
Invasion of a Horticultural Plant into Forests: Lamium galeobdolon argentatum Affects Native Above-Ground Vegetation and Soil Properties

Abstract
:1. Introduction
- (1)
- (2)
- Nehring et al. [23] reported that L. g. argentatum is now more common in suburban and urban forests than the native subspecies. This could be due to better performance of the horticultural L. g. argentatum compared to the native subspecies. We therefore expected that L. g. argentatum has a higher establishment rate of juvenile plants derived from clonal propagation, a greater number of stolons and longer stolons than native L. g. galeobdolon. Furthermore, we hypothesized that the horticultural subspecies has a greater patch growth rate and forms larger plant patches than the native subspecies.
- (3)
- We also hypothesized that compared to the native L. g. galeobdolon, the non-native L. g. argentatum reduces the richness of native plants in the ground vegetation, changes the composition of native plant species and soil properties, and alters the abundance and composition of soil microbial organisms.
2. Results
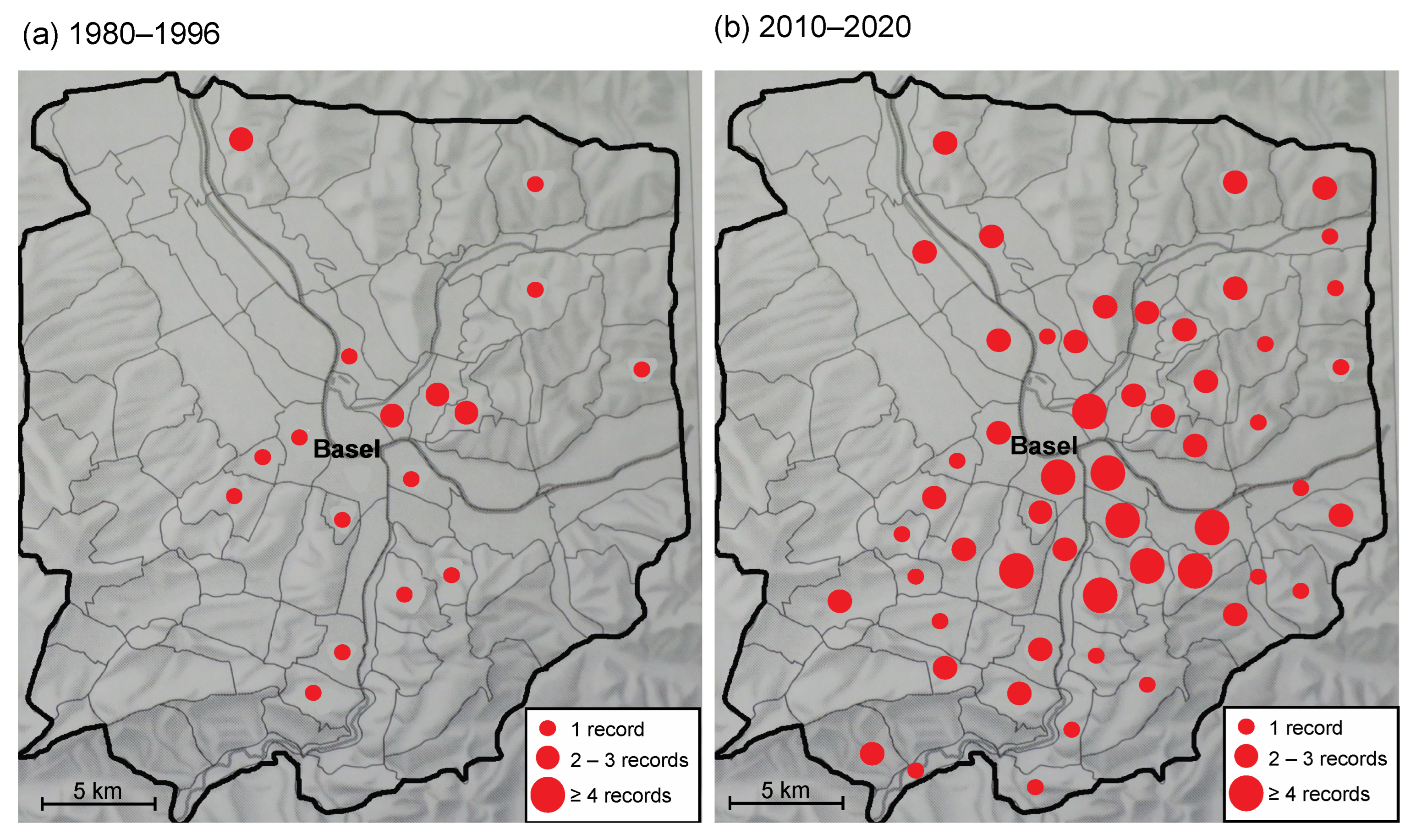
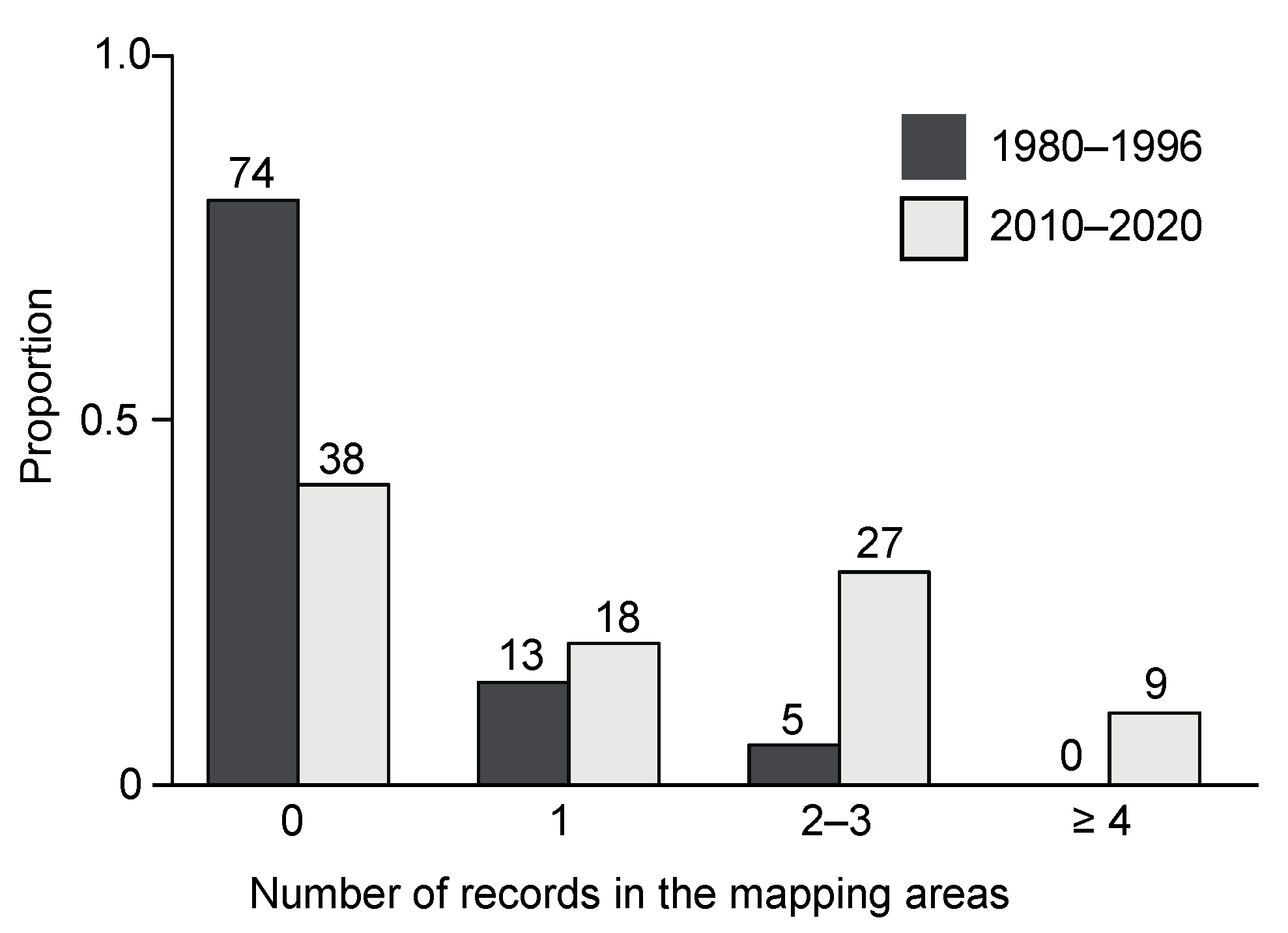
2.1. Spread of L. g. argentatum in the Region of Basel
2.2. Performance of Lamium subsp. in Swiss Forests
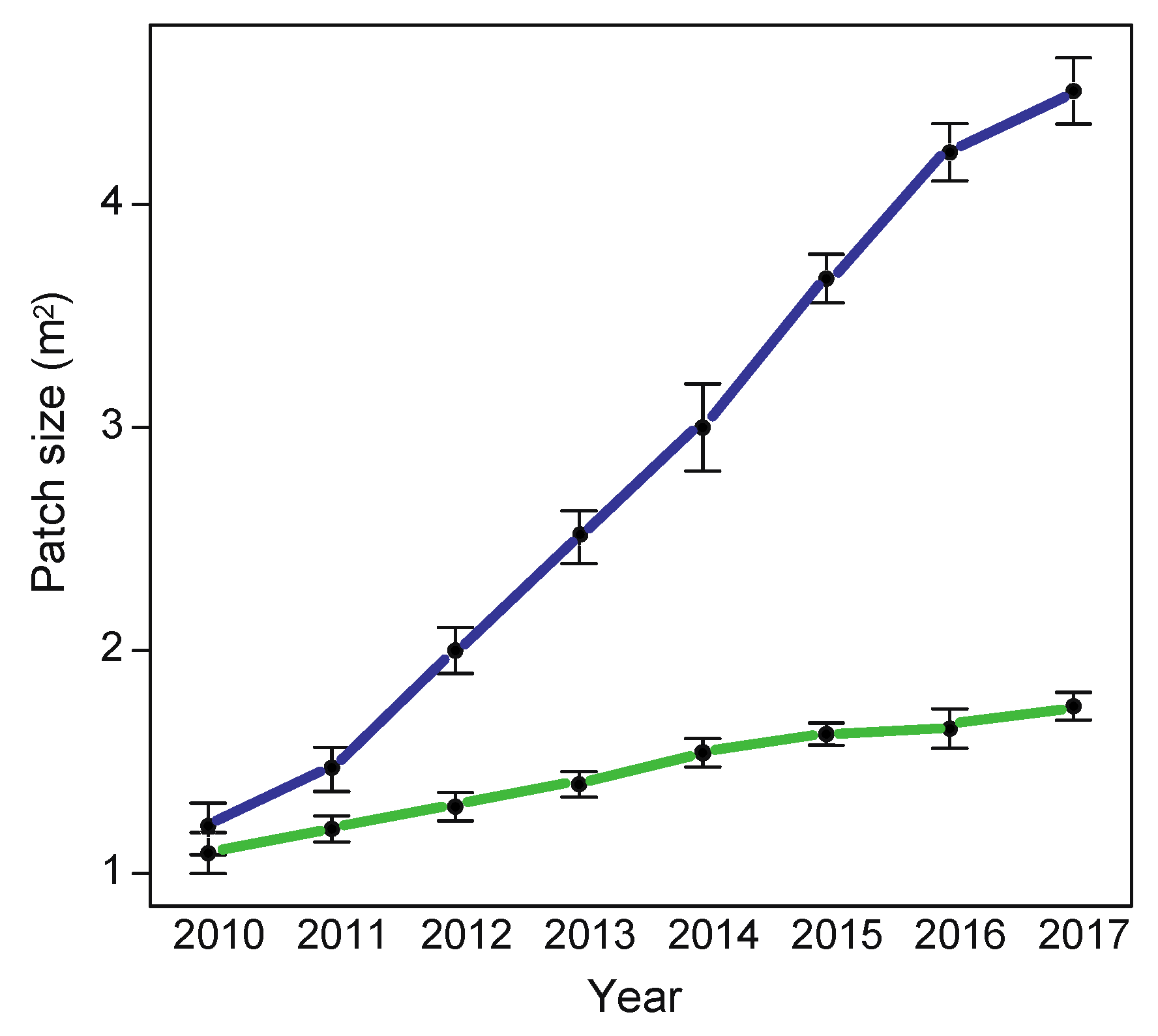
2.3. Growth of L. g. argentatum and L. g. galeobdolon Patches over 7 Years
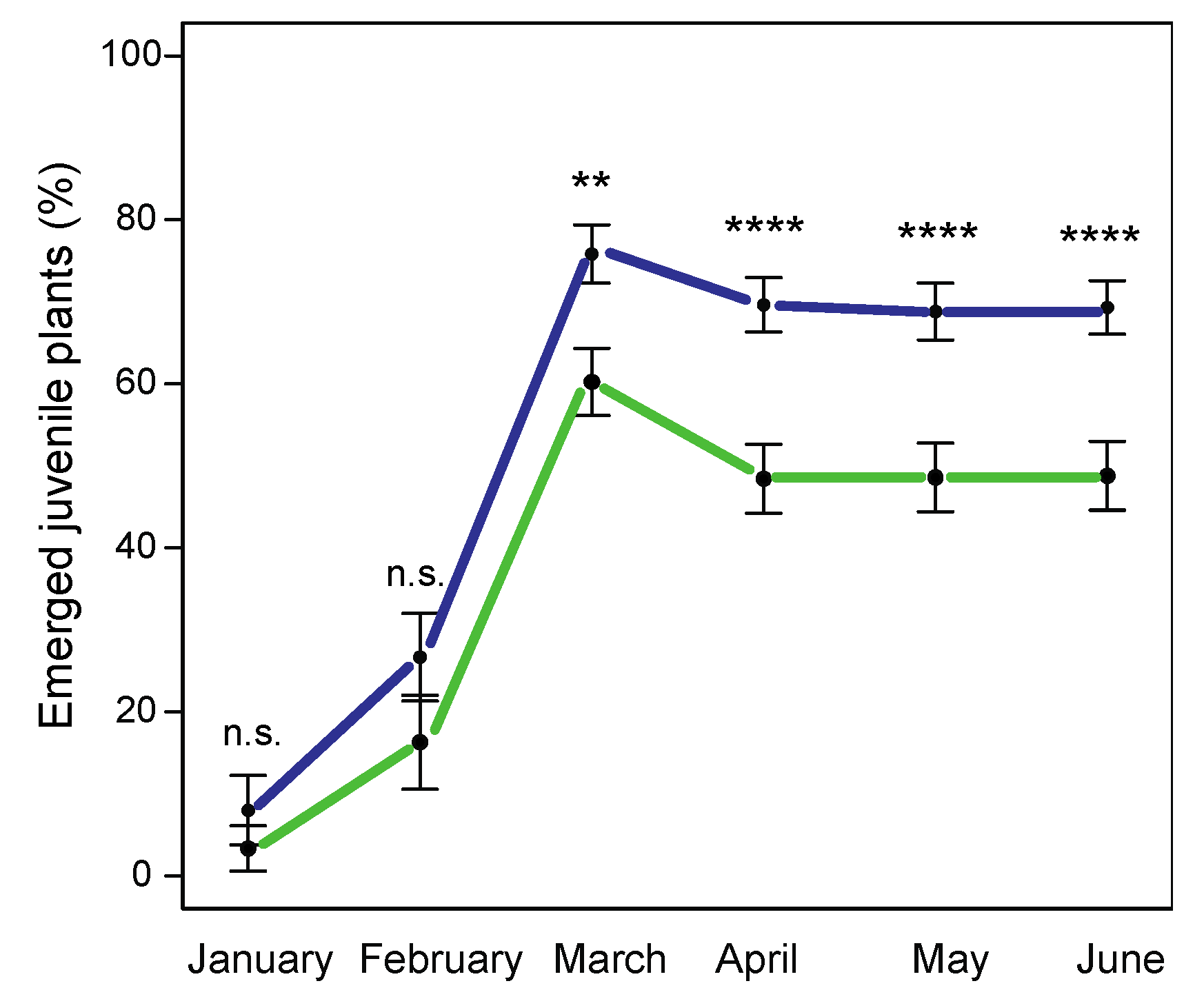
2.4. Performance under Standardized Conditions
2.5. Impact on Native Ground Vegetation and Soil Properties
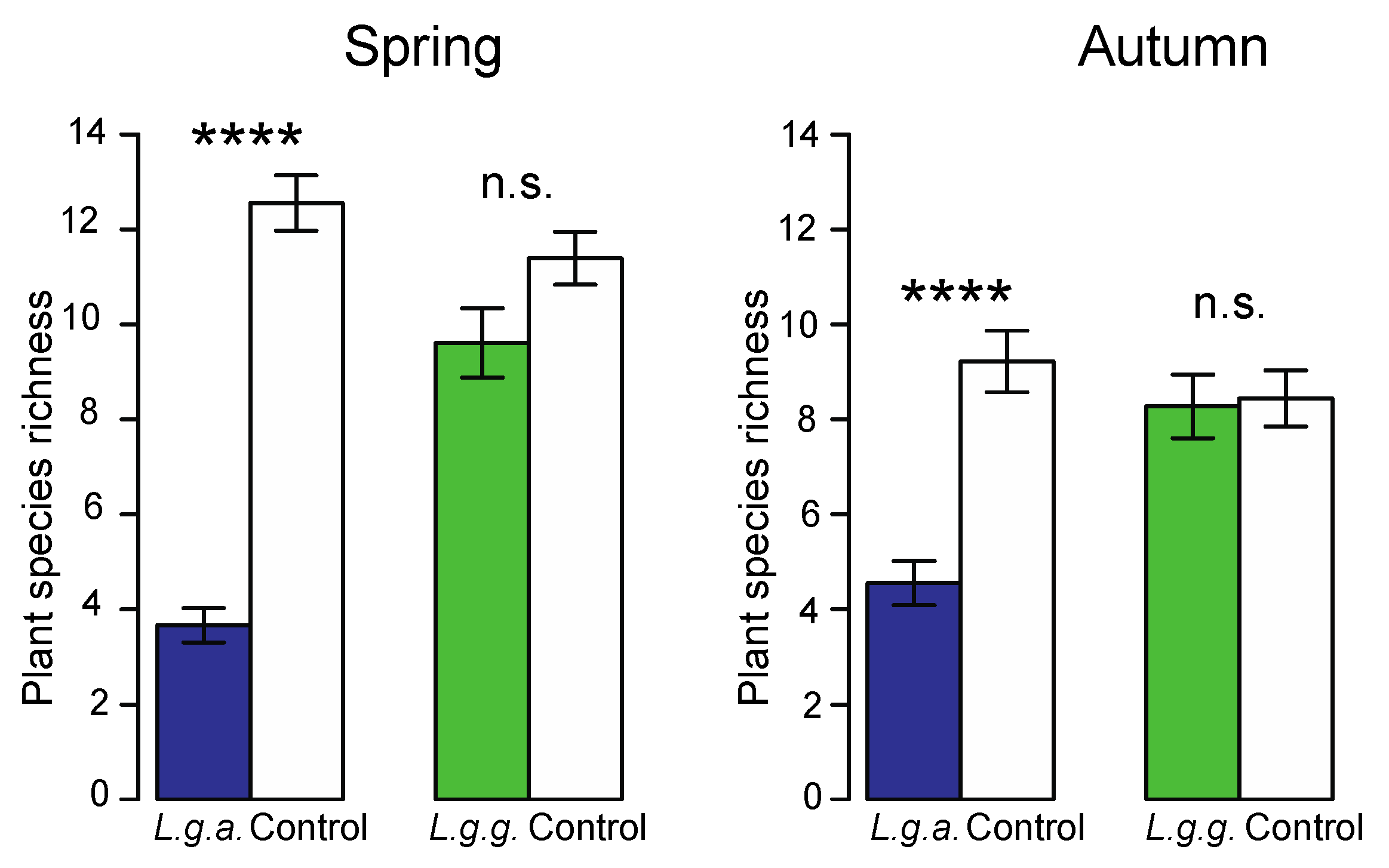
2.5.1. Native Ground Vegetation
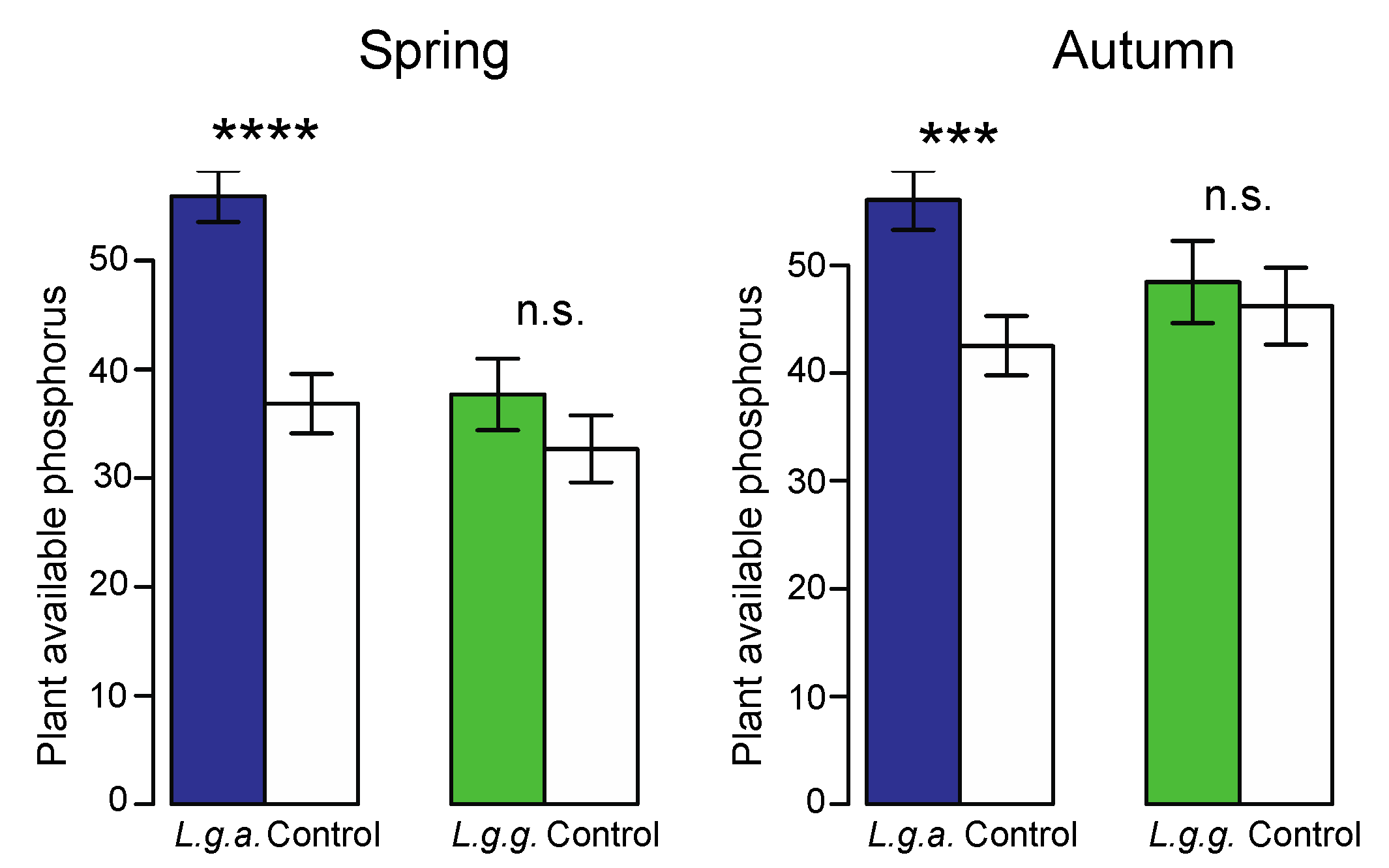
2.5.2. Physical and Chemical Soil Properties
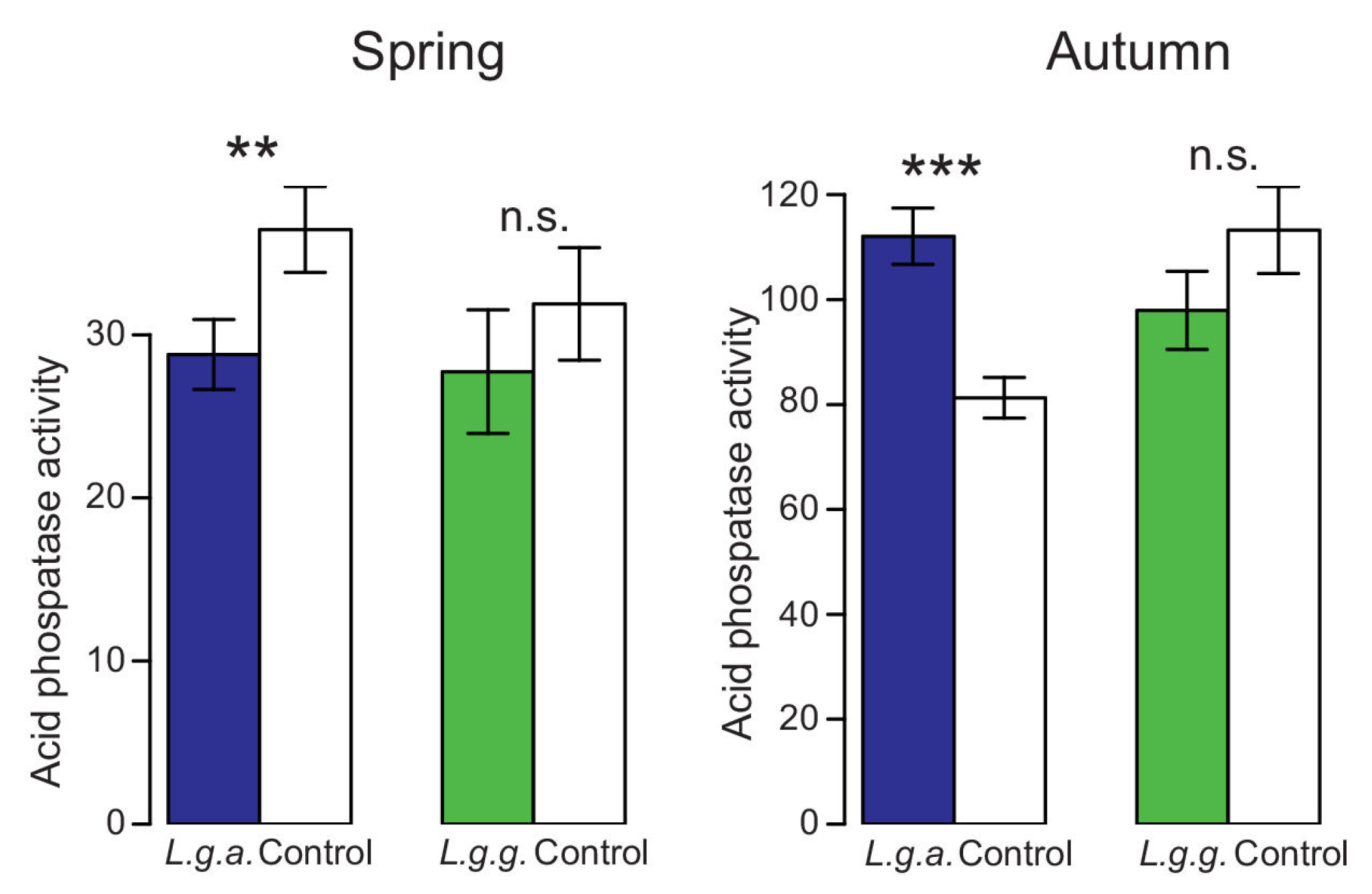
2.5.3. Soil Enzyme Activity
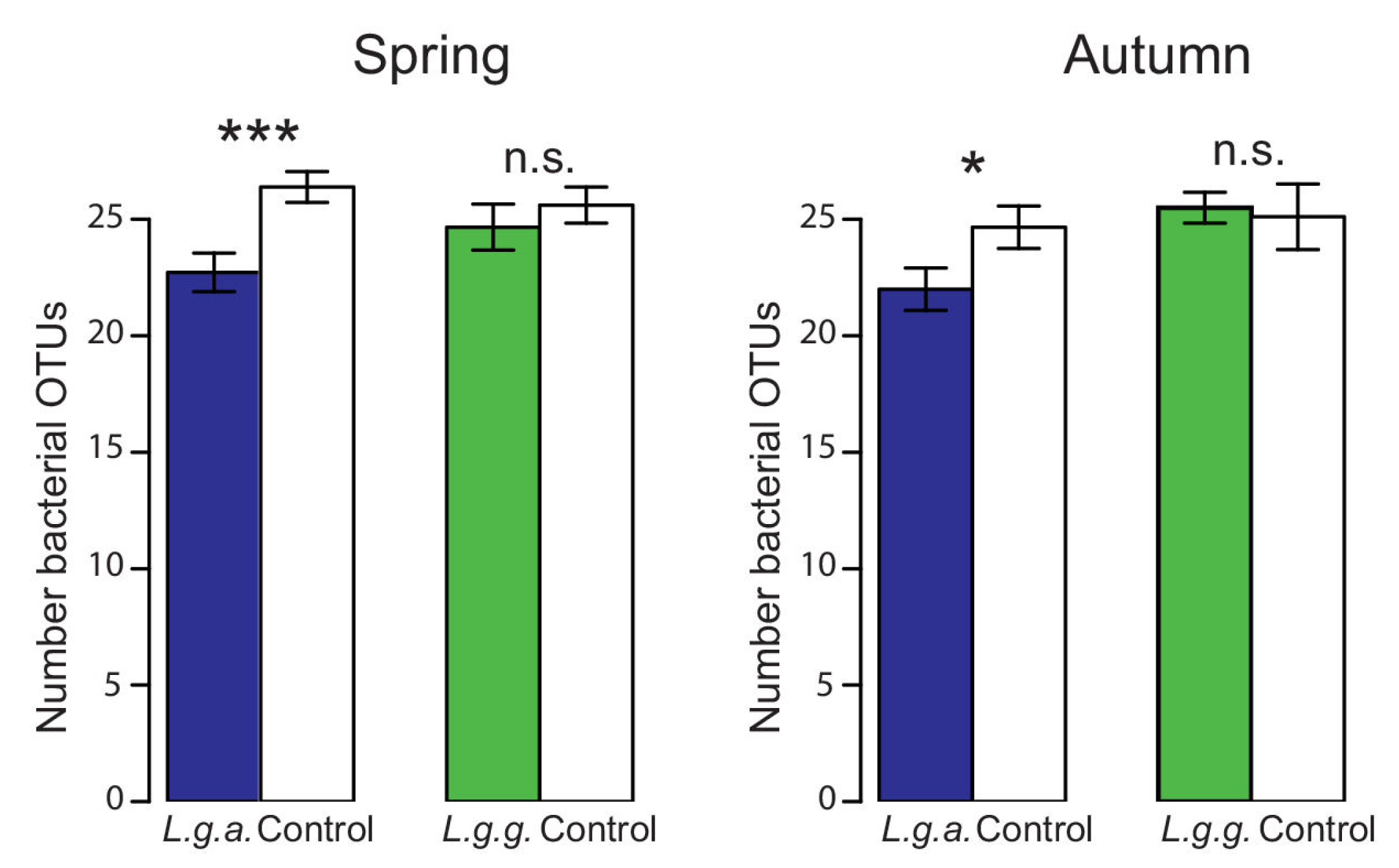
2.5.4. Soil Fungal and Bacterial Community Profiles
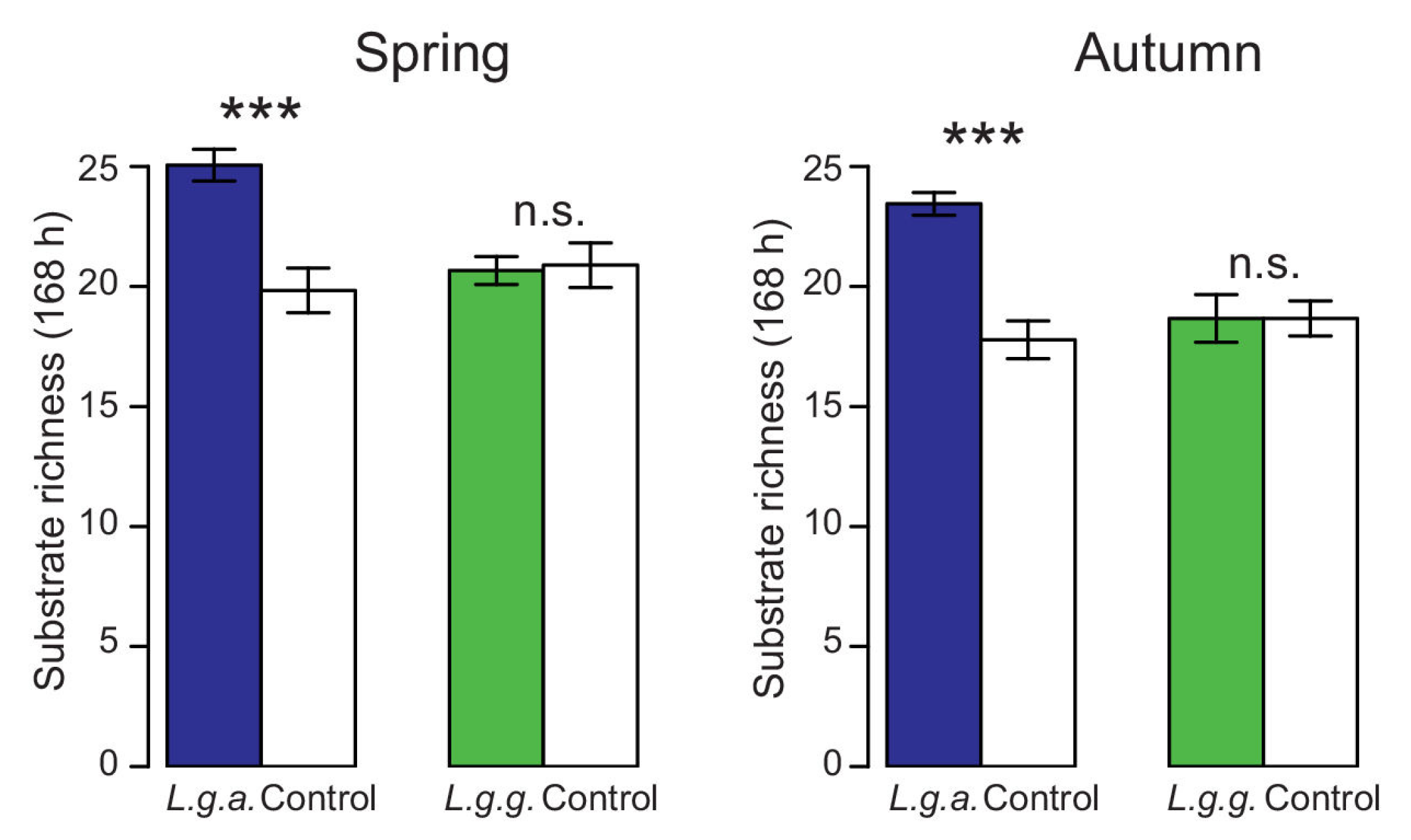
2.5.5. Physiological Profiles of Soil Bacteria
3. Discussion
3.1. Spread of L. g. argentatum in the Region of Basel
3.2. Performance of Lamium subsp. in Swiss Forests
3.3. Performance under Standardized Conditions
3.4. Impact on Native Ground Vegetation and Soil Properties
4. Materials and Methods
4.1. Spread of L. g. argentatum in the Region of Basel
4.2. Performance of Lamium subsp. in Swiss Forests
4.3. Growth of L. g. argentatum and L. g. galeobdolon Patches over 7 Years
4.4. Performance under Standardized Conditions
4.5. Impact on Native Ground Vegetation and Soil Properties
4.5.1. Native Ground Vegetation
4.5.2. Physical and Chemical Soil Properties
4.5.3. Soil Enzyme Activity
4.5.4. Soil Fungal and Bacterial Community Profiles
4.5.5. Physiological Profiles of Soil Bacteria
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Detailed Descriptions of the Methods Used
Appendix A.1. Soil Enzyme Activity (Section 4.5.3)
Appendix A.2. Soil Fungal and Bacterial Community Profiles (Section 4.5.4)
Appendix A.3. Physiological Profiles of Soil Bacteria (Section 4.5.5)
References
- Rejmánek, M.; Richardson, D.M. Trees and shrubs as invasive alien species—2013 update of the global database. Divers. Distrib. 2013, 19, 1093–1094. [Google Scholar] [CrossRef]
- van Kleunen, M.; Essl, F.; Pergl, J.; Brundu, G.; Carboni, M.; Dullinger, S.; Early, R.; González-Moreno, P.; Groom, Q.J.; Hulme, P.E.; et al. The changing role of ornamental horticulture in alien plant invasions. Biol. Rev. 2018, 93, 1421–1437. [Google Scholar] [CrossRef] [PubMed]
- Arianoutsou, M.; Bazos, I.; Christopoulou, A.; Kokkoris, Y.; Zikos, A.; Zervou, S.; Delipetrou, P.; Cardoso, A.C.; Deriu, I.; Gervasini, E.; et al. Alien plants of Europe: Introduction pathways, gateways and time trends. PeerJ 2021, 9, e11270. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.H.; Fitter, A. The characters of successful invaders. Biol. Conserv. 1996, 78, 163–170. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Gómez Aparicio, L.; Haider, S.; Heger, T.; Lortie, C.; Pyšek, P.; Strayer, D.L. Support for major hypotheses in invasion biology is uneven and declining. NeoBiota 2012, 14, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošik, V.; Pergl, J.; Winter, M.; Anastasiu, P.; et al. Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. [Google Scholar]
- Kowarik, I. Biologische Invasionen: Neophyten und Neozoen in Mitteleuropa, 2nd ed.; Eugen Ulmer: Stuttgart, Germany, 2010; p. 492. [Google Scholar]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Pejchar, L.; Mooney, H.A. Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarosik, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vila, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Weidenhamer, J.D.; Callaway, R.M. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J. Chem. Ecol. 2010, 36, 59–69. [Google Scholar] [CrossRef]
- Vogelsang, K.M.; Bever, J.D. Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion. Ecology 2009, 90, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Grove, S.; Haubensak, K.A.; Parker, I.M. Direct and indirect effects of allelopathy in the soil legacy of an exotic plant invasion. Plant Ecol. 2012, 213, 1869–1882. [Google Scholar] [CrossRef]
- Ruckli, R.; Rusterholz, H.-P.; Baur, B. Disrupting ectomycorrhizal symbiosis: Indirect effects of an annual invasive plant on growth and survival of beech (Fagus sylvatica) saplings. Perspect. Plant Ecol. Evol. Syst. 2016, 19, 12–20. [Google Scholar] [CrossRef]
- van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, P.; Kumar Pati, P.; Khan, M.L.; Khare, P.K. Plant functional traits best explain invasive species’ performance within a dynamic ecosystem—A review. Trees For. People 2022, 8, 100260. [Google Scholar] [CrossRef]
- Gallagher, R.V.; Randall, R.P.; Leishman, M.R. Trait differences between naturalized and invasive plant species independent of residence time and phylogeny. Conserv. Biol. 2015, 29, 360–369. [Google Scholar] [CrossRef]
- Hamilton, M.A.; Murray, B.R.; Cadotte, M.W.; Hose, G.C.; Baker, A.C.; Harris, C.J.; Licari, D. Life-history correlates of plant invasiveness at regional and continental scales. Ecol. Lett. 2005, 8, 1066–1074. [Google Scholar] [CrossRef]
- Schmitz, U.; Ristow, M.; May, R.; Bleeker, W. Hybridisierung zwischen Neophyten und heimischen Pflanzenarten in Deutschland—Untersuchungen zur Häufigkeit und zum Gefährdungspotential. Nat. Landsch. 2008, 83, 444–451. [Google Scholar]
- Rosenbaumová, R.; Plačková, I.; Suda, J. Variation in Lamium subg. Galeobdolon (Lamiaceae)—Insights from ploidy levels, morphology and isozymes. Plant Syst. Evol. 2004, 244, 219–244. [Google Scholar] [CrossRef]
- Eber, W. Vergleichende Untersuchungen an Sippen des Lamium galeobdolon agg. mit besonderer Berücksichtigung von L. endtmannii. Florist. Rundbr. 1999, 33, 114–129. [Google Scholar]
- Nehring, S.; Kowarik, I.; Rabitsch, W.; Essel, F. Naturschutzfachliche Invasivitätsbewertungen für in Deutschland wild lebende gebietsfremde Gefäßpflanze. BfN-Skripten 2013, 352, 1–202. [Google Scholar]
- Global Biodiversity Information Facility 2023. Available online: www.gbif.org/species/6411098 (accessed on 15 January 2023).
- Tyler, T.; Karlsson, T.; Milberg, P.; Sahlin, U.; Sundberg, S. Invasive plant species in the Swedish flora: Developing criteria and definitions, and assessing the invasiveness of individual taxa. Nord. J. Bot. 2015, 33, 300–317. [Google Scholar] [CrossRef] [Green Version]
- Plantlife 2023. Available online: www.plantlife.org.uk/uk/discover-wild-plants-nature/plant-fungi-species/variegated-yellow-archangel (accessed on 15 January 2023).
- Invasive Plant Atlas of the United States 2023. Available online: https://www.invasiveplantatlas.org/subject.html?sub=14085 (accessed on 17 January 2023).
- Packham, J.R. Lamiastrum galeobdolon (L.) Ehrend. & Polatschek (Galeobdolon luteum Hudson; Lamium galeobdolon (L.) Nath.). J. Ecol. 1983, 71, 975–997. [Google Scholar] [CrossRef]
- Gaggini, L.; Rusterholz, H.-P.; Baur, B. Settlements as a source for the spread of non-native plants into Central European suburban forests. Acta Oecol. 2017, 79, 18–25. [Google Scholar] [CrossRef]
- Rusterholz, H.-P.; Schneuwly, J.; Baur, B. Invasion of the alien shrub Prunus laurocerasus in suburban deciduous forests: Effects on native vegetation and soil properties. Acta Oecol. 2018, 92, 44–51. [Google Scholar] [CrossRef]
- Rusterholz, H.-P.; Wirz, D.; Baur, B. Garden waste deposits as a source for non-native plants in mixed deciduous forests. Appl. Veg. Sci. 2012, 15, 329–337. [Google Scholar] [CrossRef]
- Brodtbeck, T.; Zemp, M.; Frei, M.; Kienzle, U.; Knecht, D. Flora von Basel und Umgebung 1980–1996. Teil 2. Mitt. Nat. Forsch. Ges. Beider Basel 2007, 3, 545–1003. [Google Scholar]
- Bradley, B.A.; Blumenthal, D.M.; Early, R.; Grosholz, E.D.; Lawler, J.J.; Miller, L.P.; Sorte, C.J.B.; Carla, M.; D’Antonio, C.M.; Diez, J.M.; et al. Global change, global trade, and the next wave of plant invasions. Front. Ecol. Environ. 2012, 10, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Kinlock, N.L.; Dehnen-Schmutz, K.; Essl, F.; Pergl, J.; Pyšek, P.; Kreft, H.; Weigelt, P.; Yang, Q.; van Kleunen, M. Introduction history mediates naturalization and invasiveness of cultivated plants. Glob. Ecol. Biogeogr. 2022, 31, 1101–1119. [Google Scholar] [CrossRef]
- Essl, F.; Milasowszky, N.; Dirnböck, T. Plant invasions in temperate forests: Resistance or ephemeral phenomenon? Basic Appl. Ecol. 2011, 12, 1–9. [Google Scholar] [CrossRef]
- Tyler, T.; Andersson, S.; Fröberg, L.; Olsson, K.-A.; Svensson, Å.; Olsson, A. Recent changes in the frequency of plant species and vegetation types in Scania, S Sweden, compared to changes during the twentieth century. Biodivers. Conserv. 2020, 29, 709–728. [Google Scholar] [CrossRef] [Green Version]
- Online Atlas of the British and Irish Flora 2023. Available online: https://plantatlas.brc.ac.uk/plant/lamiastrum-galeobdolon-subsp-argentatum (accessed on 5 February 2023).
- Walter, E. Die Silber-Goldnessel (Galeobdolon argentatum Smejkal)—Eine verwildernde, im Gelände leicht zu erkennende neue Pflanzensippe. Florist. Rundbr. 1995, 29, 125–128. [Google Scholar]
- Wegmüller, S. A cytotaxonomic study of Lamiastrum galeobdolon (L.) Ehrend. & Polatschek in Britain. Watsonia 1971, 8, 277–288. [Google Scholar]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 97–126. [Google Scholar]
- Rusterholz, H.-P. Reproductive Characteristics of the Horticultural Lamium galeoboldon argentatum and the Native Lamium galeobdolon galeobdolon. University of Basel, Basel, Switzerland. 2019; Unpublished work. [Google Scholar]
- Divíšek, J.; Chytrý, M.; Beckage, B.; Gotelli, N.J.; Lososová, Z.; Pyšek, P.; Richardson, D.M.; Molofsky, J. Similarity of introduced plant species to native ones facilitates naturalization, but differences enhance invasion success. Nat. Commun. 2018, 9, 4631. [Google Scholar] [CrossRef] [Green Version]
- Chapin III, F.S.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Alpert, P. Clonal integration in Fragaria chiloensis differs between populations: Ramets from grassland are selfish. Oecologia 1999, 120, 69–76. [Google Scholar] [CrossRef]
- Suzuki, J.-I.; Stuefer, J. On the ecological and evolutionary significance of storage in clonal plants. Plant Species Biol. 1999, 14, 11–17. [Google Scholar] [CrossRef]
- He, L.; Xiao, X.; Zhang, X.; Jin, Y.; Pu, Z.; Lei, N.; He, X.; Chen, J. Clonal fragments of stoloniferous invasive plants benefit more from stolon storage than their congeneric native species. Flora 2021, 281, 151877. [Google Scholar] [CrossRef]
- Song, Y.B.; Zhou, M.Y.; Dai, W.H.; Jiang, D.; Li, W.B.; Dong, M. Effects of node position on regeneration of stolon fragments in congeneric invasive and native Alternanthera species in China. Plant Species Biol. 2014, 29, E93–E100. [Google Scholar] [CrossRef]
- Song, Y.B.; Yu, F.H.; Li, J.M.; Keser, L.H.; Fischer, M.; Dong, M.; van Kleunen, M. Plant invasiveness is not linked to the capacity of regeneration from small fragments: An experimental test with 39 stoloniferous species. Biol. Invasions 2013, 15, 1367–1376. [Google Scholar] [CrossRef] [Green Version]
- Martínková, J.; Klimešová, J. Enforced clonality confers a fitness advantage. Front. Plant Sci. 2016, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, K.M.; McCarthy, B.C. Changes in forest structure and species composition following invasion by a non-indigenous shrub, Amur honeysuckle (Lonicera maackii). J. Tor. Bot. Soc. 2008, 135, 245–259. [Google Scholar] [CrossRef]
- Kumar, M.; Verma, A.K.; Garkoti, S.C. Lantana camara and Ageratina adenophora invasion alter the understory species composition and diversity of chir pine forest in central Himalaya, India. Acta Oecol. 2020, 109, 103642. [Google Scholar] [CrossRef]
- Thiele, J.; Isermann, M.; Otte, A.; Kollmann, J. Competitive displacement or biotic resistance? Disentangling relationships between community diversity and invasion success of tall herbs and shrubs. J. Veg. Sci. 2010, 21, 213–220. [Google Scholar] [CrossRef]
- Scharfy, D.; Güsewell, S.; Gessner, M.O.; Venterink, H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: Effects on soil microbes, nutrients and plant-soil feedbacks. J. Ecol. 2010, 98, 1379–1388. [Google Scholar] [CrossRef]
- Stoll, P.; Gatzsch, K.; Rusterholz, H.-P.; Baur, B. Response of plant and gastropod species to knotweed invasion. Basic Appl. Ecol. 2012, 13, 232–240. [Google Scholar] [CrossRef]
- Buchholz, S.; Tietze, H.; Kowarik, I.; Schirmel, J. Effects of a major tree invader on urban woodland arthropods. PLoS ONE 2015, 10, e0137723. [Google Scholar] [CrossRef] [Green Version]
- Collier, M.H.; Vankat, J.L.; Hughes, M.R. Diminished plant richness and abundance below Lonicera maackii, an invasive shrub. Am. Midl. Nat. 2002, 147, 60–71. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Klironomos, J.N.; Wardle, D.A. Microbial ecology of biological invasions. ISME J. 2007, 1, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Ehrenfeld, J.G. Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef] [Green Version]
- Unger, I.M.; Kremer, R.J.; Veum, K.S.; Goyne, K.W. Immediate and long-term effects of invasive plant species on soil characteristics. Soil Ecol. Lett. 2022, 4, 276–288. [Google Scholar] [CrossRef]
- Heneghan, L.; Fatemi, F.; Umek, L.; Grady, K.; Fagen, K.; Workman, M. The invasive shrub European buckthorn (Rhamnus cathartica, L.) alters soil properties in Midwestern U.S. woodlands. Appl. Soil Ecol. 2006, 32, 142–148. [Google Scholar] [CrossRef]
- Martin, M.R.; Tipping, P.W.; Sickman, J.O. Invasion by an exotic tree alters above and belowground ecosystem components. Biol. Invasions 2009, 11, 1883–1894. [Google Scholar] [CrossRef]
- Qin, Z.; Xie, J.F.; Quan, G.M.; Zhang, J.E.; Mao, D.J.; DiTommaso, A. Impacts of the invasive annual herb Ambrosia artemisiifolia L. on soil microbial carbon source utilization and enzymatic activities. Europ. J. Soil Biol. 2014, 60, 58–66. [Google Scholar] [CrossRef]
- Hu, C.C.; Lei, Y.B.; Y Tan, Y.H.; Sun, X.C.; Xu, H.; Liu, C.Q.; Liu, X.Y. Plant nitrogen and phosphorus utilization under invasive pressure in a montane ecosystem of tropical China. J. Ecol. 2019, 107, 372–386. [Google Scholar] [CrossRef]
- Burns, R.G.; Dick, R.P. Enzymes in the Environment: Activity, Ecology, and Applications; Marcel Dekker Inc.: New York, NY, USA, 2002; p. 640. [Google Scholar] [CrossRef]
- Zhou, Y.; Staver, A.C. Enhanced activity of soil nutrient-releasing enzymes after plant invasion: A meta-analysis. Ecology 2019, 100, e02830. [Google Scholar] [CrossRef]
- Craig, M.E.; Fraterrigo, J.M. Plant-microbial competition for nitrogen increases microbial activities and carbon loss in invaded soils. Oecologia 2017, 184, 583–596. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Häggblom, M. Exotic plant species alter the microbial community structure and function in the soil. Ecology 2002, 83, 3152–3166. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; Wei, W.; Bai, X.; Wu, N.; Hou, Y. Effects of invasive Rhus typhina L. on bacterial diversity and community composition in soil. Écoscience 2020, 27, 177–184. [Google Scholar] [CrossRef]
- Klimek, B.; Jaźwa, M.; Gołębiewski, M.; Sikora, M.; Deja-Sikora, E. No apparent effect of invasive alien goldenrod on soil microbial communities or soil fauna feeding activity. Acta Oecol. 2020, 109, 103669. [Google Scholar] [CrossRef]
- Burke, D.J.; Chan, C.R. Effects of the invasive plant garlic mustard (Alliaria petiolata) on bacterial communities in a northern hardwood forest soil. Can. J. Microbiol. 2010, 56, 81–86. [Google Scholar] [CrossRef]
- Torres, N.; Herrera, I.; Fajardo, L.; Bustamante, R.O. Meta-analysis of the impact of plant invasion on soil microbial community. BMC Ecol. Evo. 2021, 21, 172. [Google Scholar] [CrossRef]
- Piper, C.L.; Siciliano, S.D.; Winsley, T.; Lamb, E.G. Smooth brome invasion increases rare soil bacterial species prevalence, bacterial species richness and evenness. J. Ecol. 2015, 103, 386–396. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Yee, K.; Johnson, E.A.; Maraun, M.; Parkinson, D.; Straube, D.; Scheu, S. Positive relationship between herbaceous layer diversity and the performance of soil biota in a temperate forest. Soil Biol. Biochem. 2011, 43, 462–465. [Google Scholar] [CrossRef]
- Klimek, B.; Niklińska, M.; Jaźwa, M.; Tarasek, A.; Tekielak, I.; Musielok, L. Covariation of soil bacteria functional diversity and vegetation diversity along an altitudinal climatic gradient in the Western Carpathians. Pedobiologia 2015, 58, 105–112. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Stefanowicz, A.M.; Kapusta, P.; Stanek, M.; Frąc, M.; Oszust, K.; Woch, M.W.; Zubek, S. Invasive plant Reynoutria japonica produces large amounts of phenolic compounds and reduces the biomass but not activity of soil microbial communities. Sci. Total Environ. 2021, 767, 145439. [Google Scholar] [CrossRef]
- Wei, H.; Yan, W.; Quan, G.; Zhang, J.; Liang, K. Soil microbial carbon utilization, enzyme activities and nutrient availability responses to Bidens pilosa and a non-invasive congener under different irradiances. Sci. Rep. 2017, 7, 11309. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Dai, T.; Quan, G.; Zhang, J. Changes in soil physico-chemical properties, enzyme activities and soil microbial communities under Mimosa pudica invasion. Allel. J. 2015, 36, 15–23. [Google Scholar]
- Zhang, C.B.; Wang, J.; Qian, B.Y.; Li, W.H. Effects of the invader Solidago canadensis on soil properties. Appl. Soil Ecol. 2009, 43, 163–169. [Google Scholar] [CrossRef]
- Ricciardi, A.; Hoopes, M.F.; Marchetti, M.P.; Lockwood, J.L. Progress toward understanding the ecological impacts of nonnative species. Ecol. Monogr. 2013, 83, 263–282. [Google Scholar] [CrossRef] [Green Version]
- Info Flora Schweiz 2022. Available online: www.infoflora.ch/de/flora/lamium-galeobdolon-subsp-argentatum.html (accessed on 20 October 2022).
- Burnand, J.; Hasspacher, B. Waldstandorte beider Basel; Verlag des Kantons Basel-Landschaft: Liestal, Switzerland, 1999; p. 266. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Blackburn Press: Caldwell, NJ, USA, 2002; p. 547. [Google Scholar]
- Lauber, K.; Wagner, G.; Gygax, A. Flora Helvetica, 5th ed.; Haupt: Bern, Switzerland, 2012; p. 1686. [Google Scholar]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific: Oxford, UK, 1989; p. 368. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis, Part 3, Chemical Methods, 3rd ed.; ASA: Madison, WI, USA, 1996; p. 1424. [Google Scholar]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Thies, J.E. Soil microbial community analysis using terminal restriction fragment length polymorphisms. Soil Sci. Soc. Am. J. 2007, 71, 579–591. [Google Scholar] [CrossRef]
- Zak, J.C.; Willig, M.R.; Moorhead, D.L.; Wildman, H.G. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem. 1994, 26, 1101–1108. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 21 October 2022).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, R. Development Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. 2012, R Package Version 3.1. Available online: http://www.R-project.org/ (accessed on 21 October 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, R.; Stevens, M.H.H.; Wagner, H. Development Core Team. Vegan Community Package. 2019, R Package Version 3.6.3. Available online: http://www.R-project.org/ (accessed on 12 January 2023).
- Clarke, K.R.; Green, R.H. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Braschler, B.; Zwahlen, V.; Gilgado, J.D.; Rusterholz, H.P.; Baur, B. Owners’ perceptions do not match actual ground-dwelling invertebrate diversity in their gardens. Diversity 2021, 13, 189. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, biodiversity, and conservation. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- German, D.P.; Chacon, S.S.; Allison, S.D. Substrate concentration and enzyme allocation can affect rates of microbial decomposition. Ecology 2011, 92, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Arafat, Y.; Wu, L.; Xiao, Z.; Li, Q.; Kham, M.A.; Khan, M.U.; Lin, S.; Lin, W. Shifts in soil microbial community, soil enzymes and crop yield under peanut/maize intercropping with reduced nitrogen levels. Appl. Soil Ecol. 2018, 124, 327–334. [Google Scholar] [CrossRef]
- Li, F.; Hullar, M.A.J.; Lampe, J.W. Optimization of terminal restriction fragment polymorphism (TRFLP) analysis of human gut microbiota. J. Microbiol. Methods 2007, 68, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Barto, E.K.; Antunes, P.M.; Stinson, K.A.; Koch, A.M.; Klironomos, J.N.; Cipollini, D. Differences in arbuscular mycorrhizal fungal communities associated with sugar maple seedlings in and outside of invaded garlic mustard forest patches. Biol. Invasions 2011, 13, 2755–2762. [Google Scholar] [CrossRef]
- Kissling, M.; Hegetschweiler, K.T.; Rusterholz, H.-P.; Baur, B. Short-term and long-term effects of human trampling on above-ground vegetation, soil density, soil organic matter and soil microbial processes in suburban beech forests. Appl. Soil Ecol. 2009, 42, 303–314. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates N/E | Subspecies | Patch Size (m2) 1 | Plant Cover (%) 1 | Number of Inflorescences 2 | Number of Stolons 2 | Stolon Length (cm) 2 | Number of Nodes 2 |
|---|---|---|---|---|---|---|---|---|
| Allschwil | 47.53119/7.54510 | Horticultural | 57.5 (7.5–450.0) | 77.5 (60–90) | 0.5 (0–4) | 3.5 (2–5) | 79.5 (58.0–117.0) | 9 (5–14) |
| Native | 4.5 (1.0–10.0) | 67.5 (50–80) | 0.5 (0–2) | 3 (1–5) | 57.5 (40.0–111.0) | 6.5 (3–12) | ||
| Bottmingen | 47.50928/7.56916 | Horticultural | 11.0 (5.0–250.0) | 77.5 (60–80) | 1.5 (0–4) | 3.5 (1–5) | 56.5 (41.0–107.0) | 7 (5–11) |
| Native | 3.5 (2.0–7.5) | 50.0 (50–75) | 1.5 (0–3) | 3 (1–5) | 60.0 (35.0–112.0) | 6.5 (3–12) | ||
| Riehen | 47.57129/7.65276 | Horticultural | 100.0 (15.0–500.0) | 90.0 (50–100) | 1.0 (0–2) | 3 (3–5) | 86.5 (47.0–125.0) | 7 (5–11) |
| Native | 5.0 (2.0–10.0) | 70.5 (50–80) | 1.0 (0–2) | 3 (2–3) | 50.0 (42.0–97.0) | 5.5 (4–7) | ||
| Zell (Ger) | 47.69662/7.85479 | Horticultural | 12.5 (5.0–650.0) | 75 (60–80) | 0.0 (0–3) | 4 (1–6) | 59.0 (40.0–101.0) | 5 (5–7) |
| Native | 5.0 (3.0–10.0) | 68 (50–90) | 1.0 (0–3) | 2 (1–3) | 50.5 (47.0–69.0) | 6 (4–9) | ||
| Seprais | 47.36978/7.22973 | Horticultural | 10.5 (5.0–12.0) | 63 (40–75) | 0.0 (0–0) | 3 (2–5) | 70.0 (52.0–91.0) | 9 (5–11) |
| Native | 2.0 (1.0–4.0) | 75 (50–80) | 1.0 (0–2) | 3 (1–4) | 52.0 (35.0–61.0) | 4.5 (3–6) | ||
| Allaman | 46.46913/6.41164 | Horticultural | 155.0 (18.0–550.0) | 55 (40–70) | 1.0 (0–2) | 4 (2–5) | 72.5 (48.0–101.0) | 4 (3–6) |
| Native | 3.5 (1.0 –10.0) | 90 (50–90) | 1.0 (0–2) | 2 (1–4) | 62.0 (21.0–107.0) | 6 (3–8) | ||
| Köniz | 46.93223/7.41164 | Horticultural | 10.0 (2.0–25.0) | 75 (50–100) | 2.0 (0–4) | 3 (3–5) | 91.5 (50.0–101.0) | 6 (4–9) |
| Native | 5.5 (2.0–9.0) | 65 (10–75) | 2.0 (0–3) | 2 (2–5) | 66.5 (41.0–121.0) | 5 (4–9) | ||
| Oberwangen | 46.92649/7.46436 | Horticultural | 10.0 (4.0–12.0) | 75 (50–80) | 1.0 (0–2) | 3 (2–5) | 61.0 (41.0–101.0) | 9.5 (7–11) |
| Native | 5.0 (4.0–12.0) | 75 (10–80) | 1.0 (0–2) | 3 (1–4) | 66.5 (41.0–141.0) | 6.5 (4–11) | ||
| Wabern | 46.92649/7.46436 | Horticultural | 9.0 (6.0–25.0) | 77.5 (60–90) | 0.0 (0–2) | 3 (3–5) | 90.5 (51.0–123.0) | 6 (4–9) |
| Native | 4.0 (2.0–8.0) | 70.5 (40–90) | 0.5 (0–2) | 3 (1–4) | 57.5 (43.0–96.0) | 6 (3–11) | ||
| Hägglingen | 47.37990/8.26482 | Horticultural | 57.5 (15.0–325.0) | 82.5 (75–90) | 1.0 (0–2) | 3 (2–4) | 51.5 (34.0–118.0) | 5 (3–13) |
| Native | 13.5 (6.0–100.0) | 72.5 (60–90) | 1.0 (0–2) | 2 (1–3) | 56.0 (45.0–112.0) | 6 (4–11) | ||
| Zürich | 47.38537/8.57158 | Horticultural | 11.0 (6.0–22.5) | 75 (60–90) | 1.0 (0–2) | 4 (2–5) | 63.5 (41.0–109.0) | 7 (4–11) |
| Native | 5.5 (2.0–9.0) | 90 (60–100) | 0.5 (0–4) | 3 (2–4) | 56.0 (28.0–128.0) | 7 (5–11) | ||
| Zollikon | 47.34246/8.58590 | Horticultural | 11.0 (6.0–36.0) | 77.5 (60–90) | 0.0 (0–0) | 3 (2–5) | 70.0 (47.0–107.0) | 7 (5–8) |
| Native | 10.0 (4.0–30.0) | 75 (60–90) | 1.0 (0–4) | 2.5 (1–5) | 70.5 (58.0–105.0) | 6 (3–10) |
| Plant Species Richness | Plant-Available Phosphorus 1 | Soil Moisture | Acid Phosphatase Activity 2 | Number of Bacterial OTUs | AWCD (168 h) | Substrate Richness (168 h) | |
|---|---|---|---|---|---|---|---|
| Subspecies | F1,30 = 76.29, p < 0.001 | F1,30 = 8.16, p = 0.006 | F1,30 = 0.66, p = 0.422 | F1,30 = 12.86, p = 0.001 | F1,30 = 7.76, p = 0.001 | F1,30 = 2.76, p = 0.107 | F1,30 = 22.72, p < 0.001 |
| Forest | F2,30 = 0.49, p = 0.619 | F2,30 = 0.99, p = 0.384 | F2,30 = 1.25, p = 0.300 | F2,30 = 2.25, p = 0.123 | F2,30 = 1.31, p = 0.285 | F2,30 = 0.78, p = 0.467 | F2,30 = 1.80, p = 0.183 |
| Season | F1,30 = 34.91, p < 0.001 | F1,30 = 3.95, p = 0.056 | F1,30 = 1.95, p = 0.173 | F1,30 = 15.52, p = 0.001 | F1,30 =1.35, p = 0.254 | F1,30 = 0.77, p = 0.386 | F1,30 = 0.12, p = 0.729 |
| Subspecies × Forest | F2,30 = 0.84, p = 0.442 | F2,30 = 0.34, p = 0.716 | F2,30 = 1.05, p = 0.365 | F2,30 = 1.81, p = 0.182 | F2,30 = 4.64, p = 0.018 | F2,30 = 0.46, p = 0.638 | F2,30 = 0.74, p = 0.486 |
| Subspecies × Season | F1,30 = 6.99, p = 0.013 | F1,30 = 0.14, p = 0.714 | F1,30 = 4.85, p = 0.035 | F1,30 = 31.42, p < 0.001 | F1,30 = 0.03, p = 0.869 | F1,30 = 0.65, p = 0.421 | F1,30 = 0.01, p = 0.908 |
| Subspecies × Forest × Season | F4,30 = 1.60, p = 0.199 | F4,30 = 1.73, p = 0.170 | F4,30 = 0.42, p = 0.796 | F4,30 = 2.57, p = 0.058 | F4,30 = 5.48, p = 0.002 | F4,30 = 1.37, p = 0.269 | F4,30 = 2.08, p = 0.109 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusterholz, H.-P.; Huber, K.; Baur, B. Invasion of a Horticultural Plant into Forests: Lamium galeobdolon argentatum Affects Native Above-Ground Vegetation and Soil Properties. Plants 2023, 12, 1527. https://doi.org/10.3390/plants12071527
Rusterholz H-P, Huber K, Baur B. Invasion of a Horticultural Plant into Forests: Lamium galeobdolon argentatum Affects Native Above-Ground Vegetation and Soil Properties. Plants. 2023; 12(7):1527. https://doi.org/10.3390/plants12071527
Chicago/Turabian StyleRusterholz, Hans-Peter, Katharina Huber, and Bruno Baur. 2023. "Invasion of a Horticultural Plant into Forests: Lamium galeobdolon argentatum Affects Native Above-Ground Vegetation and Soil Properties" Plants 12, no. 7: 1527. https://doi.org/10.3390/plants12071527