docking Studies. Am. J. Drug Discov. Dev. 2013, 3, 200–205. [Google Scholar] [CrossRef] [Green Version]
- Talalay, P.; Dinkova-Kostova, A.T. Role of nicotinamide quinone oxidoreductase 1 (NQO1) in protection against toxicity of electrophiles and reactive oxygen intermediates. Methods Enzymol. 2004, 382, 355–364. [Google Scholar] [PubMed]
- LeVorse, A.T.; Weiss, A.R.; Pawlak, M. Use of Methyl Benzoic Acid Ester in Perfume Compositions. U.S. Patent No. 7,304,028 B2, 4 December 2007. [Google Scholar]
- Mann, F.; Vanjari, S.; Rosser, N.; Mann, S.; Dasmahapatra, K.K.; Corbin, C.; Linares, M.; Pardo-Diaz, C.; Salazar, C.; Jiggins, C.; et al. The Scent Chemistry of Heliconius Wing Androconia. J. Chem. Ecol. 2017, 43, 843–857. [Google Scholar] [CrossRef]
- The Good Scents Company. Available online: http://www.thegoodscentscompany.com/ (accessed on 1 January 2023).
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI). Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 1 January 2023).
- Stpiczyńska, M.; Davies, K.L.; Kamińska, M. Diverse labellar secretions in African Bulbophyllum (Orchidaceae: Bulbophyllinae) sections Ptiloglossum, Oreonastes and Megaclinium. Bot. J. Linn. Soc. 2015, 179, 266–287. [Google Scholar] [CrossRef] [Green Version]
- Stpiczyńska, M.; Davies, K.L. Evidence for the dual role of floral secretory cells in Bulbophyllum. Acta Biol. Cracov. Ser. Bot. 2016, 58, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Kowalkowska, A.K.; Turzyński, S.; Kozieradzka-Kiszkurno, M.; Wiśniewska, N. Floral structure of two species of Bulbophyllum section Cirrhopetalum Lindl.: B. weberi Ames and B. cumingii (Lindl.) Rchb. f. (Bulbophyllinae Schltr., Orchidaceae). Protoplasma 2017, 254, 1431–1449. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, T.S.; Larson, B.M.; Kevan, P.G.; Inouye, D.W.; Lunau, K. Flies and Flowers II: Floral Attractants and Rewards. J. Poll. Ecol. 2014, 12, 63–94. [Google Scholar] [CrossRef]
- Meve, U.; Liede, S. Floral biology and pollination in stapeliads—New results and a literature review. Plant Syst. Evol. 1994, 192, 99–116. [Google Scholar] [CrossRef]
- Banziger, H.; Pape, T. Flowers, faeces and cadavers: Natural feeding and laying habits of flesh flies in Thailand (Diptera: Sarcophagidae, Sarcophaga spp.). J. Nat. Hist. 2004, 38, 1681–1698. [Google Scholar] [CrossRef]
- Figueiredo, A.C.S.; Pais, M.S. Ultrastructural aspects of the nectary spur of Limodorum abortivum (L) Sw. (Orchidaceae). Ann. Bot. 1992, 70, 325–331. [Google Scholar] [CrossRef]
- Pridgeon, A.M.; Stern, W.L. Osmophores of Scaphosepalum (Orchidaceae). Bot. Gaz. 1985, 146, 115–123. [Google Scholar] [CrossRef]
- Stern, W.L.; Curry, K.J.; Pridgeon, A.M. Osmophores of Stanhopea (Orchidaceae). Amer. J. Bot. 1987, 74, 1323–1331. [Google Scholar] [CrossRef]
- Paiva, E.A.S. How do secretory products cross the plant cell wall to be released? A new hypothesis involving cyclic mechanical actions of the protoplast. Ann. Bot. 2016, 117, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Kowalkowska, A.K.; Pawłowicz, M.; Guzanek, P.; Krawczyńska, A.T. Floral nectary and osmophore of Epipactis helleborine (L.) Crantz (Orchidaceae). Protoplasma 2018, 255, 1811–1825. [Google Scholar] [CrossRef] [Green Version]
- Stpiczyńnska, M.; Davies, K.L. Elaiophore structure and oil secretion in flowers of Oncidium trulliferum Lindl. and Ornithophora radicans (Rchb.f.) Garay & Pabst (Oncidiinae: Orchidaceae). Ann. Bot. 2008, 101, 375–384. [Google Scholar]
- Wiśniewska, N.; Gdaniec, A.; Kowalkowska, A.K. Micromorphological, histochemical and ultrastructural analysis of flower secretory structures in two species pollinated by flies (Diptera) of Asclepiadoideae Burnett. S. Afr. J. Bot. 2021, 137, 60–67. [Google Scholar] [CrossRef]
- van de Meene, A.M.L.; Doblin, M.S.; Bacic, A. The plant secretory pathway seen through the lens of the cell wall. Protoplasma 2017, 254, 75–94. [Google Scholar] [CrossRef]
- Wang, J.; Ding, Y.; Wang, J.; Hillmer, S.; Miao, Y.; Lo, S.W.; Wang, X.; Robinson, D.G.; Jiang, L. EXPO, an exocyst-positive organelle distinct from multivesicular endosomes and autophagosomes, mediates cytosol to cell wall exocytosis in Arabidopsis and tobacco cells. Plant Cell 2010, 22, 4009–4030. [Google Scholar] [CrossRef] [Green Version]
- Jürgens, A.; Dötterl, S.; Meve, U. The chemical nature of fetid floral odours in stapeliads (Apocynaceae-Asclepiadoideae-Ceropegieae). New Phytol. 2006, 172, 452–468. [Google Scholar] [CrossRef]
- Dekeirsschieter, J.; Verheggen, F.J.; Gohy, M.; Hubrecht, F.; Bourguignon, L.; Lognay, G.; Haubruge, E. Cadaveric volatile organic compounds released by decaying pig carcasses (Sus domesticus L.) in different biotopes. Forensic Sci. Int. 2009, 189, 46–53. [Google Scholar] [CrossRef]
- Ollerton, J.; Raguso, R. The sweet stench of decay. New Phytol. 2006, 172, 382–385. [Google Scholar] [CrossRef]
- Tan, K.H.; Nishida, R. Synomone or kairomone? Bulbophyllum apertum flower releases raspberry ketone to attract Bactrocera fruit flies during pollination. J. Chem. Ecol. 2005, 31, 509–519. [Google Scholar]
- Ong, P.T.; Hee, A.K.W.; Wee, S.L.; Tan, K.H. The attraction of flowers of Bulbophyllum (Section Sestochilus) to Bactrocera fruit flies (Diptera: Tephritidae). Males. Orchid J. 2011, 8, 93–102. [Google Scholar]
- Tarkowská, D. Plants are Capable of Synthesizing Animal Steroid Hormones. Molecules 2019, 24, 2585. [Google Scholar] [CrossRef] [Green Version]
- Janeczko, A.; Skoczowski, A. Mammalian sex hormones in plants. Folia Histochem. Cytobiol. 2005, 43, 71–79. [Google Scholar] [PubMed]
- Iino, M.; Nomura, T.; Tamaki, Y.; Yamada, Y.; Yoneyama, K.; Takeuchi, Y.; Mori, M.; Asami, T.; Nakano, T.; Yokota, T. Progesterone: Its occurrence in plants and involvement in plant growth. Phytochemistry 2007, 68, 1664–1673. [Google Scholar] [CrossRef]
- Feder, N.; O’Brien, T.P. Plant microtechnique: Some principles and new methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- Ruzin, S. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Jensen, W. Botanical Histochemistry; Freeman: San Francisco, CA, USA, 1962. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company: New York, NY, USA, 1940. [Google Scholar]
- Gahan, P.B. Plant Histochemistry and Cytochemistry: An Introduction; Academic Press: London, UK, 1984. [Google Scholar]
- Heslop-Harrison, Y. The pollen stigma interaction: Pollen tube penetration in Crocus. Ann. Bot. 1977, 41, 913–922. [Google Scholar] [CrossRef]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Bronner, R. Simultaneous demonstration of lipid and starch in planttissues. Stain Technol. 1975, 50, 1–4. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TR: Retention Time | Relative Content (%) | Bulbophyllum carunculatum Labellum | ||
|---|---|---|---|---|
| 1 | 3.900 | 0.50 | 3,5,5-trimethyl-1-hexene, | Alkane (hydrocarbons) |
| 2 | 4.670 | 0.46 | 2,3-dimethyl-2-heptene (trimethyl-hexene) | Alkane (hydrocarbons) |
| 3 | 8.071 | 3.94 | 3-ethyl-2-methyl-1-heptene | Alkane (hydrocarbons) Noted in the floral scent of B. echinolabium [6] and in the scent of ox carcass [23] |
| 4 | 8.179 | 0.74 | 1,2,3,4,5-pentamethyl-cyclopentane (pentamethyl-cyclopentane) | Alkane (hydrocarbons) Noted in Maxillaria sanguinea (Orchidaceae) [24] |
| 5 | 9.504 | 4.50 | 3-methyl-2-nonene, | Alkene (hydrocarbons) Noted in an aroma of cured pork [25] |
| 6 | 9.614 | 1.06 | 1-methyl-2-propyl-cyclohexane, | Alkane (hydrocarbons) Noted in the floral scent of Orchidaceae: B. echinolabium [6] and M. sanguinea [24] |
| 7 | 21.098 | 1.10 | Dodecane | Aliphatic alkane (hydrocarbons) Noted in the floral scent of Orchidaceae: B. echinolabium [6], M. sanguinea [24], and Aristolochiaceae: A. gigantea [26] Pheromone and allomone Hymenoptera, Coleoptera, found in Araceae and Orchidaceae [27] |
| 8 | 24.891 | 5.14 | Tridecane | Aliphatic alkane (hydrocarbons) Noted in the floral scent of Orchidaceae: B. echinolabium [6], M. sanguinea [24], Aristolochiaceae: A. gigantea [26], and Apocynaceae: Huernia hystrix [12] Attractant of Diptera (Chloropidae and Milichiidae), pheromone and allomone of Hymenoptera, Heteroptera, and Coleoptera [27]. Found mainly in Araceae, Arecaceae, Orchidaceae (Coryanthes spp., Cymbidium spp., Ophrys spp., Phalaenopsis spp.), Magnoliaceae, and Moraceae [27] |
| 9 | 28.404 | 3.27 | Tetradecane | Aliphatic alkane (hydrocarbons) Noted in the floral scent of Orchidaceae: B. echinolabium [6], M. vulganica [24] Pheromone and allomone Hymenoptera, Heteroptera, Coleoptera, and Astigmata [27] |
| 10 | 30.146 | 0.20 | Humulene | Sesquiterpenoid (hydrocarbons) Noted in the floral scent of fly-pollinated Araceae: A. maculatum and A. italicum ssp. neglectum [28], Sauromatum guttatum [29], and Orchidaceae: B. variegatum [30]. Noted also in small amounts in horse dung [12] pheromone Diptera, Hymenoptera, and Coleoptera, and attractant Hymenoptera, Heteroptera, and Coleoptera [27]. Humulene gives a woody, earthy, and spicy fragrance |
| 11 | 30.621 | 2.83 | 2,5-di-tert-butyl-1,4-benzoquinone (DTBBQ) | Dicarbonyl (diketones), a 2,5-disubstituted quinone. Noted in the floral scent of fly-pollinated Carraluma europea [31] It is an antibacterial compound, neurotoxic for humans [32], with antimicrobial activity against Bacillus cereus [33]. Quinones are highly electrophilic compounds that are dietary plant components and arise also from the metabolism of benzene, phenols, and other aromatics, including polycyclic aromatics of environmental origin [34] |
| 12 | 30.905 | 4.92 | Benzoic acid, 3-hydroxy-, methyl ester (Methyl 3-hydroxybenzoate) | Benzoate (an aromatic carboxylic acid ester). Esters of benzoic acids have a pleasant, intense fragrance and are used in the perfume industry [35] |
| 13 | 32.250 | 3.89 | Phenol, 3,5-di-tert-butyl- | Phenol derivative |
| 14 | 32.538 | 11.29 | Benzoic acid, 4-ethoxy-, ethyl ester (Ethyl 4-ethoxybenzoate) | Benzoate (an aromatic carboxylic acid ester). Esters of benzoic acids have a pleasant, intense fragrance and are used in the perfume industry Noted in male wing androconia of the neotropical butterfly Heliconius [36] |
| 15 | 34.796 | 11.69 | Hexadecane | Hydrocarbons (aliphatic alkane). Noted in the floral odor of Arum maculatum [28] and Sauromatum guttatum [29] Odor characteristic: fusel-like, fruity, sweet attractant of papaya fruit fly (Toxotrypana curvicauda), the pheromone of Hymenoptera, Lepidoptera, Diptera, Homoptera, Coleoptera, Iguanidae, and Rodentia. Found in many Orchidaceae (i.e., Aerangis spp., Cattleya spp., Dendrobium spp., Odontoglossum spp., Orphys spp.), Cactaceae, and Araceae [27] |
| 16 | 37.090 | 1.15 | n-hexyl salicylate (Hexyl 2-hydroxybenzoate) | Benzoic acid ester (derivative of benzoic acid) Used as flavoring and odor agents. Scent described as fresh, sweet, herbal, pleasant, natural, and floral [37] |
| 17 | 44.293 | 12.89 | Benzenepropanoic acid, 3,5-bis(1,1-dimethylethyl)-4-hydroxy, methyl ester (Methyl 3-(3,5-di-tert-butyl-4-hydroxyphenyl)propionate) | Phenol derivative Noted in the floral scent of Orchidaceae: B. echinolabium [6] |
| 18 | 61.331 | 30.43 | Pregnane-3,20-dione (5-alpha-pregnan-3,20-dione) | Lipids, steroid compounds. 5-alpha-pregnan-3, 20-dione is part of the protein modification, steroid hormone biosynthesis, and prostate cancer pathways. It is a substrate for probable polyprenol reductase, 3-oxo-5-alpha-steroid 4-dehydrogenase 1, and 3-oxo-5-alpha-steroid 4-dehydrogenase 2 [38]. A biologically active 5-alpha-reduced metabolite of plasma progesterone. It is the immediate precursor of 5-alpha-pregnan-3-alpha-ol-20-one (allopregnanolone), a neuroactive steroid that binds with the gaba(a) receptor. A pregnane was found in the urine of pregnant women and sows. It has anesthetic, hypnotic, and sedative properties [39]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiśniewska, N.; Gołębiowski, M.; Kowalkowska, A.K. Labellum Features and Chemical Composition of Floral Scent in Bulbophyllum carunculatum Garay, Hamer & Siegrist (Section Lepidorhiza Schltr., Bulbophyllinae Schltr., Orchidaceae Juss.). Plants 2023, 12, 1568. https://doi.org/10.3390/plants12071568
Wiśniewska N, Gołębiowski M, Kowalkowska AK. Labellum Features and Chemical Composition of Floral Scent in Bulbophyllum carunculatum Garay, Hamer & Siegrist (Section Lepidorhiza Schltr., Bulbophyllinae Schltr., Orchidaceae Juss.). Plants. 2023; 12(7):1568. https://doi.org/10.3390/plants12071568
Chicago/Turabian StyleWiśniewska, Natalia, Marek Gołębiowski, and Agnieszka K. Kowalkowska. 2023. "Labellum Features and Chemical Composition of Floral Scent in Bulbophyllum carunculatum Garay, Hamer & Siegrist (Section Lepidorhiza Schltr., Bulbophyllinae Schltr., Orchidaceae Juss.)" Plants 12, no. 7: 1568. https://doi.org/10.3390/plants12071568