
Identification of Candidate Genes Involved in the Determinism of Pollen Grain Aperture Morphology by Comparative Transcriptome Analysis in Papaveraceae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
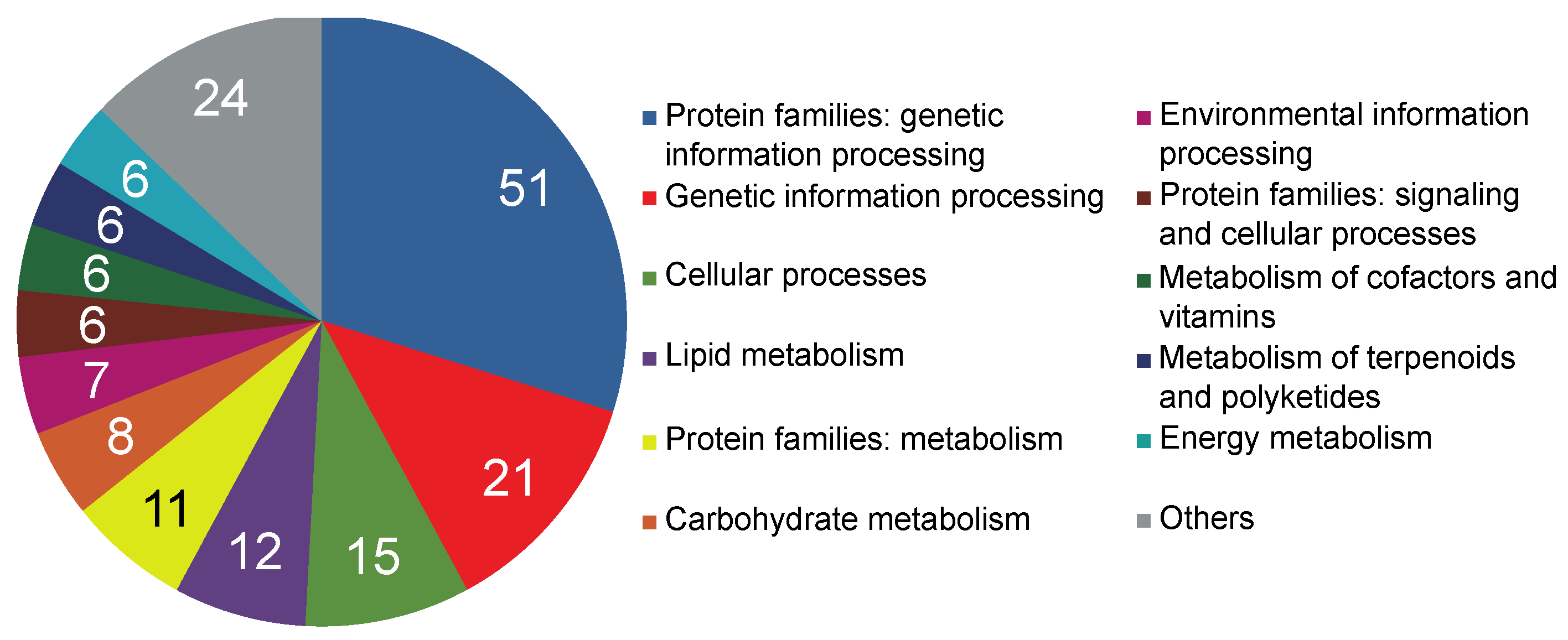{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
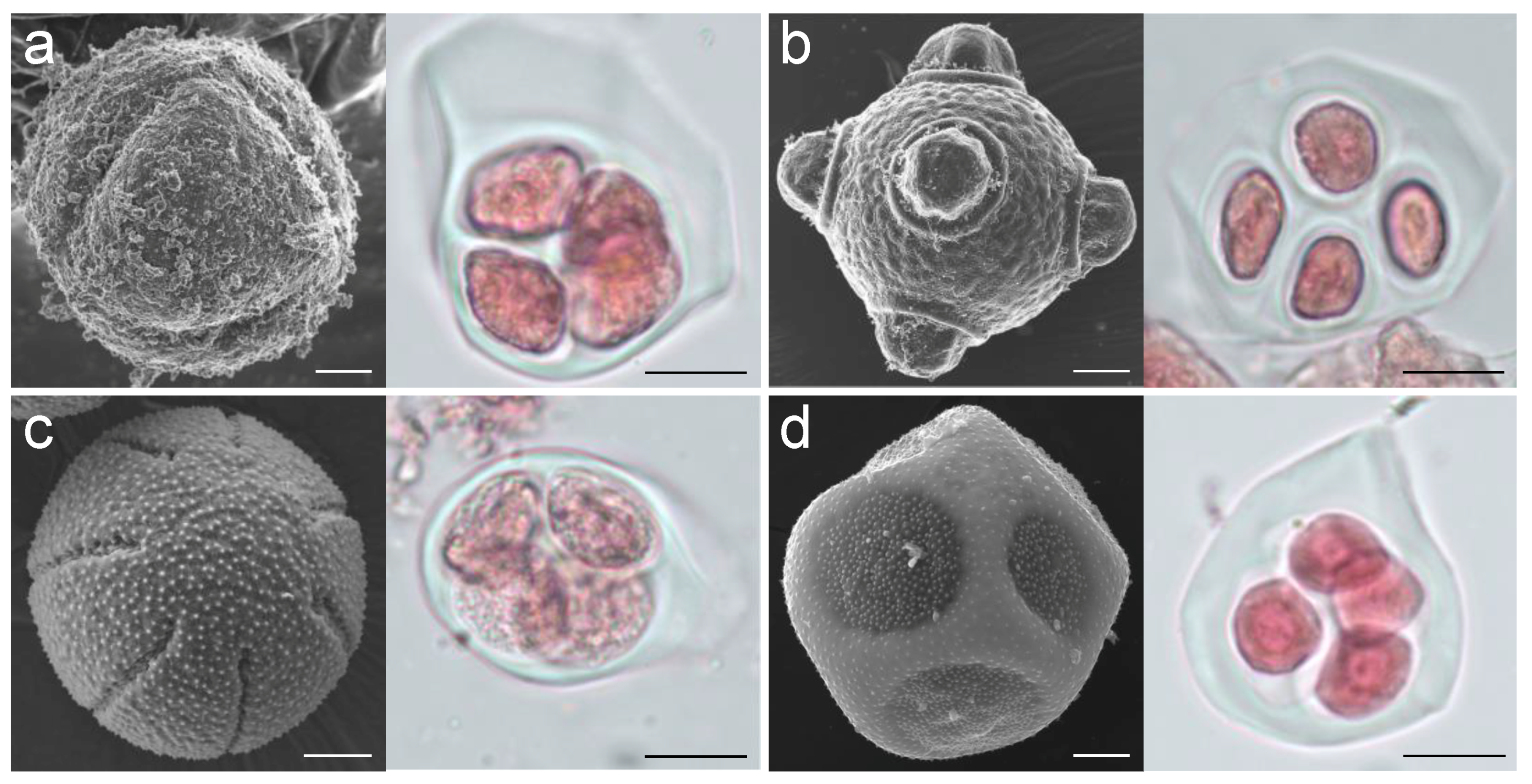
2. Results
2.1. Transcriptome Sequencing and Assembly
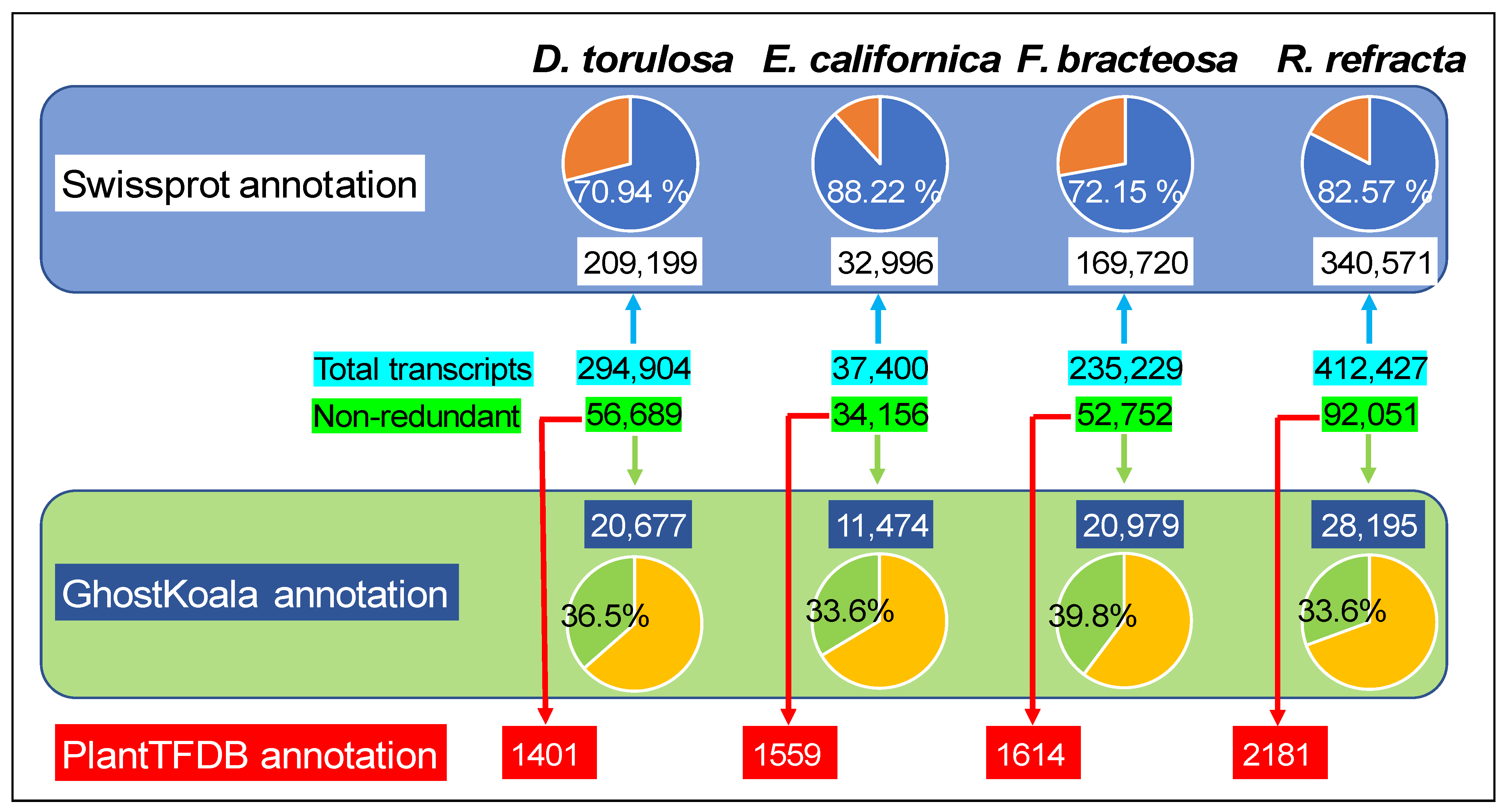
2.2. Transcriptome Functional Annotation and Classification
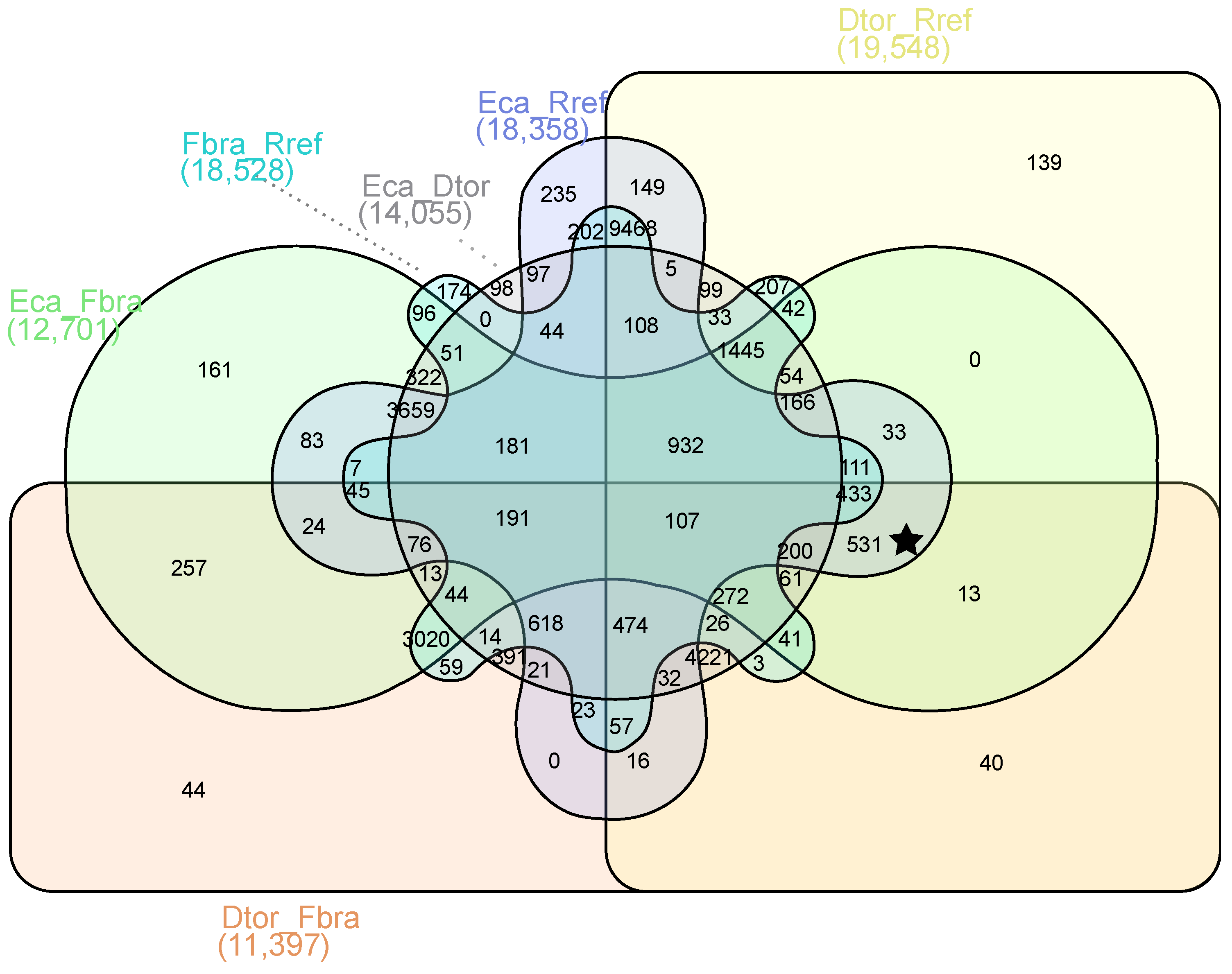
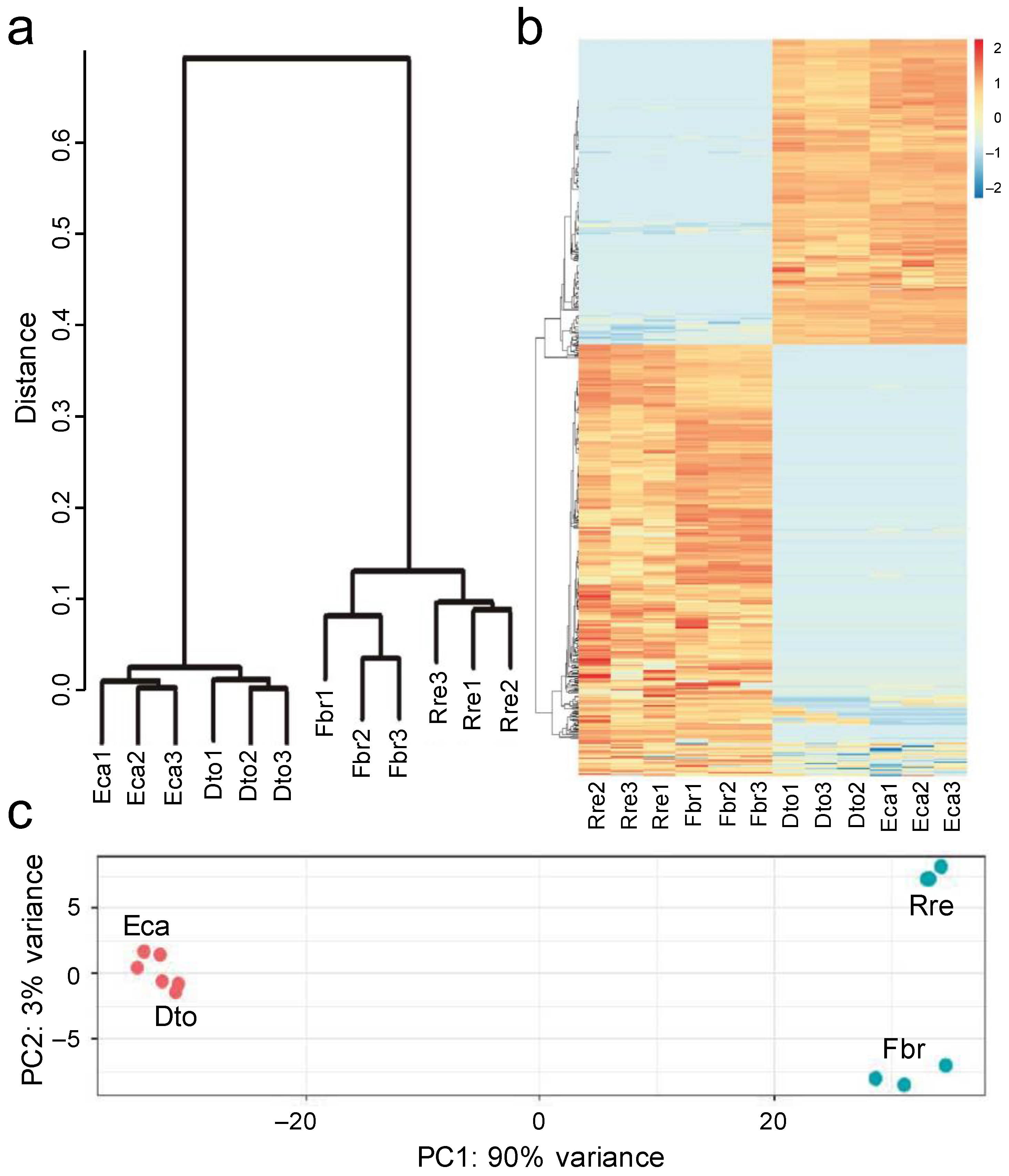
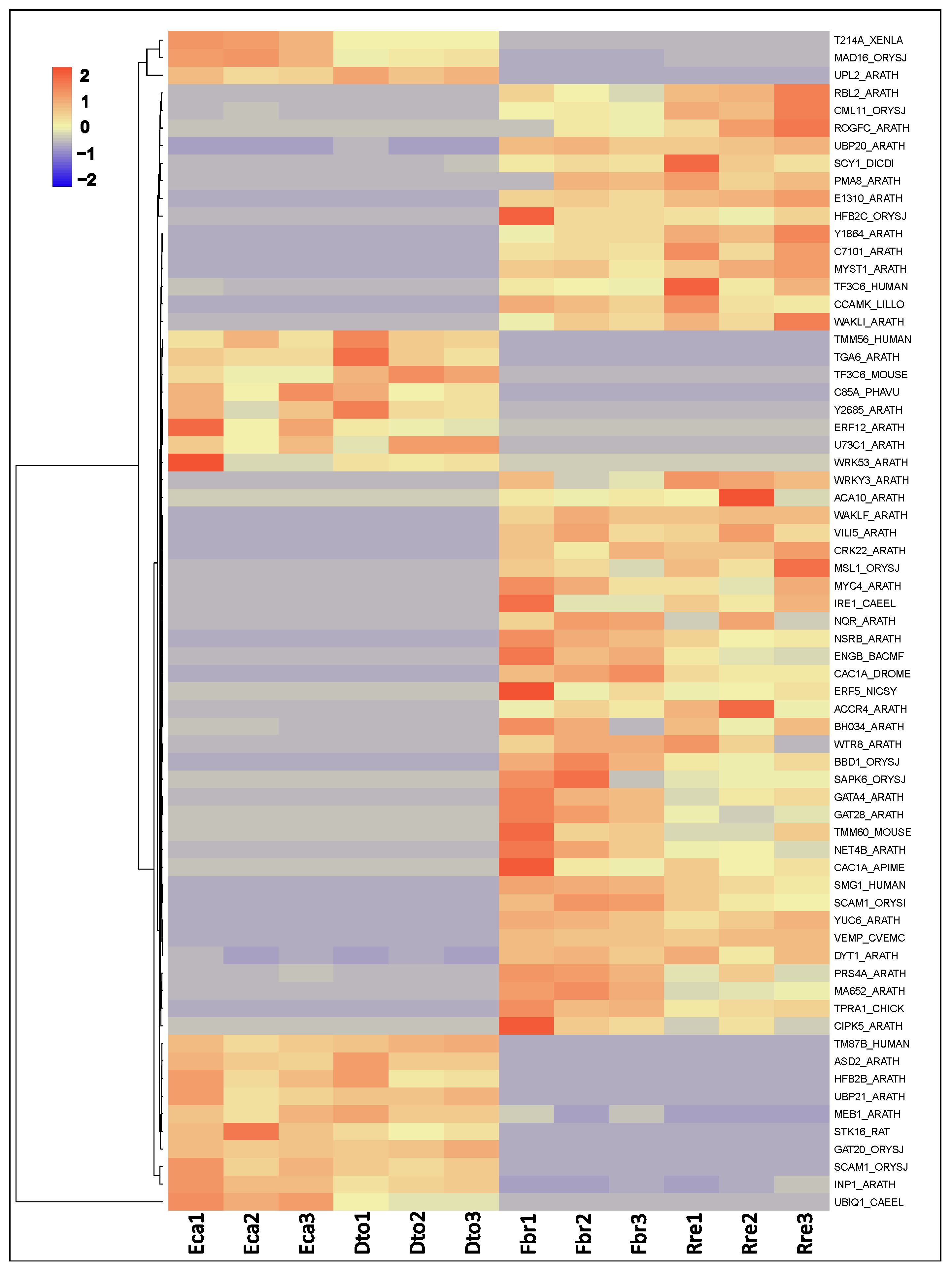
2.3. Differential Gene Expression Analysis
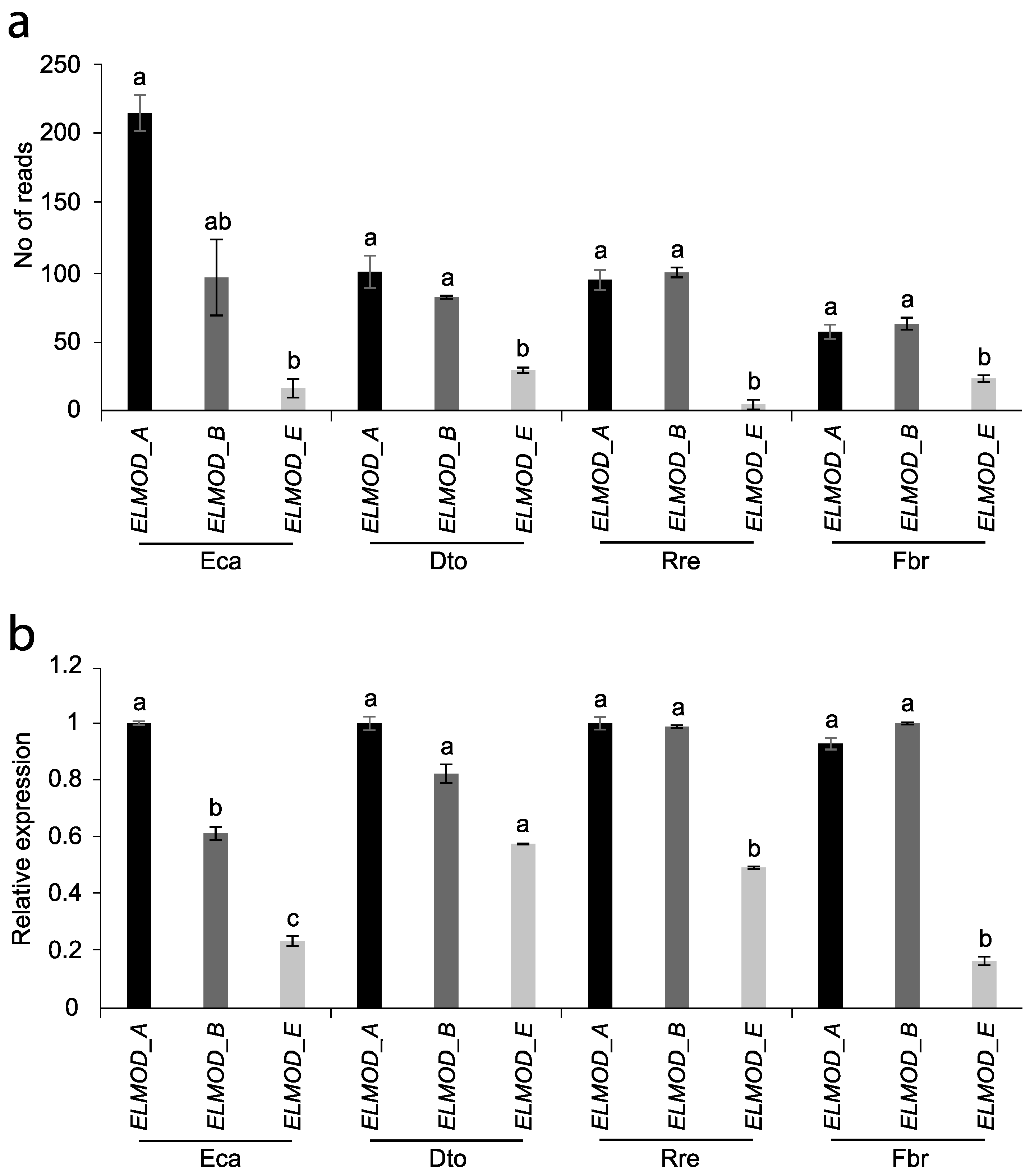
2.4. ELMOD-like Gene Expression
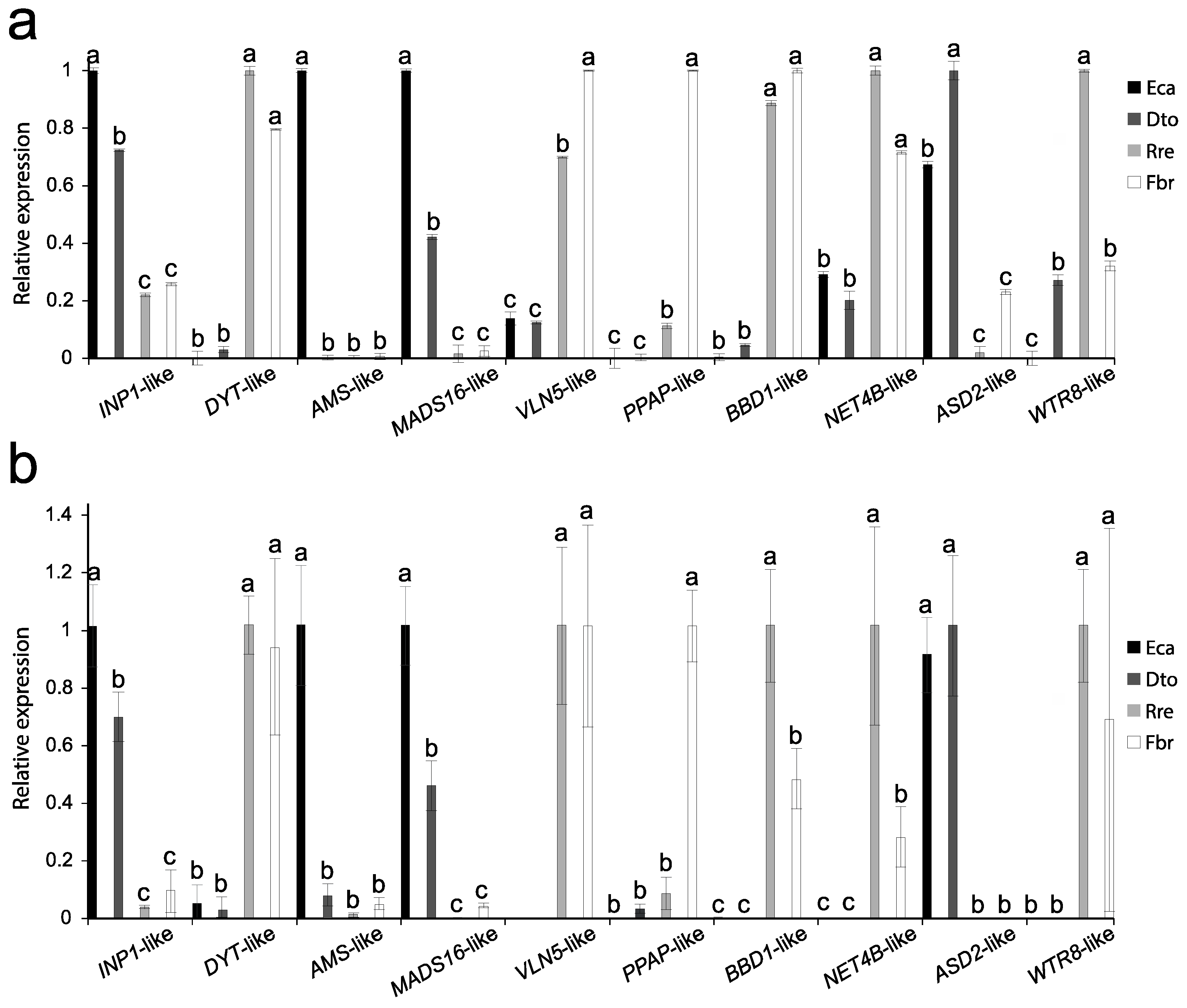
2.5. Verification of RNA-Seq Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. RNA Extraction, Library Construction, Sequencing, and Read Filtering
5.3. De Novo Transcriptome Assembly and Transcript Reconstruction
5.4. Transcript Quantification
5.5. Transcriptome Annotation and Functional Classification
5.6. Analysis of DEGs
5.7. Expression Analysis of ELMOD Genes
5.8. Verification of RNA-Seq Analysis by RT-qPCR
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furness, C.A.; Rudall, P.J. Pollen aperture evolution—A crucial factor for eudicot success? Trends Plant Sci. 2004, 9, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Dobritsa, A.A. Formation of aperture sites on the pollen surface as a model for development of distinct cellular domains. Plant Sci. 2019, 288, 110222. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.W.; Doyle, J.A. The bases of angiosperm phylogeny: Palynology. Ann. Mo. Bot. Gard. 1975, 62, 664–723. [Google Scholar] [CrossRef]
- PalDat A Palynological Database. Available online: https://www.paldat.org. (accessed on 3 May 2021).
- Heslop-Harrison, J. An interpretation of the hydrodynamics of pollen. Am. J. Bot. 1979, 66, 737–743. [Google Scholar] [CrossRef]
- Matamoro-Vidal, A.; Raquin, C.; Brisset, F.; Colas, H.; Izac, B.; Albert, B.; Gouyon, P.H. Links between morphology and function of the pollen wall: An. experimental approach. Bot. J. Linn. Soc. 2016, 180, 478–490. [Google Scholar] [CrossRef] [Green Version]
- Wodehouse, R. Pollen Grains, Their Structure, Identification and Significance in Science and Medicine; McGraw-Hill: New York, NY, USA, 1935. [Google Scholar]
- Volkova, O.A.; Severova, E.E.; Polevova, S.V. Structural basis of harmomegathy: Evidence from Boraginaceae pollen. Plant Syst. Evol. 2013, 299, 1769–1779. [Google Scholar] [CrossRef]
- Božič, A.; Šiber, A. Mechanical design of apertures and the infolding of pollen grain. Proc. Natl. Acad. Sci. USA 2020, 117, 26600–26607. [Google Scholar] [CrossRef]
- Katifori, E.; Alben, S.; Cerda, E.; Nelson, D.R.; Dumais, J. Foldable structures and the natural design of pollen grains. Proc. Natl. Acad. Sci. USA 2010, 107, 7635–7639. [Google Scholar] [CrossRef] [Green Version]
- Payne, W.W. Observations of harmomegathy in pollen of Anthophyta. Grana 1972, 12, 93–98. [Google Scholar] [CrossRef]
- Prieu, C.; Sauquet, H.; Gouyon, P.H.; Albert, B. More than sixty origins of pantoporate pollen in angiosperms. Am. J. Bot. 2017, 104, 1837–1845. [Google Scholar] [CrossRef] [Green Version]
- Franchi, G.G.; Nepi, M.; Dafni, A.; Pacini, E. Partially hydrated pollen: Taxonomic distribution, ecological and evolutionary significance. Plant Syst. Evol. 2002, 234, 211–227. [Google Scholar] [CrossRef]
- Dobritsa, A.A.; Coerper, D. The novel plant protein INAPERTURATE POLLEN1 marks distinct cellular domains and controls formation of apertures in the Arabidopsis pollen exine. Plant Cell 2012, 24, 4452–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.H.; Weber, Z.T.; Zourelidou, M.; Hofmeister, B.T.; Schmitz, R.J.; Schwechheimer, C.; Dobritsa, A.A. Arabidopsis protein kinase D6PKL3 is involved in the formation of distinct plasma membrane aperture domains on the pollen surface. Plant Cell 2018, 30, 2038–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhao, G.; Tan, Q.; Yuan, H.; Betts, N.; Zhu, L.; Zhang, D.; Liang, W. Rice pollen aperture formation is regulated by the interplay between OsINP1 and OsDAF1. Nat. Plants 2020, 6, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Wang, R.; Moberg, I.M.; Reeder, S.H.; Amom, P.; Tan, M.H.; Amstutz, K.; Chandna, P.; Helton, A.; Andrianova, E.P.; et al. A species-specific functional module controls formation of pollen apertures. Nat. Plants 2021, 7, 966–978. [Google Scholar] [CrossRef]
- Zhou, Y.; Amom, P.; Reeder, S.H.; Lee, B.H.; Helton, A.; Dobritsa, A.A. Members of the ELMOD protein family specify formation of distinct aperture domains on the Arabidopsis pollen surface. Elife 2021, 10, e71061. [Google Scholar] [CrossRef]
- Li, P.; Ben-Menni Schuler, S.; Reeder, S.H.; Wang, R.; Suárez Santiago, V.N.; Dobritsa, A.A. INP1 involvement in pollen aperture formation is evolutionarily conserved and may require species-specific partners. J. Exp. Bot. 2018, 69, 983–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazuecos-Aguilera, I.; Romero-García, A.T.; Klodová, B.; Honys, D.; Fernández-Fernández, M.C.; Schuler, S.B.-M.; Dobritsa, A.A.; Suárez-Santiago, V.N. The role of INAPERTURATE POLLEN1 as a pollen aperture factor is conserved in the basal eudicot Eschscholzia californica (Papaveraceae). Front. Plant Sci. 2021, 12, 1405. [Google Scholar] [CrossRef]
- Pérez-Gutiérrez, M.A.; Romero-García, A.T.; Fernández, M.C.; Blanca, G.; Salinas-Bonillo, M.J.; Suárez-Santiago, V.N. Evolutionary history of fumitories (subfamily Fumarioideae, Papaveraceae): An old story shaped by the main geological and climatic events in the Northern Hemisphere. Mol. Phylogenetics Evol. 2015, 88, 75–92. [Google Scholar] [CrossRef]
- Wang, R.; Dobritsa, A.A. Exine and aperture patterns on the pollen surface: Their formation and roles in plant reproduction. Annu. Plant Rev. Online 2018, 1, 589–628. [Google Scholar] [CrossRef]
- Albert, B.; Nadot, S.; Dreyer, L.; Ressayre, A. The influence of tetrad shape and intersporal callose wall formation on pollen aperture pattern ontogeny in two eudicot species. Ann. Bot. 2010, 106, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Prieu, C.; Toghranegar, Z.; Matamoro-Vidal, A.; Nadot, S.; Albert, B. Additional callose deposits are located at the future apertural regions in sulcate, ulcerate, porate, colporate, colpate and syncolpate pollen grains. Biol. J. Linn. Soc. 2017, 183, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Lu, D.; Ma, X.; Peng, Y.; Sun, Y.; Ning, G.; Ma, H. Regulation of the Arabidopsis anther transcriptome by DYT1 for pollen development. Plant J. 2012, 72, 612–624. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, H.; Li, R.; Hong, Q.; Li, Y.; Xia, Q.; Zhang, W. Function identification of the nucleotides in key cis-element of Dysfunctional Tapetum1 (DYT1) promoter. Front. Plant. Sci. 2017, 8, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Sun, Y.; Timofejeva, L.; Chen, C.; Grossniklaus, U.; Ma, H. Regulation of Arabidopsis tapetum development and function by DYSFUNCTIONAL TAPETUM1 (DYT1) encoding a putative bHLH transcription factor. Development 2006, 133, 3085–3095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.; Xu, X.F.; Zhu, J.; Gu, J.N.; Blackmore, S.; Yang, Z.N. The tapetal AHL family protein TEK determines nexine formation in the pollen wall. Nat. Commun. 2014, 5, 3855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, S.-X.; Zeng, Q.-Y.; Hou, J.-Q.; Hou, L.-L.; Zhu, J.; Yang, M.; Yang, Z.-N.; Lou, Y. The temporal regulation of TEK contributes to pollen wall exine patterning. PLoS Genet. 2020, 16, e1008807. [Google Scholar] [CrossRef]
- Stieglitz, H. Role of β-1, 3-glucanase in postmeiotic microspore release. Dev. Biol. 1977, 57, 87–97. [Google Scholar] [CrossRef]
- Levy, A.; Erlanger, M.; Rosenthal, M.; Epel, B.L. A plasmodesmata-associated β-1, 3-glucanase in Arabidopsis. Plant J. 2007, 49, 669–682. [Google Scholar] [CrossRef]
- Delp, G.; Palva, E.T. A novel flower-specific Arabidopsis gene related to both pathogen-induced and developmentally regulated plant β-1, 3-glucanase genes. Plant Mol. Biol. 1999, 39, 565–575. [Google Scholar] [CrossRef]
- You, M.K.; Shin, H.Y.; Kim, Y.J.; Ok, S.H.; Cho, S.K.; Jeung, J.U.; Yoo, S.D.; Kim, J.K.; Shin, J.S. Novel bifunctional nucleases, OmBBD and AtBBD1, are involved in abscisic acid-mediated callose deposition in Arabidopsis. Plant Physiol. 2010, 152, 1015–1029. [Google Scholar] [CrossRef] [Green Version]
- Huque, A.K.M.; So, W.M.; You, M.K.; Shin, J.S. Phylogenetic Analysis and In Vitro Bifunctional Nuclease Assay of Arabidopsis BBD1 and BBD2. Molecules 2020, 25, 2169. [Google Scholar] [CrossRef] [PubMed]
- Ranocha, P.; Denancé, N.; Vanholme, R.; Freydier, A.; Martinez, Y.; Hoffmann, L.; Köhler, L.; Pouzet, C.; Renou, J.-P.; Sundberg, B.; et al. WALLS ARE THIN 1 (WAT1), an Arabidopsis homolog of Medicago truncatula NODULIN21, is a tonoplast-localized protein required for secondary wall formation in fibers. Plant J. 2010, 63, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Busov, V.B.; Johannes, E.; Whetten, R.W.; Sederoff, R.R.; Spiker, S.L.; Lanz-Garcia, C.; Goldfarb, B. An auxin-inducible gene from loblolly pine (Pinus taeda L.) is differentially expressed in mature and juvenile-phase shoots and encodes a putative transmembrane protein. Planta 2004, 218, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Pomares-Viciana, T.; Del Río-Celestino, M.; Román, B.; Die, J.; Pico, B.; Gómez, P. First RNA-seq approach to study fruit set and parthenocarpy in zucchini (Cucurbita pepo L.). BMC Plant Biol. 2019, 19, 61. [Google Scholar] [CrossRef] [Green Version]
- Ressayre, A.; Raquin, C.; Mignot, A.; Godelle, B.; Gouyon, P.H. Correlated variation in microtubule distribution, callose deposition during male post-meiotic cytokinesis, and pollen aperture number across Nicotiana species (Solanaceae). Am. J. Bot. 2002, 89, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, C.A.P.; Webster, B.D.; Jernstedt, J.A. Spatial congruence between exine pattern, microtubules and endomembranes in Vigna pollen. Sex. Plant Reprod. 1995, 8, 147–151. [Google Scholar] [CrossRef]
- Sheldon, J.M.; Dickinson, H.G. Pollen wall formation in Lilium: The effect of chaotropic agents, and the organisation of the microtubular cytoskeleton during pattern development. Planta 1986, 168, 11–23. [Google Scholar] [CrossRef]
- Ariizumi, T.; Toriyama, K. Genetic regulation of sporopollenin synthesis and pollen exine development. Annu. Rev. Plant Biol. 2011, 62, 437–460. [Google Scholar] [CrossRef]
- Deeks, M.J.; Calcutt, J.R.; Ingle, E.K. A superfamily of actin-binding proteins at the actin-membrane nexus of higher plants. Curr. Biol. 2012, 22, 1595–1600. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, S.; Eisa, A.; Kleine-Vehn, J.; Scheuring, D. NET4 modulates the compactness of vacuoles in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 4752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Qu, X.; Bao, C.; Khurana, P.; Wang, Q.; Xie, Y.; Zheng, Y.; Chen, N.; Blanchoin, L.; Staiger, C.J.; et al. Arabidopsis VILLIN5, an actin filament bundling and severing protein, is necessary for normal pollen tube growth. Plant Cell 2010, 22, 2749–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, M.C.; Romero García, A.T.; Rodríguez García, M.I. Aperture structure, development and function in Lycopersicum esculentum Miller (Solanaceae) pollen grain. Rev. Palaeobot. Palynol. 1992, 72, 41–48. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.; Papanicolaou, A. TransDecoder (Find Coding Regions within Transcripts). Available online: http://transdecoder.github.io (accessed on 15 March 2020).
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2016, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650. [Google Scholar] [CrossRef]
- Bairoch, A.; Boeckmann, B. The SWISS-PROT protein sequence data bank. Nucleic Acids Res. 1992, 20, 2019. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Götz, S.; Garcia-Gomez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Jin, J.; Tang, L.; Zhao, Y.; Gu, X.; Gao, G.; Luo, J. PlantTFDB 2.0: Update and improvement of the comprehensive plant transcription factor database. Nucleic Acids Res. 2011, 39, D1114–D1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazuecos-Aguilera, I.; Suárez-Santiago, V.N. Identification of Candidate Genes Involved in the Determinism of Pollen Grain Aperture Morphology by Comparative Transcriptome Analysis in Papaveraceae. Plants 2023, 12, 1570. https://doi.org/10.3390/plants12071570
Mazuecos-Aguilera I, Suárez-Santiago VN. Identification of Candidate Genes Involved in the Determinism of Pollen Grain Aperture Morphology by Comparative Transcriptome Analysis in Papaveraceae. Plants. 2023; 12(7):1570. https://doi.org/10.3390/plants12071570
Chicago/Turabian StyleMazuecos-Aguilera, Ismael, and Víctor N. Suárez-Santiago. 2023. "Identification of Candidate Genes Involved in the Determinism of Pollen Grain Aperture Morphology by Comparative Transcriptome Analysis in Papaveraceae" Plants 12, no. 7: 1570. https://doi.org/10.3390/plants12071570