
Comprehensive Identification and Expression Analysis of the YTH Family of RNA-Binding Proteins in Strawberry

Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Characterization of FvYTHs in Strawberry
2.2. Chromosome Localization and Synteny Relationships of FvYTH Genes
2.3. Gene Structure, Conserved Domain, and Motif Analysis of FvYTHs
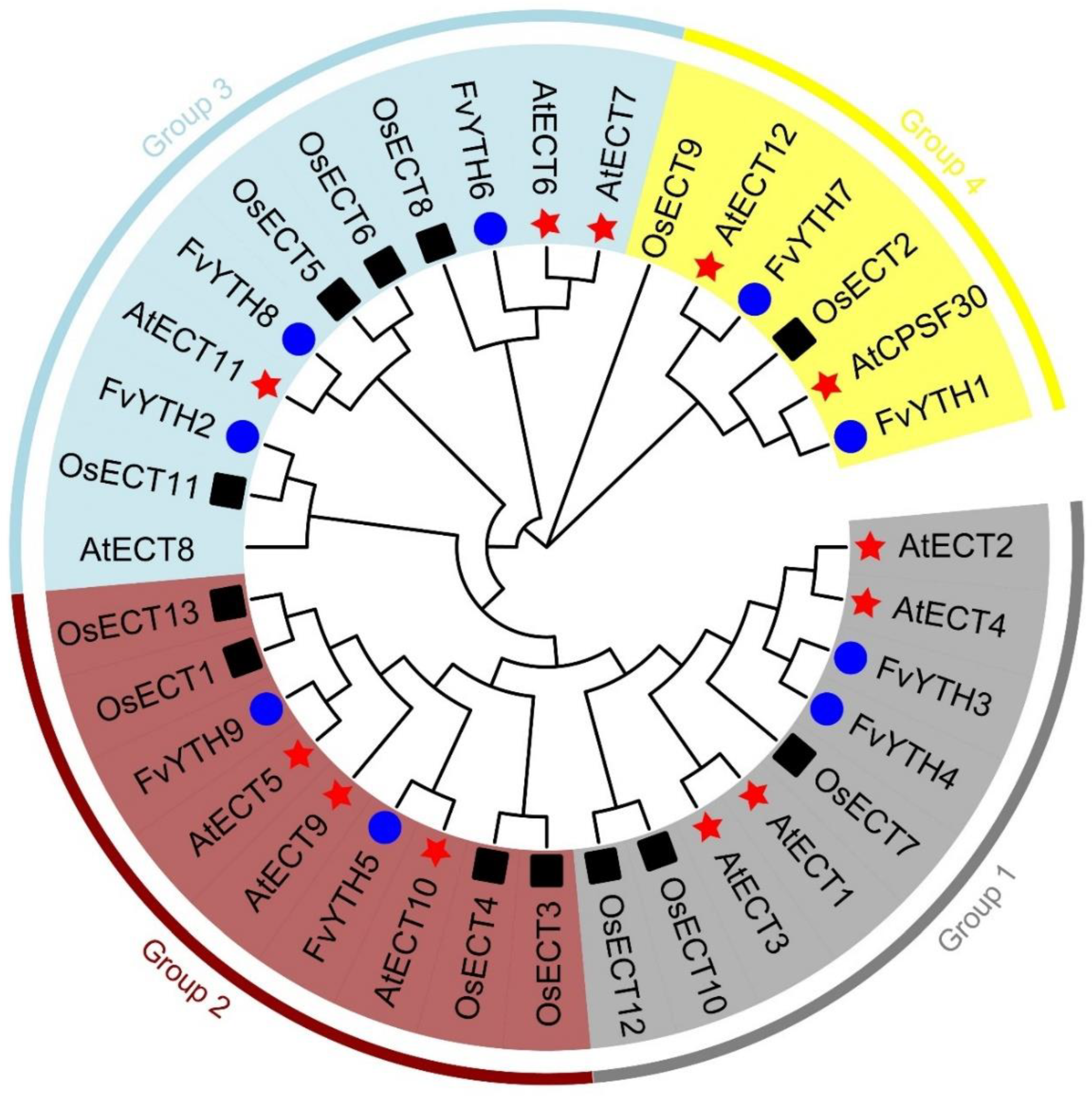
2.4. Phylogenetic Analysis of YTH Proteins in Strawberry
2.5. Cis-Acting Regulatory Elements in Promoter Regions of FvYTH Genes
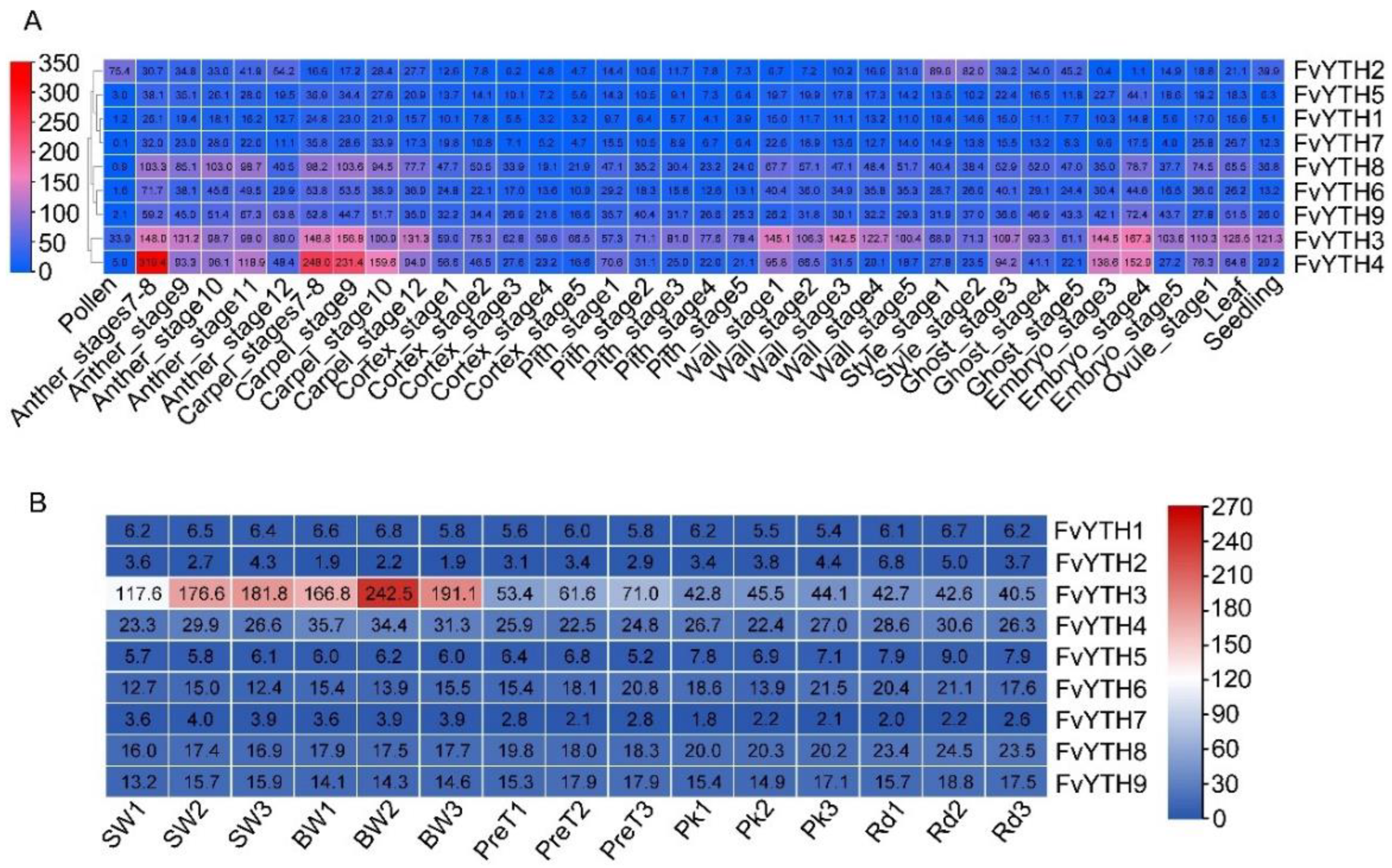
2.6. Analysis of FvYTH Gene Expression in Different Tissues and Fruit Development Stages
3. Discussion
4. Materials and Methods
4.1. Identification and Protein Properties of YTH Proteins in Strawberry
4.2. Chromosome Localization, Synteny Relationship, Gene Structure, Conserved Domain, and Motif Analysis
4.3. Phylogenetic Analysis and Cis-Acting Element Prediction in Promoters
4.4. Expression Analysis of FvYTH Gene
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A Landscape Takes Shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piątkowski, P.; Bagiński, B.; Wirecki, T.K.; de Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A Database of RNA Modification Pathways. 2017 Update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene Expression Regulation Mediated through Reversible M6A RNA Methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Bokar, J.A.; Shambaugh, M.E.; Polayes, D.; Matera, A.G.; Rottman, F.M. Purification and CDNA Cloning of the AdoMet-Binding Subunit of the Human MRNA (N6-Adenosine)-Methyltransferase. RNA 1997, 3, 1233–1247. [Google Scholar]
- Bujnicki, J.M.; Feder, M.; Radlinska, M.; Blumenthal, R.M. Structure Prediction and Phylogenetic Analysis of a Functionally Diverse Family of Proteins Homologous to the MT-A70 Subunit of the Human MRNA:M(6)A Methyltransferase. J. Mol. Evol. 2002, 55, 431–444. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 Complex Mediates Mammalian Nuclear RNA N6-Adenosine Methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Ping, X.-L.; Sun, B.-F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.-J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.-S.; et al. Mammalian WTAP Is a Regulatory Subunit of the RNA N6-Methyladenosine Methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Mei, Y.; Hou, M. Knockdown RBM15 Inhibits Colorectal Cancer Cell Proliferation and Metastasis Via N6-Methyladenosine (M6A) Modification of MyD88 MRNA. Cancer Biother. Radio. 2022, 37, 976–986. [Google Scholar] [CrossRef]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.-J.; Chen, Q.; et al. Reversible Methylation of M6Am in the 5′ Cap Controls MRNA Stability. Nature 2017, 541, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.-G.; et al. N6-Methyladenosine in Nuclear RNA Is a Major Substrate of the Obesity-Associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Jia, G.; Fu, Y.; He, C. Reversible RNA Adenosine Methylation in Biological Regulation. Trends Genet. 2013, 29, 108–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, D.P.; Pickering, B.F.; Jaffrey, S.R. Reading M6A in the Transcriptome: M6A-Binding Proteins. Trends Cell Biol. 2018, 28, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Hsu, P.J.; Xing, X.; Fang, J.; Lu, Z.; Zou, Q.; Zhang, K.-J.; Zhang, X.; Zhou, Y.; Zhang, T.; et al. Mettl3-/Mettl14-Mediated MRNA N6-Methyladenosine Modulates Murine Spermatogenesis. Cell Res. 2017, 27, 1216–1230. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Shi, H.; Wang, F.; Wang, Y. M 6 A MRNA Methylation Controls Autophagy and Adipogenesis by Targeting Atg5 and Atg7. Autophagy 2020, 16, 1221–1235. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Zhang, T.; Yang, Y.; Zhang, D.; Gao, J.; Li, J.; Yue, L.; Gao, D.; Shi, B.; Han, Y.; et al. METTL3-Mediated MRNA N6-Methyladenosine Is Required for Oocyte and Follicle Development in Mice. Cell Death Dis. 2021, 12, 989. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Wang, X.; Xu, Z.; Cao, Y.; Gong, R.; Yu, Y.; Yu, Y.; Guo, X.; Liu, S.; Yu, M.; et al. ALKBH5 Regulates Cardiomyocyte Proliferation and Heart Regeneration by Demethylating the MRNA of YTHDF1. Theranostics 2021, 11, 3000–3016. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liang, Z.; Gu, X.; Chen, Y.; Teo, Z.W.N.; Hou, X.; Cai, W.M.; Dedon, P.C.; Liu, L.; Yu, H. N6-Methyladenosine RNA Modification Regulates Shoot Stem Cell Fate in Arabidopsis. Dev. Cell 2016, 38, 186–200. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.-C.; Wei, L.-H.; Zhang, C.; Wang, Y.; Chen, L.; Lu, Z.; Chen, P.R.; He, C.; Jia, G. ALKBH10B Is an RNA N 6 -Methyladenosine Demethylase Affecting Arabidopsis Floral Transition. Plant Cell 2017, 29, 2995–3011. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Tang, R.; Li, X.; Tian, S.; Li, B.; Qin, G. N6-Methyladenosine RNA Modification Regulates Strawberry Fruit Ripening in an ABA-Dependent Manner. Genome Biol. 2021, 22, 168. [Google Scholar] [CrossRef]
- Hou, N.; Li, C.; He, J.; Liu, Y.; Yu, S.; Malnoy, M.; Mobeen Tahir, M.; Xu, L.; Ma, F.; Guan, Q. MdMTA-mediated m 6 A Modification Enhances Drought Tolerance by Promoting MRNA Stability and Translation Efficiency of Genes Involved in Lignin Deposition and Oxidative Stress. New Phytol. 2022, 234, 1294–1314. [Google Scholar] [CrossRef]
- Zhou, L.; Tian, S.; Qin, G. RNA Methylomes Reveal the M6A-Mediated Regulation of DNA Demethylase Gene SlDML2 in Tomato Fruit Ripening. Genome Biol. 2019, 20, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, S.; Sun, H.; Xu, C. YTH Domain: A Family of N 6 -Methyladenosine (m 6 A) Readers. Genom. Proteom. Bioinf. 2018, 16, 99–107. [Google Scholar] [CrossRef]
- Meyer, K.D.; Jaffrey, S.R. Rethinking m 6 A Readers, Writers, and Erasers. Annu. Rev. Cell Dev. Biol. 2017, 33, 319–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhang, H.; Hong, Y.; Huang, L.; Li, X.; Zhang, Y.; Ouyang, Z.; Song, F. Genome-Wide Identification, Biochemical Characterization, and Expression Analyses of the YTH Domain-Containing RNA-Binding Protein Family in Arabidopsis and Rice. Plant Mol. Biol. Rep. 2014, 32, 1169–1186. [Google Scholar] [CrossRef]
- Arribas-Hernández, L.; Bressendorff, S.; Hansen, M.H.; Poulsen, C.; Erdmann, S.; Brodersen, P. An m6A-YTH Module Controls Developmental Timing and Morphogenesis in Arabidopsis. Plant Cell 2018, 30, 952–967. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Sun, J.; Wu, B.; Gao, Y.; Nie, H.; Nie, Z.; Quan, S.; Wang, Y.; Cao, X.; Li, S. CPSF30-L-Mediated Recognition of MRNA M6A Modification Controls Alternative Polyadenylation of Nitrate Signaling-Related Gene Transcripts in Arabidopsis. Mol. Plant 2021, 14, 688–699. [Google Scholar] [CrossRef]
- Song, P.; Yang, J.; Wang, C.; Lu, Q.; Shi, L.; Tayier, S.; Jia, G. Arabidopsis N6-Methyladenosine Reader CPSF30-L Recognizes FUE Signals to Control Polyadenylation Site Choice in Liquid-like Nuclear Bodies. Mol. Plant 2021, 14, 571–587. [Google Scholar] [CrossRef]
- Guo, T.; Liu, C.; Meng, F.; Hu, L.; Fu, X.; Yang, Z.; Wang, N.; Jiang, Q.; Zhang, X.; Ma, F. The m 6 A Reader MhYTP2 Regulates MdMLO19 MRNA Stability and Antioxidant Genes Translation Efficiency Conferring Powdery Mildew Resistance in Apple. Plant Biotechnol. J. 2022, 20, 511–525. [Google Scholar] [CrossRef]
- Muthusamy, M.; Kim, J.-H.; Kim, J.A.; Lee, S.-I. Plant RNA Binding Proteins as Critical Modulators in Drought, High Salinity, Heat, and Cold Stress Responses: An Updated Overview. Int. J. Mol. Sci. 2021, 22, 6731. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Ao, Q.; Tan, C.; Yang, Y. Genome-Wide Identification and Characterization of YTH Domain-Containing Genes, Encoding the M6A Readers, and Their Expression in Tomato. Plant Cell Rep. 2021, 40, 1229–1245. [Google Scholar] [CrossRef]
- Wang, N.; Yue, Z.; Liang, D.; Ma, F. Genome-Wide Identification of Members in the YTH Domain-Containing RNA-Binding Protein Family in Apple and Expression Analysis of Their Responsiveness to Senescence and Abiotic Stresses. Gene 2014, 538, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Bie, X.M.; Wang, N.; Zhang, X.S.; Gao, X.-Q. Genome-Wide Identification and Expression Analysis of YTH Domain-Containing RNA-Binding Protein Family in Common Wheat. BMC Plant Biol. 2020, 20, 351. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhuang, X. M6A-Binding YTHDF Proteins Promote Stress Granule Formation. Nat. Chem. Biol. 2020, 16, 955–963. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Diao, J.; Shi, Y.G.; Shi, Y.; Ma, H.; Shen, H. Binding to M6A RNA Promotes YTHDF2-Mediated Phase Separation. Protein Cell 2020, 11, 304–307. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Pei, G.; Li, D.; Li, R.; Shao, Y.; Zhang, Q.C.; Li, P. Multivalent M6A Motifs Promote Phase Separation of YTHDF Proteins. Cell Res. 2019, 29, 767–769. [Google Scholar] [CrossRef]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, Writing and Erasing MRNA Methylation. Nat. Rev. Mol. Cell Bio. 2019, 20, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Boeynaems, S.; Alberti, S.; Fawzi, N.L.; Mittag, T.; Polymenidou, M.; Rousseau, F.; Schymkowitz, J.; Shorter, J.; Wolozin, B.; Van Den Bosch, L.; et al. Protein Phase Separation: A New Phase in Cell Biology. Trends Cell Biol. 2018, 28, 420–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Yue, H.; Nie, X.; Yan, Z.; Weining, S. N6-Methyladenosine Regulatory Machinery in Plants: Composition, Function and Evolution. Plant Biotechnol. J. 2019, 17, 1194–1208. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A Webserver for Visualization, Annotation, and Management of Phylogenetic Trees. Nucleic. Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic. Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.; Jia, S.; Huang, X.; Wang, L.; Fu, W.; Huo, G.; Gan, L.; Ding, J.; Li, Y. Transcriptome and Hormone Analyses Provide Insights into Hormonal Regulation in Strawberry Ripening. Planta 2019, 250, 145–162. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Molecular Weigh (Da) | Isoelectric Point | Length (aa) |
|---|---|---|---|---|
| FvYTH1 | FvH4_3g09980 | 75,927.17 | 6.37 | 689 |
| FvYTH2 | FvH4_3g13840 | 65,995.31 | 6.37 | 592 |
| FvYTH3 | FvH4_3g45840 | 84,983.28 | 8.53 | 773 |
| FvYTH4 | FvH4_3g45841 | 69,024.37 | 5.67 | 631 |
| FvYTH5 | FvH4_4g00190 | 75,181.65 | 5.12 | 682 |
| FvYTH6 | FvH4_4g21030 | 75,433.06 | 6.1 | 686 |
| FvYTH7 | FvH4_5g01140 | 40,460.09 | 6.24 | 355 |
| FvYTH8 | FvH4_5g30420 | 58,841.15 | 6.44 | 534 |
| FvYTH9 | FvH4_6g36470 | 72,549.24 | 5.51 | 659 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, P.; Li, X.; Fan, J.; Wang, C.; Lin, A.; Lian, H. Comprehensive Identification and Expression Analysis of the YTH Family of RNA-Binding Proteins in Strawberry. Plants 2023, 12, 1449. https://doi.org/10.3390/plants12071449
Xu P, Li X, Fan J, Wang C, Lin A, Lian H. Comprehensive Identification and Expression Analysis of the YTH Family of RNA-Binding Proteins in Strawberry. Plants. 2023; 12(7):1449. https://doi.org/10.3390/plants12071449
Chicago/Turabian StyleXu, Pengbo, Xinyu Li, Junmiao Fan, Chong Wang, Anqi Lin, and Hongli Lian. 2023. "Comprehensive Identification and Expression Analysis of the YTH Family of RNA-Binding Proteins in Strawberry" Plants 12, no. 7: 1449. https://doi.org/10.3390/plants12071449