

Effects of White and Blue-Red Light on Growth and Metabolism of Basil Grown under Microcosm Conditions
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
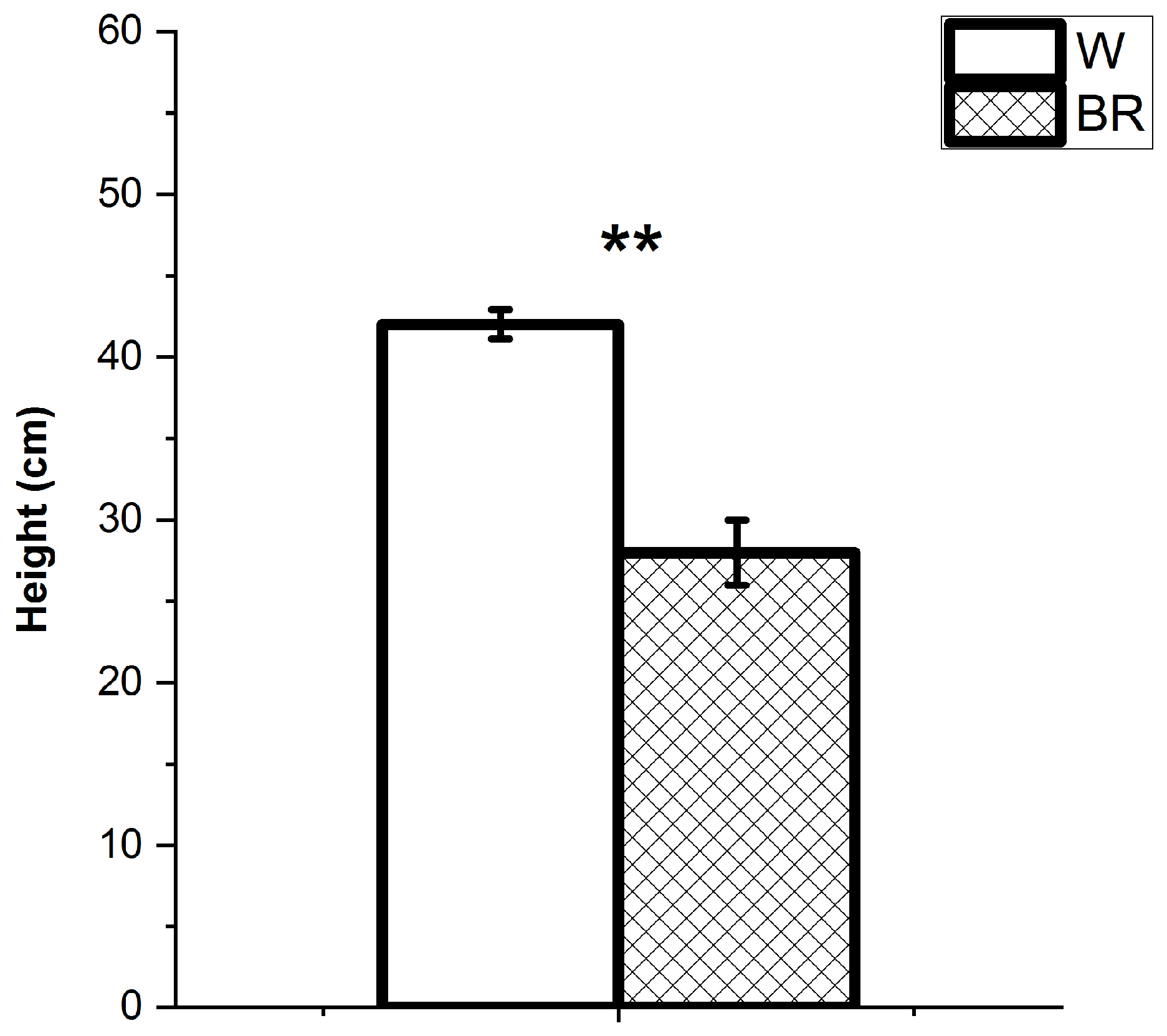
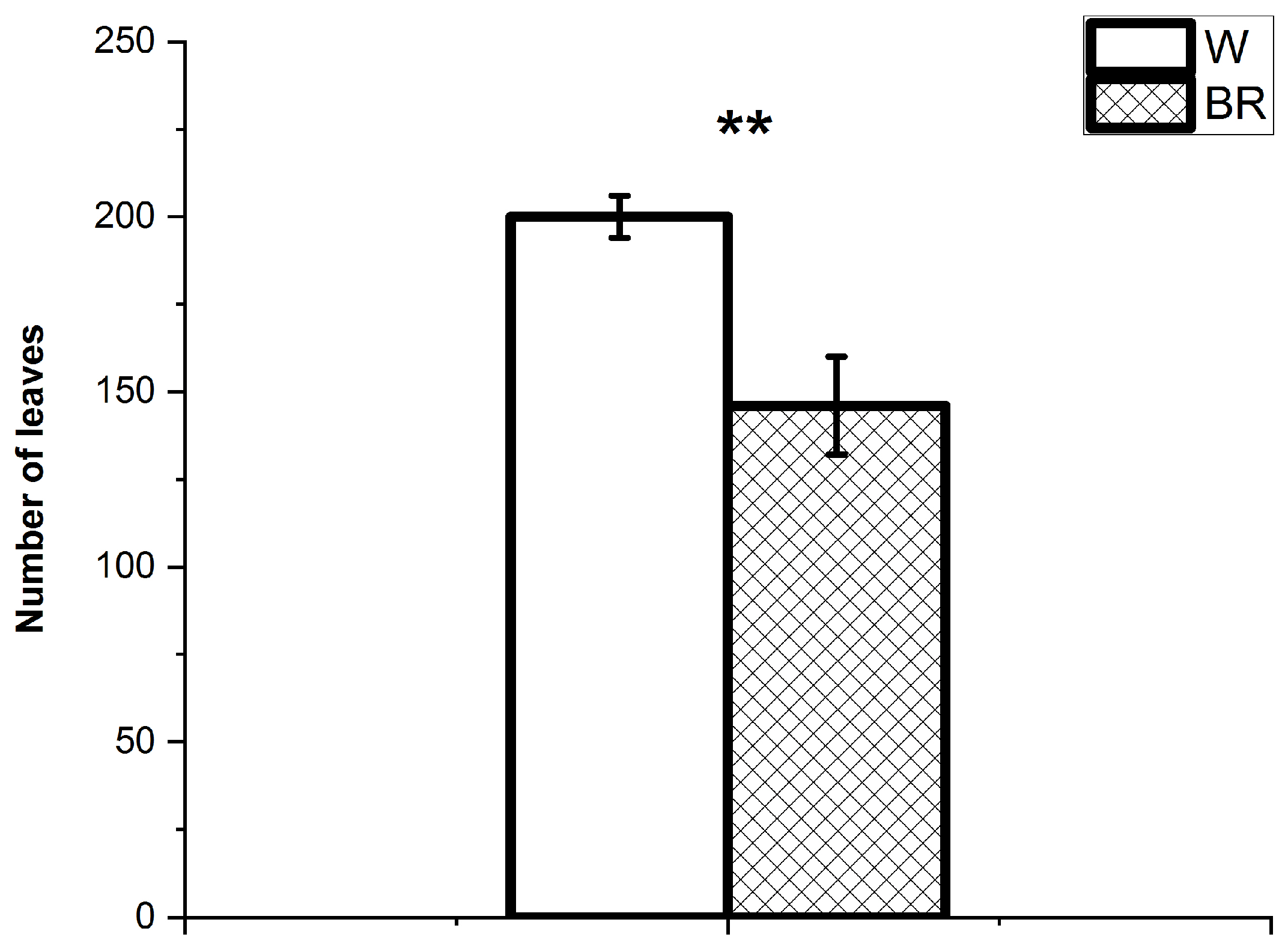
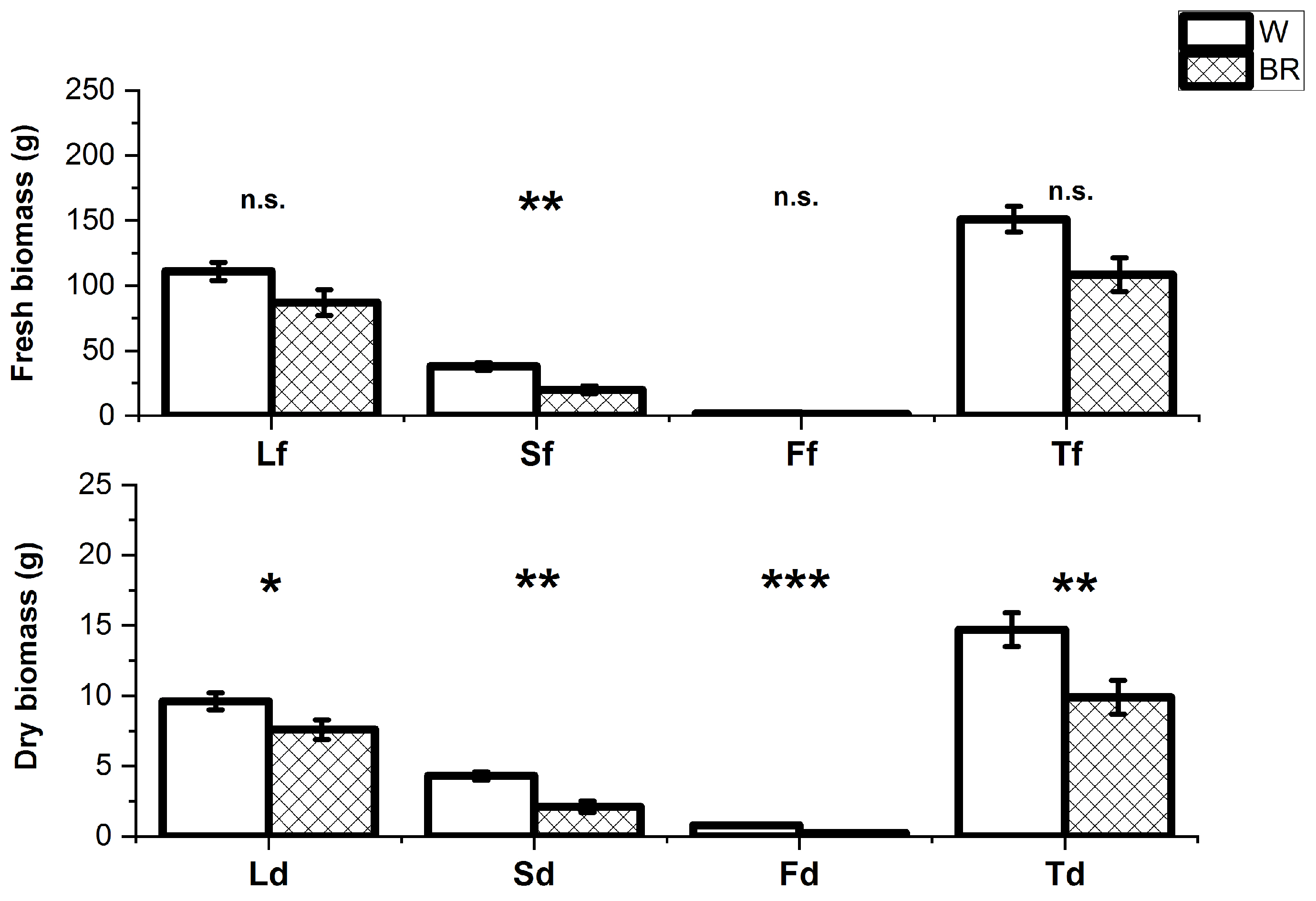
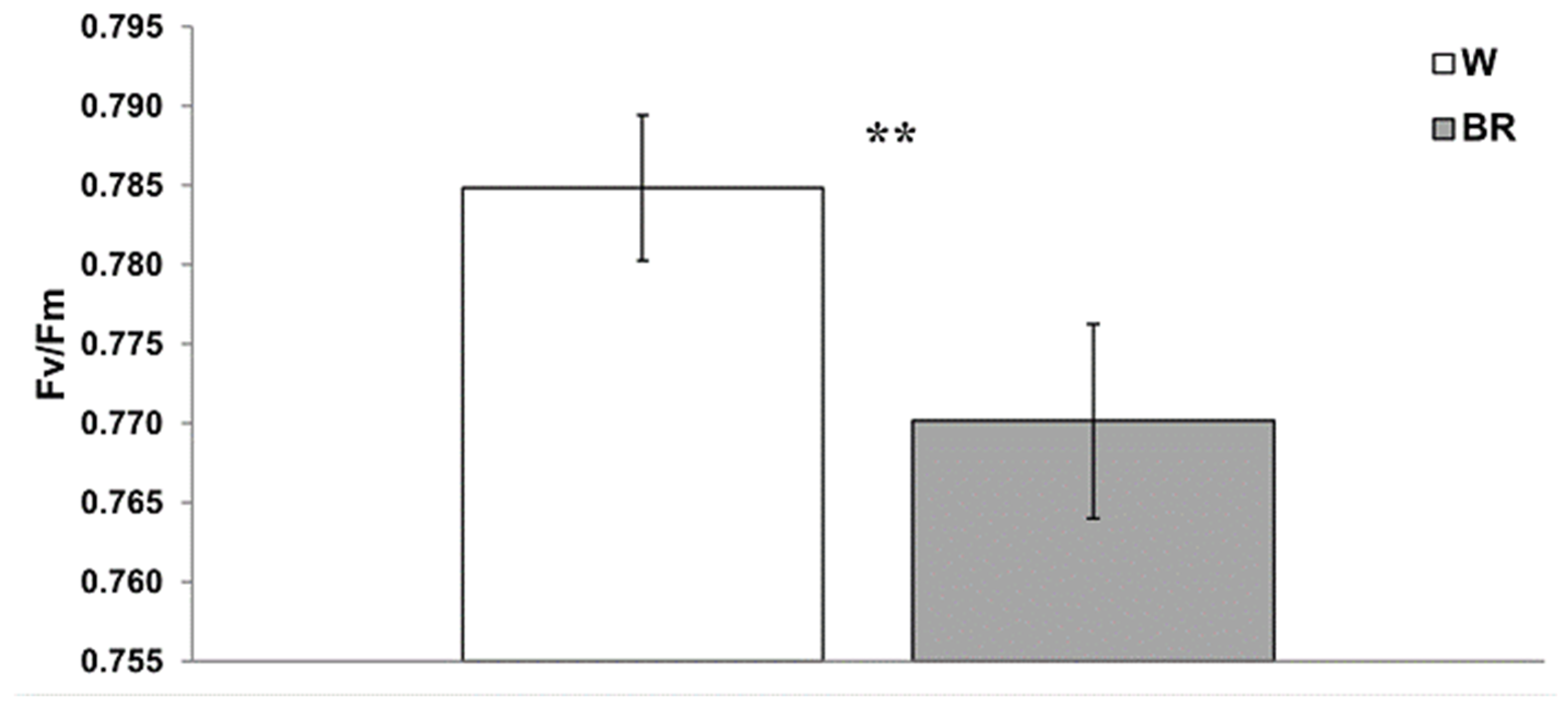
2. Results
2.1. Plant Growth and Development
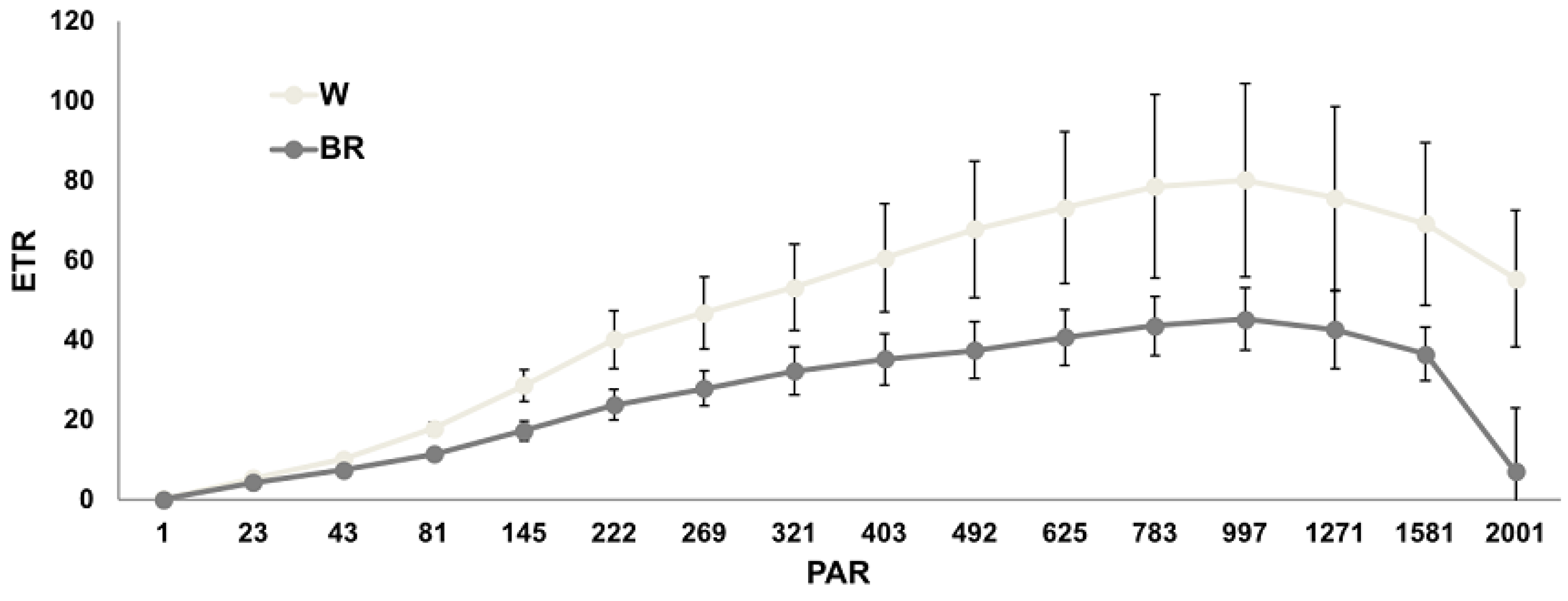
2.2. Chlorophyll Fluorescence Analysis
2.3. Nutrient Concentration
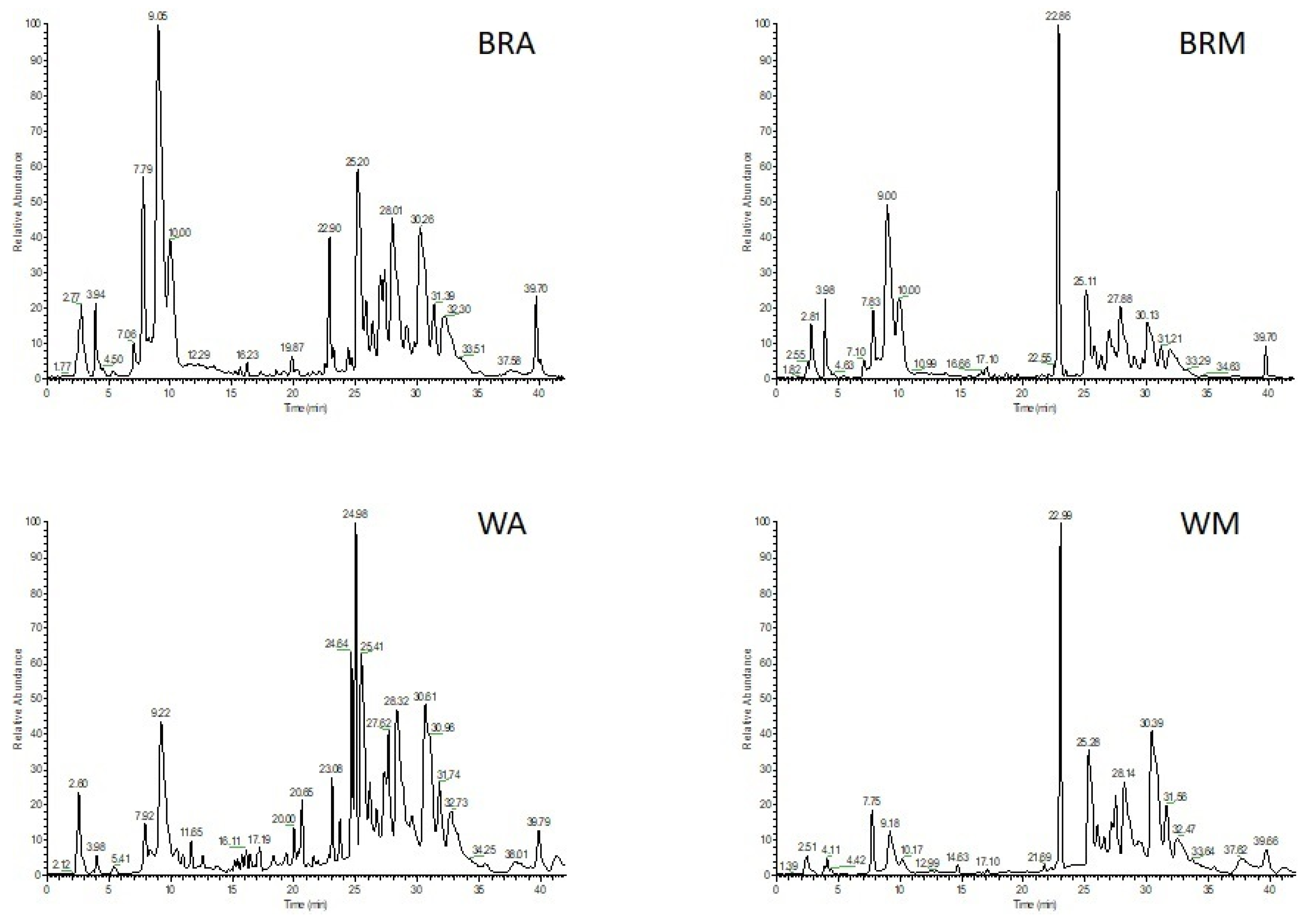
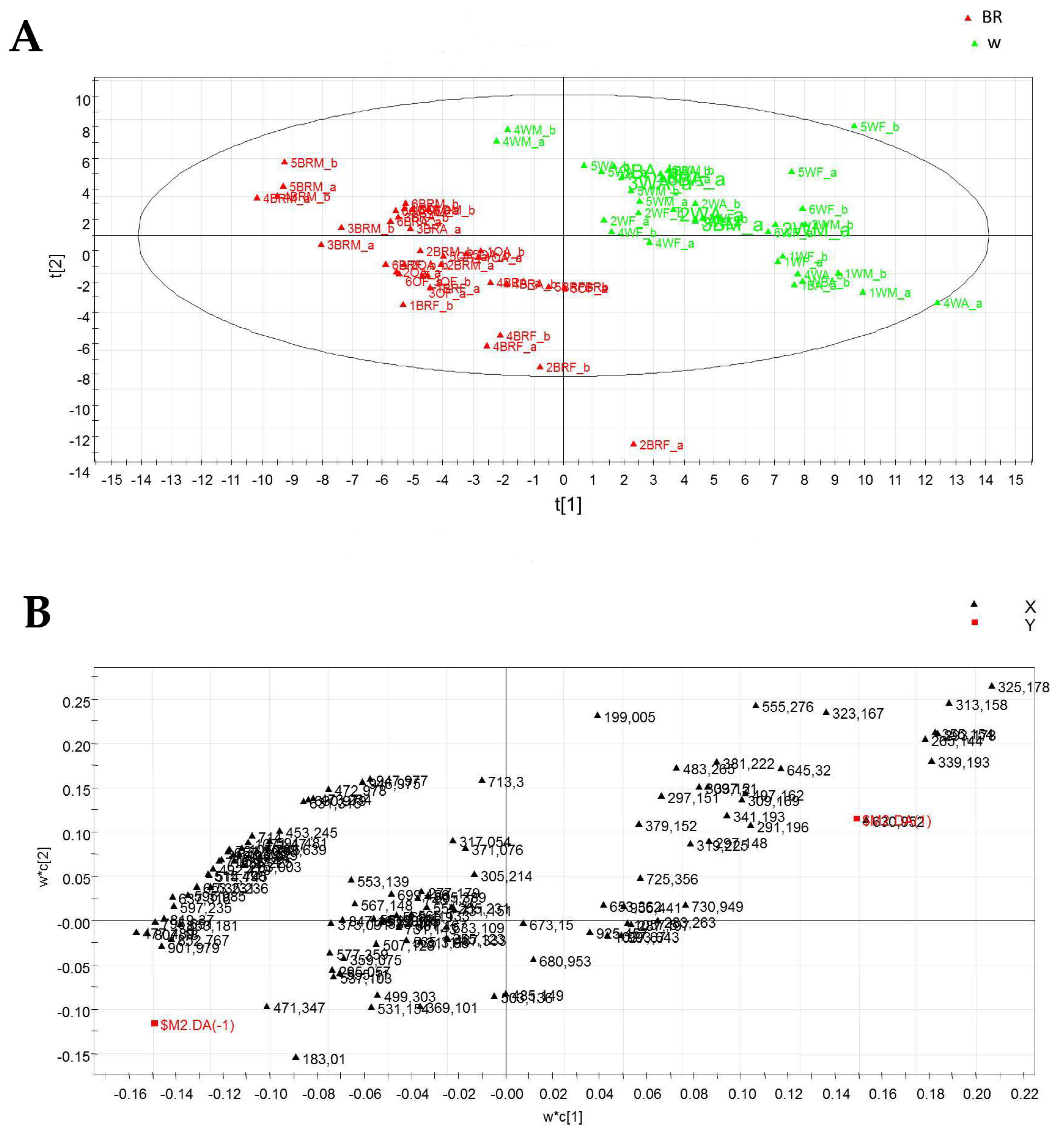
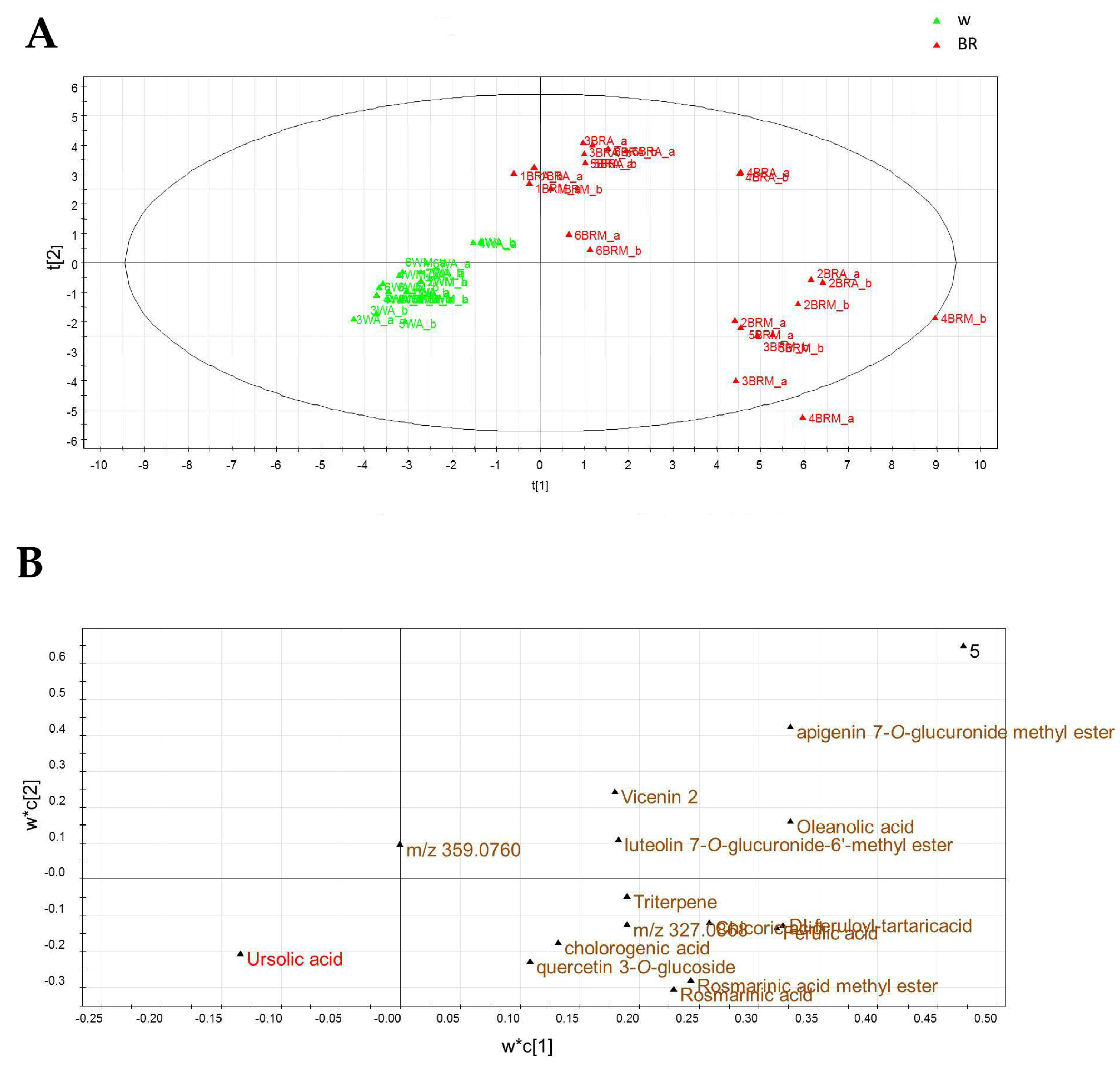
2.4. Metabolic Profiling
3. Discussion
4. Materials and Methods
4.1. Germplasm and Growing Conditions
4.2. Biometric Determinations
4.3. Chlorophyll Fluorescence Analysis
4.4. Ion Chromatography Analysis
4.5. Plant Extraction and Hyphenated Liquid Chromatography High-Resolution Mass Spectrometry (LC-ESI-Orbitrap-MS) Analysis
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zabel, P.; Bamsey, M.; Schubert, D.; Tajmar, M. Review and analysis of plant growth chambers and greenhouse modules for space. In Proceedings of the 44th International Conference on Environmental Systems ICES-2014-120, Tucson, AZ, USA, 13–17 July 2014. [Google Scholar]
- Abdullah, M.J.; Zhang, Z.; Matsubae, K. Potential for Food Self-Sufficiency Improvements through Indoor and Vertical Farming in the Gulf Cooperation Council: Challenges and Opportunities from the Case of Kuwait. Sustainability 2021, 13, 12553. [Google Scholar] [CrossRef]
- Benke, K.; Tomkins, B. Future food-production systems: Vertical farming and controlled-environment agriculture. Sustain. Sci. Pract. Policy 2017, 13, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Goto, E. Plant production in a closed plant factory with artificial lighting. Acta Hortic. 2012, 956, 37–50. [Google Scholar] [CrossRef]
- Kozai, T.; Ohyama, K.; Chun, C. Commercialized closed systems with artificial lighting for plant production. Acta Hortic. 2006, 711, 61–70. [Google Scholar] [CrossRef]
- SharathKumar, M.; Heuvelink, E.; Marcelis, L.F.M. Vertical farming: Moving from genetic to environmental modification. Trends Plant Sci. 2020, 25, 724–727. [Google Scholar] [CrossRef]
- Wong, C.E.; Teo, Z.W.N.; Shen, L.; Yu, H. Seeing the lights for leafy greens in indoor vertical farming. Trends Food Sci. Technol. 2020, 106, 48–63. [Google Scholar] [CrossRef]
- Avercheva, O.V.; Berkovich, Y.A.; Konovalova, I.O.; Radchenko, S.G.; Lapach, S.N.; Bassarskaya, E.M.; Kochetova, G.V.; Zhigalova, T.V.; Yakovleva, O.S.; Tarakanov, I.G. Optimizing LED lighting for space plant growth unit: Joint effects of photon flux density, red to white ratios and intermittent light pulses. Life Sci. Space Res. 2016, 11, 29–42. [Google Scholar] [CrossRef]
- Bula, R.J.; Morrow, R.C.; Tibbitts, T.W.; Barta, D.J.; Ignatius, R.W.; Martin, T.S. Light-emitting Diodes as a Radiation Source for Plants. HortScience 1991, 26, 203–205. [Google Scholar] [CrossRef] [Green Version]
- Morrow, R.C. Led Lighting in Horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for energy efficient greenhouse lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Viršilė, A.; Olle, M.; Duchovskis, P. LED Lighting in Horticulture. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 113–147. [Google Scholar] [CrossRef]
- Zhang, H.; Burr, J.; Zhao, F. A comparative life cycle assessment (LCA) of lighting technologies for greenhouse crop production. J. Clean. Prod. 2017, 140, 705–713. [Google Scholar] [CrossRef]
- Chutimanukul, P.; Wanichananan, P.; Janta, S.; Toojinda, T.; Darwell, C.T.; Mosaleeyanon, K. The influence of different light spectra on physiological responses, antioxidant capacity and chemical compositions in two holy basil cultivars. Sci. Rep. 2022, 12, 588. [Google Scholar] [CrossRef]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Phil. Trans. R. Soc. B 2018, 369, 20130243. [Google Scholar] [CrossRef] [Green Version]
- Folta, K.M.; Childers, K.S. Light as a Growth Regulator: Controlling Plant Biology with Narrow-bandwidth Solid-state Lighting Systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef] [Green Version]
- Hammock, H.A.; Kopsell, D.A.; Sams, C.E. Narrowband Blue and Red LED Supplements Impact Key Flavor Volatiles in Hydroponically Grown Basil Across Growing Seasons. Front. Plant Sci. 2021, 12, 623314. [Google Scholar] [CrossRef]
- Larsen, D.H.; Woltering, E.J.; Nicole, C.C.S.; Marcelis, L.F.M. Response of Basil Growth and Morphology to Light Intensity and Spectrum in a Vertical Farm. Front. Plant Sci. 2020, 11, 597906. [Google Scholar] [CrossRef]
- Massa, G.D.; Kim, H.; Wheeler, R.M.; Mitchell, C.A. Plant Productivity in Response to LED Lighting. HortScience 2008, 43, 1951–1956. [Google Scholar] [CrossRef]
- Olle, M.; Viršilė, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, L.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the Role of Red:Blue LED Lights on Resource Use Efficiency and Nutritional Properties of Indoor Grown Sweet Basil. Front. Plant Sci. 2019, 10, 305. [Google Scholar] [CrossRef] [Green Version]
- Sipos, L.; Balázs, L.; Székely, G.; Jung, A.; Sárosi, S.; Radácsi, P.; Csambalik, L. Optimization of basil (Ocimum basilicum L.) production in LED light environments—A review. Sci Hortic. 2021, 289, 110486. [Google Scholar] [CrossRef]
- Nazir, M.; Tungmunnithum, D.; Bose, S.; Drouet, S.; Garros, L.; Giglioli-Guivarc’h, N.; Abbasi, B.H.; Hano, C. Differential Production of Phenylpropanoid Metabolites in Callus Cultures of Ocimum basilicum L. with Distinct in Vitro Antioxidant Activities and in Vivo Protective Effects against UV Stress. J. Agric. Food Chem. 2019, 67, 1847–1859. [Google Scholar] [CrossRef] [PubMed]
- Jakovljević, D.; Stanković, M.; Warchoł, M.; Skrzypek, E. Basil (Ocimum L.) cell and organ culture for the secondary metabolites production: A review. Plant Cell Tissue Organ Cult. 2022, 149, 61–79. [Google Scholar] [CrossRef]
- Johnson, A.J.; Meyerson, E.; de la Parra, J.; Savas, T.L.; Miikkulainen, R.; Harper, C.B. Flavor-cyber-agriculture: Optimization of plant metabolites in an open-source control environment through surrogate modeling. PLoS ONE 2019, 14, e0213918. [Google Scholar] [CrossRef] [Green Version]
- Aldarkazali, M.; Rihan, H.Z.; Carne, D.; Fuller, M.P. The Growth and Development of Sweet Basil (Ocimum basilicum) and Bush Basil (Ocimum minimum) Grown under Three Light Regimes in a Controlled Environment. Agronomy 2019, 9, 743. [Google Scholar] [CrossRef] [Green Version]
- Amaki, W.; Yamazaki, N.; Ichimura, M.; Watanabe, H. Effects of Light Quality on the Growth and Essential Oil Content in Sweet Basil. Acta Hortic. 2011, 907, 91–94. [Google Scholar] [CrossRef]
- Avgoustaki, D.D. Optimization of Photoperiod and Quality Assessment of Basil Plants Grown in a Small-Scale Indoor Cultivation System for Reduction of Energy Demand. Energies 2019, 12, 3980. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hortic. 2016, 198, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Barbi, S.; Barbieri, F.; Bertacchini, A.; Barbieri, L.; Montorsi, M. Effects of Different LED Light Recipes and NPK Fertilizers on Basil Cultivation for Automated and Integrated Horticulture Methods. Appl. Sci. 2021, 11, 2497. [Google Scholar] [CrossRef]
- Carvalho, S.D.; Schwieterman, M.L.; Abrahan, C.E.; Colquhoun, T.A.; Folta, K.M. Light Quality Dependent Changes in Morphology, Antioxidant Capacity, and Volatile Production in Sweet Basil (Ocimum basilicum). Front. Plant Sci. 2016, 7, 1328. [Google Scholar] [CrossRef] [Green Version]
- Corrado, G.; Chiaiese, P.; Lucini, L.; Miras-Moreno, B.; Colla, G.; Rouphael, Y. Successive Harvests Affect Yield, Quality and Metabolic Profile of Sweet Basil (Ocimum basilicum L.). Agronomy 2020, 10, 830. [Google Scholar] [CrossRef]
- d’Aquino, L.; Lanza, B.; Gambale, E.; Sighicelli, M.; Menegoni, P.; Modarelli, G.C.; Rimauro, J.; Chianese, E.; Nenna, G.; Fasolino, T.; et al. Growth and metabolism of basil grown in a new-concept microcosm under different lighting conditions. Sci. Hortic. 2022, 299, 111035. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M. Photosynthesis, Morphology, Yield, and Phytochemical Accumulation in Basil Plants Influenced by Substituting Green Light for Partial Red and/or Blue Light. HortScience 2019, 54, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Responses of Sweet Basil to Different Daily Light Integrals in Photosynthesis, Morphology, Yield, and Nutritional Quality. HortScience 2018, 53, 496–503. [Google Scholar] [CrossRef]
- Frąszczak, B.; Golcz, A.; Zawirska-Wojtasiak, R.; Janowska, B. Growth rate of sweet basil and lemon balm plants grown under fluorescent lamps and led modules. Acta Sci. Pol. Hortorum Cultus 2014, 13, 3–13. [Google Scholar]
- Hosseini, A.; Mehrjerdi, M.Z.; Aliniaeifard, S.; Seif, M. Photosynthetic and growth responses of green and purple basil plants under different spectral compositions. Physiol. Mol. Biol. Plants 2019, 25, 741–752. [Google Scholar] [CrossRef]
- Ioannidis, D.; Bonner, L.; Johnson, C.B. UV-B is Required for Normal Development of Oil Glands in Ocimum basilicum L. (Sweet Basil). Ann. Bot. 2002, 90, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.B.; Kirby, J.; Naxakis, G.; Pearson, S. Substantial UV-B-mediated induction of essential oils in sweet basil (Ocimum basilicum L.). Phytochemistry 1999, 51, 507–510. [Google Scholar] [CrossRef]
- Kondrat’ev, V.M.; Osipova, G.S.; Kiselyov, M.V.; Gudiev, O.Y. Influence of LED lighting power on basil (Ocimum basilicum L.). IOP Conf. Ser. Earth Environ. Sci. 2021, 723, 032079. [Google Scholar] [CrossRef]
- Litvin, A.G.; Currey, C.J.; Wilson, L.A. Effects of Supplemental Light Source on Basil, Dill, and Parsley Growth, Morphology, Aroma, and Flavor. J. Amer. Soc. Hort. Sci. 2020, 145, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Lobiuc, A.; Vasilache, V.; Pintilie, O.; Stoleru, T.; Burducea, M.; Oroian, M.; Zamfirache, M. Blue and Red LED Illumination Improves Growth and Bioactive Compounds Contents in Acyanic and Cyanic Ocimum basilicum L. Microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef] [Green Version]
- Matysiak, B.; Kowalski, A. White, blue and red LED lighting on growth, morphology and accumulation of flavonoid compounds in leafy greens. Zemdirb. Agric. 2019, 106, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q.; Runkle, E.S. Far-red radiation interacts with relative and absolute blue and red photon flux densities to regulate growth, morphology, and pigmentation of lettuce and basil seedlings. Sci. Hortic. 2019, 255, 269–280. [Google Scholar] [CrossRef]
- Milenković, L.; Stanojević, J.; Cvetković, D.; Stanojević, L.; Lalević, D.; Šunić, L.; Fallik, E.; Ilić, Z.S. New technology in basil production with high essential oil yield and quality. Ind. Crops Prod. 2019, 140, 111718. [Google Scholar] [CrossRef]
- Mosadegh, H.; Trivellini, A.; Ferrante, A.; Lucchesini, M.; Vernieri, P.; Mensuali, A. Applications of UV-B lighting to enhance phenolic accumulation of sweet basil. Sci. Hortic. 2018, 229, 107–116. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Pennisi, G.; Pistillo, A.; Orsini, F.; Cellini, A.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Crepaldi, A.; Gianquinto, G.; Marcelis, L.F.M. Optimal light intensity for sustainable water and energy use in indoor cultivation of lettuce and basil under red and blue LEDs. Sci. Hortic. 2020, 272, 109508. [Google Scholar] [CrossRef]
- Piovene, C.; Orsini, F.; Bosi, S.; Sanoubar, R.; Bregola, V.; Dinelli, G.; Gianquinto, G. Optimal red:blue ratio in led lighting for nutraceutical indoor horticulture. Sci. Hortic. 2015, 193, 202–208. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Aljafer, N.; Jbara, M.; McCallum, L.; Lengger, S.; Fuller, M.P. The Impact of LED Lighting Spectra in a Plant Factory on the Growth, Physiological Traits and Essential Oil Content of Lemon Balm (Melissa officinalis). Plants 2022, 11, 342. [Google Scholar] [CrossRef]
- Samuoliene, G.; Brazaityte, A.; Sirtatutas, R.; Sakalauskiene, S.; Jankauskiene, J.; Duchovskis, P.; Novičkovas, A. The Impact of Supplementary Short-Term Red LED Lighting on the Antioxidant Properties of Microgreens. Acta Hortic. 2012, 956, 649–655. [Google Scholar] [CrossRef]
- Schenkels, L.; Saeys, W.; Lauwers, A.; De Proft, M.P. Green light induces shade avoidance to alter plant morphology and increases biomass production in Ocimum basilicum L. Sci. Hortic. 2020, 261, 109002. [Google Scholar] [CrossRef]
- Shiga, T.; Shoji, K.; Shimada, H.; Hashida, S.; Goto, F.; Yoshihara, T. Effect of light quality on rosmarinic acid content and antioxidant activity of sweet basil, Ocimum basilicum L. Plant Biotechnol 2009, 26, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Solbach, J.A.; Fricke, A.; Stützel, H. Seasonal Efficiency of Supplemental LED Lighting on Growth and Photomorphogenesis of Sweet Basil. Front. Plant Sci. 2021, 12, 609975. [Google Scholar] [CrossRef]
- Tarakanov, I.; Yakovleva, O.; Konovalova, I.; Paliutina, G.; Anisimov, A. Light-Emitting Diodes: On the Way to Combinatorial Lighting Technologies for Basic Research and Crop Production. Acta Hortic. 2012, 956, 171–178. [Google Scholar] [CrossRef]
- Taulavuori, K.; Hyöky, V.; Oksanen, J.; Taulavuori, E.; Julkunen-Tiitto, R. Species-specific differences in synthesis of flavonoids and phenolic acids under increasing periods of enhanced blue light. Environ. Exp. Bot. 2016, 121, 145–150. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Beynon-Davies, R.; Carslaw, N.; Dodd, I.C.; Ashworth, K. Yield, resource use efficiency or flavour: Trade-offs of varying blue-to-red lighting ratio in urban plant factories. Sci. Hortic. 2022, 295, 110802. [Google Scholar] [CrossRef]
- Paradiso, R.; Arena, C.; Rouphael, Y.; d’Aquino, L.; Makris, K.; Vitaglione, P.; De Pascale, S. Growth, photosynthetic activity and tuber quality of two potato cultivars in controlled environment as affected by light source. Plant Biosyst. 2019, 153, 725–735. [Google Scholar] [CrossRef]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef] [Green Version]
- Bolhàr-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Öquist, G.; Schreider, U.; Lechner, E.G. Chlorophyll fluorescence as probe of the photosynthetic competence of leaves in the field: A review of current instrument. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Hamadani, S.; Khan, N.; Perveen, S.; Qu, M.; Jiang, J.; Govindjee; Zhu, X. Changes in the photosynthesis properties and photoprotection capacity in rice (Oryza sativa) grown under red, blue, or white light. Photosynth. Res. 2019, 139, 107–121. [Google Scholar] [CrossRef]
- Janeeshma, E.; Johnson, R.; Amritha, M.S.; Noble, L.; Aswathi, K.P.R.; Telesiński, A.; Kalaji, H.M.; Auriga, A.; Puthur, J.T. Modulations in Chlorophyll a Fluorescence Based on Intensity and Spectral Variations of Light. Int. J. Mol. Sci. 2022, 23, 5599. [Google Scholar] [CrossRef] [PubMed]
- Yousef, A.F.; Ali, M.M.; Rizwan, H.M.; Tadda, S.A.; Kalaji, H.M.; Yang, H.; Ahmed, M.A.A.; Wróbel, J.; Xu, Y.; Chen, F. Photosynthetic apparatus performance of tomato seedlings grown under various combinations of LED illumination. PLoS ONE 2021, 16, e0249373. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Kim, H.-J. Characterizing Nutrient Composition and Concentration in Tomato-, Basil-, and Lettuce-Based Aquaponic and Hydroponic Systems. Water 2020, 12, 1259. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Suh, J.H. Multi-omics approach in tea polyphenol research regarding tea plant growth, development and tea processing: Current technologies and perspectives. Food Sci. Hum. Wellness 2022, 11, 524–536. [Google Scholar] [CrossRef]
- Theodoridis, G.; Pechlivanis, A.; Thomaidis, N.S.; Spyros, A.; Georgiou, C.A.; Albanis, T.; Skoufos, I.; Kalogiannis, S.; Tsangaris, G.T.; Stasinakis, A.S.; et al. FoodOmicsGR_RI: A Consortium for Comprehensive Molecular Characterisation of Food Products. Metabolites 2021, 11, 74. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Devi, P.; Chaudhary, S.; Rani, A.; Jha, U.C.; Kumar, S.; Bindumadhava, H.; Vara Prasad, P.V.; Sharma, K.D.; Siddique, K.H.M.; et al. ‘Omics’ approaches in developing combined drought and heat tolerance in food crops. Plant Cell Rep. 2022, 41, 699–739. [Google Scholar] [CrossRef]
- Nataraja, K.N.; Madhura, B.G.; Parvathi, M.S. OMICS: Modern tools for precise understanding of drought adaptation in plants. In Plant OMICS and Crop Breeding, 1st ed.; Zargar, S.M., Rai, V., Eds.; Apple Academic Press Inc.: New York, NY, USA, 2017; pp. 263–294. [Google Scholar] [CrossRef]
- Khaki, A.; Fathiazad, F.; Nouri, M.; Khaki, A.A. Effect of Ocimum basilicum on apoptosis in testis of rats after exposure to electromagnetic field. Afr. J. Pharm. Pharmacol. 2011, 5, 1534–1537. [Google Scholar] [CrossRef]
- Tsai, Y.L.; Chiou, S.Y.; Chan, K.C.; Sung, J.M.; Lin, S.D. Caffeic acid derivatives, total phenols, antioxidant and antimutagenic activities of Echinacea purpurea flower extracts. LWT-Food Sci. Technol. 2012, 46, 169–176. [Google Scholar] [CrossRef]
- Lee, J.; Scagel, C.F. Chicoric acid found in basil (Ocimum basilicum L.) leaves. Food Chem. 2009, 115, 650–656. [Google Scholar] [CrossRef]
- Sarais, G.; D’Urso, G.; Lai, C.; Pirisi, F.M.; Pizza, C.; Montoro, P. Targeted and untargeted mass spectrometric approaches in discrimination between Myrtus communis cultivars from Sardinia region. J. Mass Spectrom. 2016, 51, 704–715. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
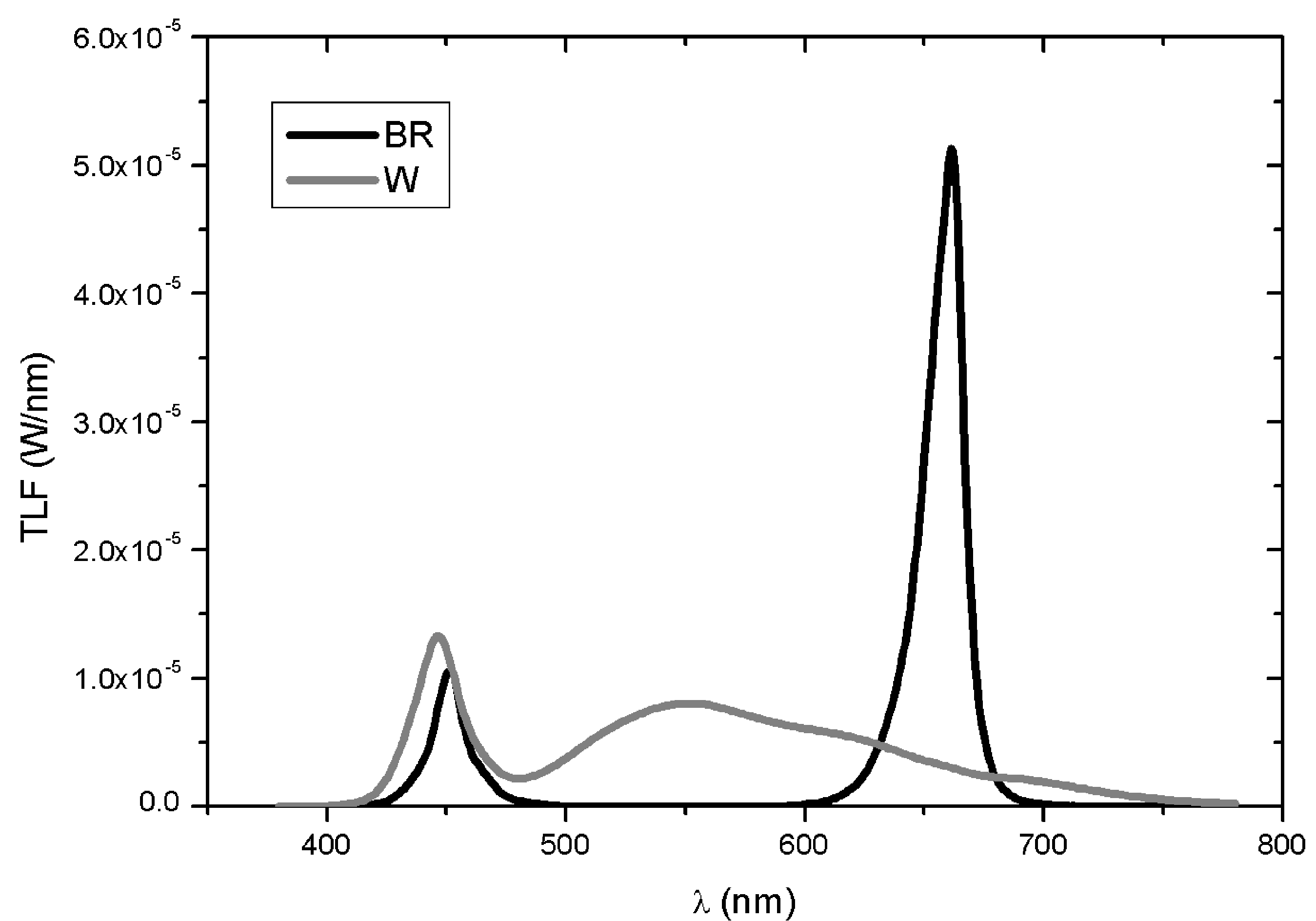{kind=link}
| Ion Concentrations (mg g−1) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | NH4+ | NO3− | P | PO43− | K+ | Mg++ | Ca++ | S | SO42− | Na+ | Cl− | ∑ions | ||
| Leaves | W | 6.21 ± 0.04 | 1.17 ± 0.04 | 23.47 ± 0.04 | 2.071 ± 0.009 | 6.35 ± 0.03 | 49.58 ± 0.03 | 3.72 ± 0.03 | 3.49 ± 0.02 | 1.33 ± 0.00 | 4.00 ± 0.01 | 0.01 ± 0.01 | 12.01 ± 0.01 | 113.37 ± 0.25 |
| BR | 5.95 ± 0.02 | 1.57 ± 0.02 | 20.96 ± 0.02 | 2.619 ± 0.004 | 8.03 ± 0.01 | 46.54 ± 0.02 | 4.34 ± 0.02 | 4.93 ± 0.03 | 1.49 ± 0.01 | 4.48 ± 0.02 | 0.01 ± 0.01 | 11.37 ± 0.02 | 112.30 ± 0.20 | |
| Stems | W | 4.12 ± 0.05 | 0.87 ± 0.05 | 15.25 ± 0.02 | 1.451 ± 0.006 | 4.45 ± 0.02 | 39.28 ± 0.01 | 1.94 ± 0.02 | 2.17 ± 0.02 | 0.76 ± 0.00 | 2.29 ± 0.01 | 1.72 ± 0.02 | 6.94 ± 0.01 | 81.25 ± 0.24 |
| BR | 3.01 ± 0.00 | 0.80 ± 0.00 | 10.58 ± 0.01 | 1.414 ± 0.007 | 4.33 ± 0.02 | 28.68 ± 0.01 | 1.45 ± 0.01 | 1.76 ± 0.01 | 0.88 ± 0.00 | 2.64 ± 0.01 | 0.92 ± 0.02 | 4.61 ± 0.02 | 61.09 ± 0.12 | |
| Leaves/Stems ratio | W | 1.51 | 1.34 | 1.54 | 1.428 | 1.43 | 1.26 | 1.92 | 1.59 | 1.74 | 1.74 | 0.01 | 1.73 | 1.39 |
| BR | 1.98 | 1.95 | 1.98 | 1.853 | 1.85 | 1.62 | 3 | 2.8 | 1.7 | 1.7 | 0.01 | 2.46 | 1.84 | |
| Significance | N | NH4+ | NO3− | P | PO43− | K+ | Mg++ | Ca++ | S | SO42− | Na+ | Cl− | ∑ions | |
| Leaves | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.895 ns | 0.00 ** | 0.006 ** | |
| Stems | 0.00 ** | 0.107 ** | 0.00 ** | 0.002 ** | 0.002 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.002 ** | |
| Leaves/Stems ratio | 0.00 ** | 0.001 *** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** | 0.003 ** | 0.527 ns | 0.00 ** | 0.00 ** | |
| N. | RT | [M-H]− | Molecular Formula | Δ ppm | MS/MS | Identity |
|---|---|---|---|---|---|---|
| 1 | 2.37 | 227.0611 | C7H13O8 | 2.6 | 179 | caffeic acid + formiate adduct |
| 2 | 3.86 | 263.0043 | C8H7O10 | 4.7 | 244 | sugar |
| 3 | 3.86 | 355.1237 | C13H23O11 | 1.9 | 193 | sugar |
| 4 | 10.20 | 593.1497 | C27H29O15 | 0.5 | 353/473 | vicenin 2 |
| 5 | 12.47 | 197.0454 | C9H9O5 | 3.09 | 153/129 | syringic acid |
| 6 | 12.56 | 463.0868 | C21H19O12 | 0.5 | 301 | quercetin 3-O-glucoside |
| 7 | 12.81 | 300.9982 | C14H5O8 | 1.01 | 283/257/233/227 | ellagic acid |
| 8 | 13.22 | 473.0718 | C22H17O12 | chicoric acid | ||
| 9 | 13.80 | 353.0872 | C16H17O9 | 1.4 | - | cholorogenic acid |
| 10 | 13.85 | 665.3887 | C36H57O11 | 621/431 | triterpene | |
| 11 | 13.96 | 311.1130 | C15H19O7 | 183/267/293 | 1-O-β-d-glucopyranosyloxy-2-hydroxy-4-allylbenzene | |
| 12 | 14.39 | 475.0869 | C22H19O12 | −0.3 | 285 | luteolin 7-O-glucuronide-6′-methyl ester |
| 13 | 14.61 | 359.0760 | C18H15O8 | 1.02 | 161 | rosmarinic acid |
| 14 | 15.09 | 738.3691 | C37H56O14N | 0.7 | 648/283 | unknown |
| 15 16 | 15.64 15.80 | 459.0923 327.0868 | C22H19O11 C18H15O6 | 0.35 1.5 | 269 185/199/283/309 | apigenin 7-O-glucuronide methyl ester |
| 17 | 15.93 | 193.0505 | C10H9O4 | 5.04 | 161/134/178 | ferulic acid |
| 18 | 16.00 | 565.1550 | C26H29O14 | 0.1 | 367/197 | unknown |
| 19 | 16.40 | 227.0708 | C14H11O3 | 17 | resveratrol | |
| 20 | 17.48 | 373.0922 | C19H17O8 | 1.4 | 179/135 | rosmarinic acid methyl ester |
| 21 | 18.74 | 327.0868 | C18H15O6 | 1.5 | 185/199/283/309 | salvigenin |
| 22 | 18.74 | 565.1343 | C29H25O12 | 0.72 | 533/353 | unknown |
| 23 | 19.14 | 311.1680 | C13 H27 O8 | 0.82 | 183 | unknown |
| 24 | 19.28 | 357.0971 | C19H17O7 | 0.6 | 339/313/289 | gardenin b |
| 25 | 20.00 | 501.1026 | C24H21O12 | 339/161 | di-feruloyl-tartaric acid | |
| 26 | 20.88 | 289.0687 | C15H13O6 | −6.9 | catechin | |
| 27 | 28.30 | 291.1989 | C15H31O3S | 1.6 | 198 | sulfurous acid |
| 28 | 29.20 | 309.1734 | C14H29O5S | 1.7 | 96 | ethanol, 2-(dodecyloxy), 1-(hydrogen sulfate) |
| 29 | 30.91 | 455.3515 | C30H47O3 | −0.15 | 283/193 | oleanolic acid |
| 30 | 37.60 | 455.3517 | C30H47O3 | −0.15 | 283/199 | ursolic acid |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
d’Aquino, L.; Cozzolino, R.; Nardone, G.; Borelli, G.; Gambale, E.; Sighicelli, M.; Menegoni, P.; Modarelli, G.C.; Rimauro, J.; Chianese, E.; et al. Effects of White and Blue-Red Light on Growth and Metabolism of Basil Grown under Microcosm Conditions. Plants 2023, 12, 1450. https://doi.org/10.3390/plants12071450
d’Aquino L, Cozzolino R, Nardone G, Borelli G, Gambale E, Sighicelli M, Menegoni P, Modarelli GC, Rimauro J, Chianese E, et al. Effects of White and Blue-Red Light on Growth and Metabolism of Basil Grown under Microcosm Conditions. Plants. 2023; 12(7):1450. https://doi.org/10.3390/plants12071450
Chicago/Turabian Styled’Aquino, Luigi, Rosaria Cozzolino, Giovanni Nardone, Gianni Borelli, Emilia Gambale, Maria Sighicelli, Patrizia Menegoni, Giuseppe Carlo Modarelli, Juri Rimauro, Elena Chianese, and et al. 2023. "Effects of White and Blue-Red Light on Growth and Metabolism of Basil Grown under Microcosm Conditions" Plants 12, no. 7: 1450. https://doi.org/10.3390/plants12071450