

Interactive Temperature and CO2 Rise, Salinity, Drought, and Bacterial Inoculation Alter the Content of Fatty Acids, Total Phenols, and Oxalates in the Edible Halophyte Salicornia ramosissima
 , , , ,
, , , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
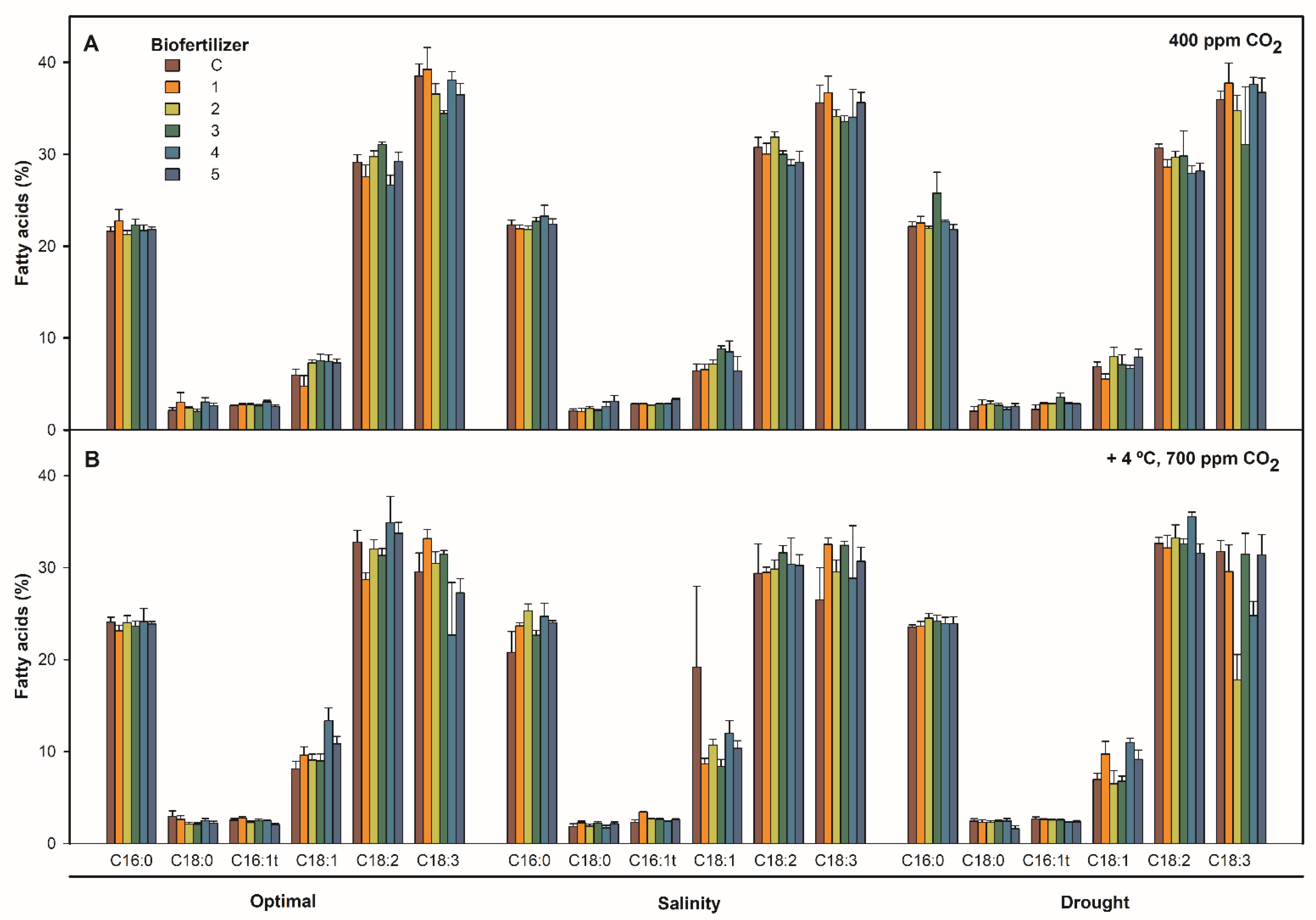
2.1. Effect of Combined Temperature, CO2, Irrigation, and Biofertilization on S. ramosissima Shoots Fatty Acid Content
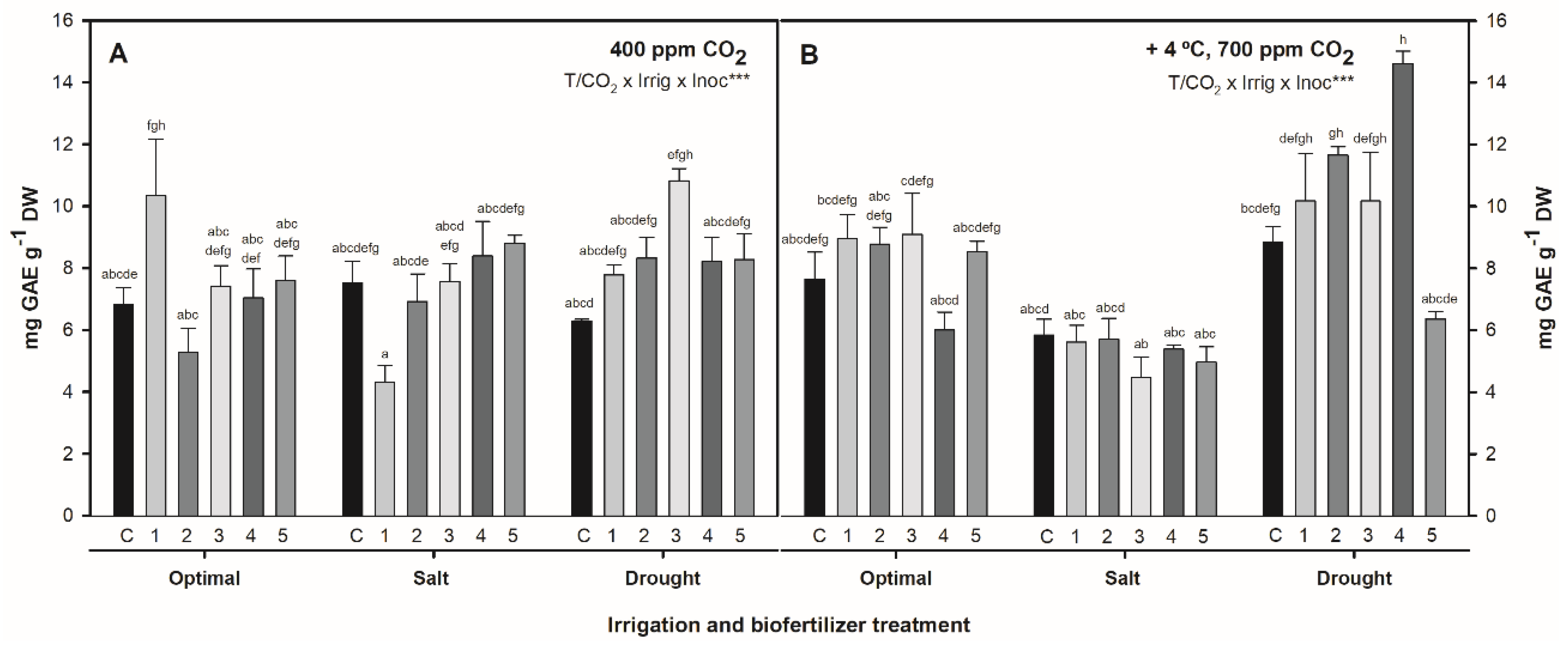
2.2. Effect of Combined Temperature, CO2, Irrigation, and Biofertilization on Total Phenol Content in S. ramosissima Shoots
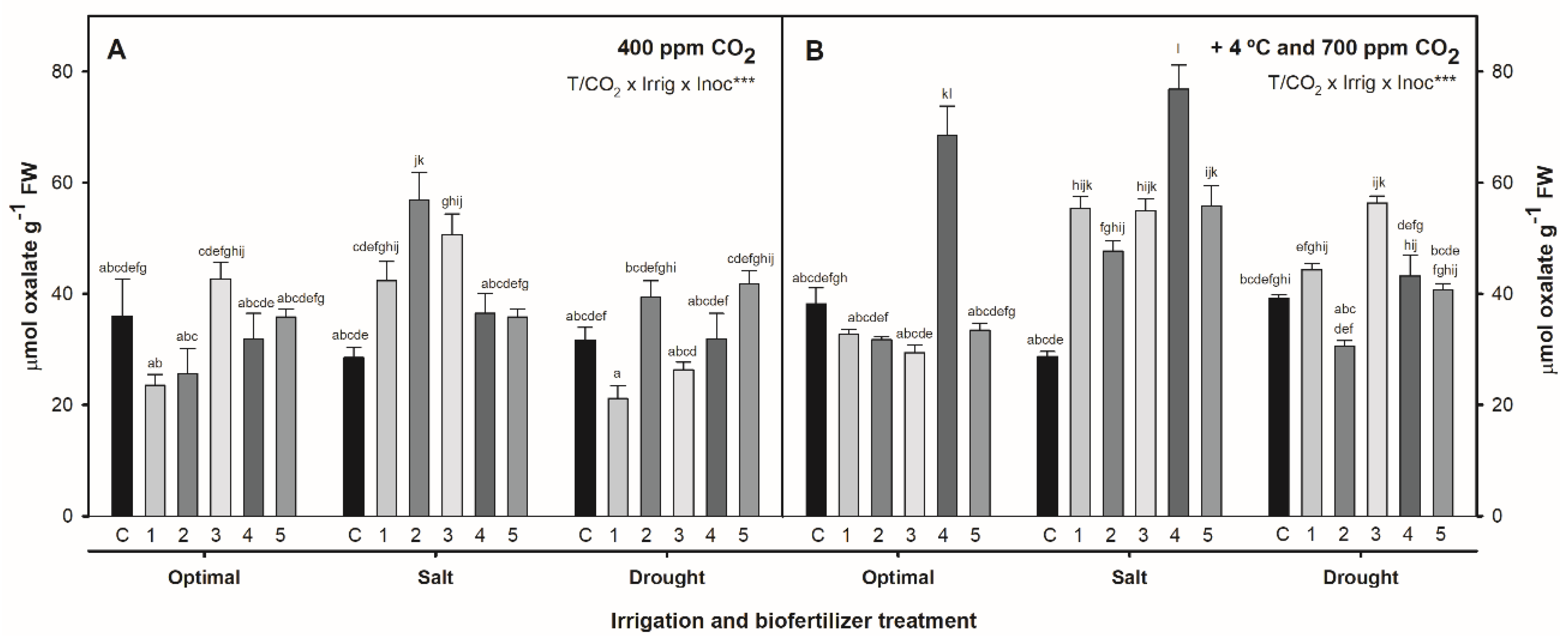
2.3. Effect of Combined Temperature, CO2, Irrigation, and Biofertilization on Oxalate Concentration in S. ramosissima Shoots
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design and Treatments
4.3. Rhizobacteria Selection and Inoculation
4.4. Fatty Acid Profiling of S. ramosissima Shoots
4.5. Determination of Total Phenolic Content in S. ramosissima Shoots
4.6. Determination of Oxalate Content in S. ramosissima Shoots
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiong, W.; Reynolds, M.; Xu, Y.; Patron, N.J.; Burgess, S.J. Climate change challenges plant breeding. Curr. Opin. Plant Biol. 2022, 70, 102308. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [PubMed]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt Tolerance and Crop Potential of Halophytes. CRC Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar]
- Loconsole, D.; Cristiano, G.; De Lucia, B. Glassworts: From wild salt marsh species to sustainable edible crops. Agriculture 2019, 9, 14. [Google Scholar]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar]
- Yensen, N.P. Halophyte uses for the twenty-first century. In Ecophysiology of High Salinity Tolerant Plants; Khan, M.A., Weber, D.J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 367–396. [Google Scholar]
- Patel, S. Salicornia: Evaluating the halophytic extremophile as a food and a pharmaceutical candidate. 3 Biotech 2016, 6, 104. [Google Scholar] [PubMed] [Green Version]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2015, 115, 529–540. [Google Scholar] [PubMed] [Green Version]
- Barreira, L.; Resek, E.; Rodrigues, M.J.; Rocha, M.I.; Pereira, H.; Bandarra, N.; Moreira Da Silva, M.; Varela, J.; Custódio, L. Halophytes: Gourmet food with nutritional health benefits? J. Food Compos. Anal. 2017, 59, 35–42. [Google Scholar]
- Ventura, Y.; Sagi, M. Halophyte crop cultivation: The case for Salicornia and Sarcocornia. Environ. Exp. Bot. 2013, 92, 144–153. [Google Scholar]
- Centofanti, T.; Bañuelos, G. Practical Uses of Halophytic Plants as Sources of Food and Fodder. In Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; Hasanuzzaman, M., Shabala, S., Fujita, M., Eds.; CABI: Boston, MA, USA, 2019; pp. 324–342. ISBN 9781786394330. [Google Scholar]
- Maciel, E.; Lillebø, A.; Domingues, P.; da Costa, E.; Calado, R.; Domingues, M.R.M. Polar lipidome profiling of Salicornia ramosissima and Halimione portulacoides and the relevance of lipidomics for the valorization of halophytes. Phytochemistry 2018, 153, 94–101. [Google Scholar]
- Limongelli, F.; Crupi, P.; Clodoveo, M.L.; Corbo, F.; Muraglia, M. Overview of the Polyphenols in Salicornia: From Recovery to Health-Promoting Effect. Molecules 2022, 27, 7954. [Google Scholar] [PubMed]
- Grigore, M.; Oprica, L. Halophytes as Possible Source of Antioxidant Compounds, in a Scenario Based on Threatened Agriculture and Food Crisis. Iran J. Public Health 2015, 44, 1153–1155. [Google Scholar] [PubMed]
- Ventura, Y.; Wuddineh, W.A.; Myrzabayeva, M.; Alikulov, Z.; Khozin-Goldberg, I.; Shpigel, M.; Samocha, T.M.; Sagi, M. Effect of seawater concentration on the productivity and nutritional value of annual Salicornia and perennial Sarcocornia halophytes as leafy vegetable crops. Sci. Hortic. (Amst.) 2011, 128, 189–196. [Google Scholar] [CrossRef]
- Guil, J.L.; Rodríguez-Garcí, I.; Torija, E. Nutritional and toxic factors in selected wild edible plants. Plant Foods Hum. Nutr. 1997, 51, 99–107. [Google Scholar] [CrossRef]
- Austenfeld, F.-A.; Leder, U. The Influence of Alkaline Earths on the Oxalate Content of Salicornia europaea L. Z. Für Pflanzenphysiol. 1978, 88, 403–412. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: Formation and function. Annu. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [PubMed]
- Nakata, P.A. Advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Sci. 2003, 164, 901–909. [Google Scholar]
- Van Oosten, M.J.; Maggio, A. Functional biology of halophytes in the phytoremediation of heavy metal contaminated soils. Environ. Exp. Bot. 2015, 111, 135–146. [Google Scholar]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014.
- Silva, H.; Caldeira, G.; Freitas, H. Salicornia ramosissima population dynamics and tolerance of salinity. Ecol. Res. 2007, 22, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Romero, J.A.; Duarte, B.; Barcia-Piedras, J.M.; Matos, A.R.; Redondo-Gómez, S.; Caçador, I.; Mateos-Naranjo, E. Investigating the physiological mechanisms underlying Salicornia ramosissima response to atmospheric CO2 enrichment under coexistence of prolonged soil flooding and saline excess. Plant Physiol. Biochem. 2019, 135, 149–159. [Google Scholar] [CrossRef]
- Pérez-Romero, J.A.; Idaszkin, Y.L.; Barcia-Piedras, J.M.; Duarte, B.; Redondo-Gómez, S.; Caçador, I.; Mateos-Naranjo, E. Disentangling the effect of atmospheric CO2 enrichment on the halophyte Salicornia ramosissima J. Woods physiological performance under optimal and suboptimal saline conditions. Plant Physiol. Biochem. 2018, 127, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Romero, J.A.; Barcia-Piedras, J.M.; Redondo-Gómez, S.; Mateos-Naranjo, E. Impact of short-term extreme temperature events on physiological performance of Salicornia ramosissima J. Woods under optimal and sub-optimal saline conditions. Sci. Rep. 2019, 9, 659. [Google Scholar] [CrossRef] [Green Version]
- Maciel, E.; Domingues, P.; Domingues, M.R.M.; Calado, R.; Lillebø, A. Halophyte plants from sustainable marine aquaponics are a valuable source of omega-3 polar lipids. Food Chem. 2020, 320, 126560. [Google Scholar] [CrossRef]
- Lima, A.R.; Castañeda-Loaiza, V.; Salazar, M.; Nunes, C.; Quintas, C.; Gama, F.; Pestana, M.; Correia, P.J.; Santos, T.; Varela, J.; et al. Influence of cultivation salinity in the nutritional composition, antioxidant capacity and microbial quality of Salicornia ramosissima commercially produced in soilless systems. Food Chem. 2020, 333, 127525. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Marín, J.; Mateos-Naranjo, E.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Redondo-Gómez, S. Synergic Effects of Rhizobacteria: Increasing Use of Halophytes in a Changing World. In Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; Hasanuzzaman, M., Shabala, S., Fujita, M., Eds.; CAB International: Wallingford, UK, 2019; pp. 240–254. [Google Scholar]
- Chebotar, V.K.; Chizhevskaya, E.P.; Baganova, M.E.; Keleinikova, O.V.; Yuzikhin, O.S.; Zaplatkin, A.N.; Khonina, O.V.; Kostitsin, R.D.; Lapenko, N.G. Endophytes from Halotolerant Plants Aimed to Overcome Salinity and Draught. Plants 2022, 11, 2992. [Google Scholar] [CrossRef] [PubMed]
- Carreiras, J.; Caçador, I.; Duarte, B. Bioaugmentation Improves Phytoprotection in Halimione portulacoides Exposed to Mild Salt Stress: Perspectives for Salinity Tolerance Improvement. Plants 2022, 11, 1055. [Google Scholar] [CrossRef]
- Mathew, B.T.; Torky, Y.; Amin, A.; Mourad, A.H.I.; Ayyash, M.M.; El-Keblawy, A.; Hilal-Alnaqbi, A.; AbuQamar, S.F.; El-Tarabily, K.A. Halotolerant Marine Rhizosphere-Competent Actinobacteria Promote Salicornia bigelovii Growth and Seed Production Using Seawater Irrigation. Front. Microbiol. 2020, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- El-Tarabily, K.A.; ElBaghdady, K.Z.; AlKhajeh, A.S.; Ayyash, M.M.; Aljneibi, R.S.; El-Keblawy, A.; AbuQamar, S.F. Polyamine-producing actinobacteria enhance biomass production and seed yield in Salicornia bigelovii. Biol. Fertil. Soils 2020, 56, 499–519. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; AlKhajeh, A.S.; Ayyash, M.M.; Alnuaimi, L.H.; Sham, A.; ElBaghdady, K.Z.; Tariq, S.; AbuQamar, S.F. Growth promotion of Salicornia bigelovii by Micromonospora chalcea UAE1, an endophytic 1-aminocyclopropane-1-carboxylic acid deaminase-producing actinobacterial isolate. Front. Microbiol. 2019, 10, 1694. [Google Scholar] [CrossRef]
- Bashan, Y.; Moreno, M.; Troyo, E. Growth promotion of the seawater-irrigated oilseed halophyte Salicornia bigelovii inoculated with mangrove rhizosphere bacteria and halotolerant Azospirillum spp. Biol. Fertil. Soils 2000, 32, 265–272. [Google Scholar] [CrossRef]
- Rueda-Puente, E.O.; Prabhaharan, R.; Murillo-Amador, B.; Ruiz-Espinoza, F.; Puente, M.; Valdez-Cepeda, R.D. Ameliorative effects of salt resistance on physiological parameters in the halophyte Salicornia bigelovii torr. with plant growth-promoting rhizobacteria. Afr. J. Biotechnol. 2013, 12, 5278–5284. [Google Scholar]
- Razzaghi Komaresofla, B.; Alikhani, H.A.; Etesami, H.; Khoshkholgh-Sima, N.A. Improved growth and salinity tolerance of the halophyte Salicornia sp. by co–inoculation with endophytic and rhizosphere bacteria. Appl. Soil Ecol. 2019, 138, 160–170. [Google Scholar] [CrossRef]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Bernabeu-Meana, M.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S. Effect of Plant Growth-Promoting Rhizobacteria on Salicornia ramosissima Seed Germination under Salinity, CO2 and Temperature Stress. Agronomy 2019, 9, 655. [Google Scholar] [CrossRef] [Green Version]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Impact of Plant Growth Promoting Bacteria on Salicornia ramosissima Ecophysiology and Heavy Metal Phytoremediation Capacity in Estuarine Soils. Front. Microbiol. 2020, 11, 553018. [Google Scholar] [CrossRef] [PubMed]
- Maia, M.; Cavaco, A.R.; Laureano, G.; Cunha, J.; Eiras-Dias, J.; Matos, A.R.; Duarte, B.; Figueiredo, A. More than just wine: The nutritional benefits of grapevine leaves. Foods 2021, 10, 2251. [Google Scholar] [CrossRef] [PubMed]
- Duarte, B.; Carreiras, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Matos, A.R.; Marques, J.C.; Caçador, I. Halophyte fatty acids as biomarkers of anthropogenic-driven contamination in Mediterranean marshes: Sentinel species survey and development of an integrated biomarker response (IBR) index. Ecol. Indic. 2018, 87, 86–96. [Google Scholar] [CrossRef]
- Guil, J.L.; Torija, M.E.; Giménez, J.J.; Rodríguez, I. Identification of fatty acids in edible wild plants by gas chromatography. J. Chromatogr. A 1996, 719, 229–235. [Google Scholar] [CrossRef]
- Anwar, F.; Bhanger, M.I.; Nasir, M.K.A.; Ismail, S. Analytical characterization of Salicornia bigelovii seed oil cultivated in Pakistan. J. Agric. Food Chem. 2002, 50, 4210–4214. [Google Scholar] [CrossRef]
- Isca, V.M.S.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, H.; Silva, A.M.S. Lipophilic profile of the edible halophyte Salicornia ramosissima. Food Chem. 2014, 165, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Radwan, H.; Nazif, N. Phytochemical Investigation of Salicornia fruticosa (L.) And Their Biological Activity. Res. J. Med. Med. Sci. 2007, 2, 72–78. [Google Scholar]
- Elsebaie, E.M.; Elsanat, S.Y.; Gouda, M.S.; Elnemr, K.M. Oil and Fatty Acids Composition in Glasswort (Salicornia Fruticosa) Seeds. IOSR J. Appl. Chem. 2013, 4, 6–9. [Google Scholar]
- Gounaris, K.; Barber, J. Isolation and characterisation of a photosystem II reaction centre lipoprotein complex. FEBS Lett. 1985, 188, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Sui, N.; Li, M.; Li, K.; Song, J.; Wang, B.S. Increase in unsaturated fatty acids in membrane lipids of Suaeda salsa L. enhances protection of photosystem II under high salinity. Photosynthetica 2010, 48, 623–629. [Google Scholar] [CrossRef]
- Duarte, B.; Cabrita, M.T.; Gameiro, C.; Matos, A.R.; Godinho, R.; Marques, J.C.; Caçador, I. Disentangling the photochemical salinity tolerance in Aster tripolium L.: Connecting biophysical traits with changes in fatty acid composition. Plant Biol. 2017, 19, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Mizusawa, N.; Wada, H. The role of lipids in photosystem II. Biochim. Biophys. Acta 2012, 1817, 194–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Ouariti, O.; Boussama, N.; Zarrouk, M.; Cherif, A.; Ghorbal, M.H. Cadmium- and copper-induced changes in tomato membrane lipids. Phytochemistry 1997, 45, 1343–1350. [Google Scholar] [CrossRef]
- Trémolières, A.; Roche, O.; Dubertret, G.; Guyon, D.; Garnier, J. Restoration of thylakoid appression by Δ3-trans-hexadecenoic acid-containing phosphatidylglycerol in a mutant of Chlamydomonas reinhardtii. Relationships with the regulation of excitation energy distribution. BBA—Bioenerg. 1991, 1059, 286–292. [Google Scholar] [CrossRef]
- Duarte, B.; Matos, A.R.; Marques, J.C.; Caçador, I. Leaf fatty acid remodeling in the salt-excreting halophytic grass Spartina patens along a salinity gradient. Plant Physiol. Biochem. 2018, 124, 112–116. [Google Scholar] [CrossRef]
- Antunes, M.D.; Gago, C.; Guerreiro, A.; Sousa, A.R.; Julião, M.; Miguel, M.G.; Faleiro, M.L.; Panagopoulos, T. Nutritional Characterization and Storage Ability of Salicornia ramosissima and Sarcocornia perennis for Fresh Vegetable Salads. Horticulturae 2021, 7, 6. [Google Scholar] [CrossRef]
- Duarte, B.; Santos, D.; Caçador, I. Halophyte anti-oxidant feedback seasonality in two salt marshes with different degrees of metal contamination: Search for an efficient biomarker. Funct. Plant Biol. 2013, 40, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Essaidi, I.; Brahmi, Z.; Snoussi, A.; Ben Haj Koubaier, H.; Casabianca, H.; Abe, N.; El Omri, A.; Chaabouni, M.M.; Bouzouita, N. Phytochemical investigation of Tunisian Salicornia herbacea L., antioxidant, antimicrobial and cytochrome P450 (CYPs) inhibitory activities of its methanol extract. Food Control 2013, 32, 125–133. [Google Scholar] [CrossRef]
- Kang, S.; Kim, M.R.; Chiang, M.; Hong, J. Evaluation and comparison of functional properties of freshwater-cultivated glasswort (Salicornia herbacea L.) with naturally-grown glasswort. Food Sci. Biotechnol. 2015, 24, 2245–2250. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, J.H. Optimization of ultrasound-assisted extraction of phenolic compounds from Salicornia herbacea powder. J. Food Sci. Nutr. 2009, 14, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Xu, J.Y.; Jia, W.; Chen, Z.; Xu, Z.C. Chloride salinity in a chloride-sensitive plant: Focusing on photosynthesis, hormone synthesis and transduction in tobacco. Plant Physiol. Biochem. 2020, 153, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Jahantigh, O.; Najafi, F.; Badi, H.N.; Khavari-Nejad, R.A.; Sanjarian, F. Changes in antioxidant enzymes activities and proline, total phenol and anthocyanine contents in Hyssopus officinalis L. plants under salt stress. Acta Biol. Hung. 2016, 67, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Sakalauskiene, S.; Sakalauskaite, J.; Lazauskas, S.; Povilaitis, V.; Auškalniene, O.; Pšibišauskiene, G.; Samuoliene, G.; Brazaityte, A.; Duchovskis, P. Interactive effects of elevated CO2, temperature and water stress on Apera spica-venti L. J. Food Agric. Environ. 2013, 11, 1121–1124. [Google Scholar]
- Bueno, M.; Lendínez, M.L.; Calero, J.; del Pilar Cordovilla, M. Salinity responses of three halophytes from inland saltmarshes of Jaén (southern Spain). Flora 2020, 266, 151589. [Google Scholar] [CrossRef]
- Muscolo, A.; Panuccio, M.R.; Piernik, A. Ecology, Distribution and Ecophysiology of Salicornia europaea L. In Sabkha Ecosystems Volume IV: Cash Crop Halophyte and Biodiversity Conservation; Khan, M.A., Böer, B., Öztürk, M., Zahran Al Abdessalaam, T., Clüsener-Godt, M., Gul, B., Eds.; Springer: New York, NY, USA, 2014; pp. 233–240. ISBN 978-94-007-7410-0. [Google Scholar]
- Ehsen, S.; Abideen, Z.; Rizvi, R.F.; Gulzar, S.; Aziz, I.; Gul, B.; Khan, M.A.; Ansari, R. Ecophysiological adaptations and anti-nutritive status of sustainable cattle feed Haloxylon stocksii under saline conditions. Flora 2019, 257, 151425. [Google Scholar] [CrossRef]
- Jothiramshekar, S.; Benjamin, J.J.; Krishnasamy, R.; Pal, A.K.; George, S.; Swaminathan, R.; Parida, A.K. Responses of selected C3 and C4 halophytes to elevated CO2 concentration under salinity. Curr. Sci. 2018, 115, 129–135. [Google Scholar] [CrossRef]
- Lv, C.; Wang, H.; Wang, J.; Ma, X.; Xia, C. The influence of elevated CO2 on Vallisneria spiralis, Radix auricularia, and their plant–herbivore interaction. Aquat. Ecol. 2019, 53, 137–150. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, H.; Sun, X.; Ding, J. Effects of elevated temperature on chemistry of an invasive plant, its native congener and their herbivores. J. Plant Ecol. 2022, 15, 450–460. [Google Scholar] [CrossRef]
- Sharma, H.C.; War, A.R.; Pathania, M.; Sharma, S.P.; Akbar, S.M.D.; Munghate, R.S. Elevated CO2 influences host plant defense response in chickpea against Helicoverpa armigera. Arthropod. Plant. Interact. 2016, 10, 171–181. [Google Scholar] [CrossRef]
- Atif, M.J.; Amin, B.; Ghani, M.I.; Ali, M.; Cheng, Z. Variation in morphological and quality parameters in garlic (Allium sativum L.) bulb influenced by different photoperiod, temperature, sowing and harvesting time. Plants 2020, 9, 155. [Google Scholar] [CrossRef] [Green Version]
- Alhdad, G.M.; Seal, C.E.; Al-Azzawi, M.J.; Flowers, T.J. The effect of combined salinity and waterlogging on the halophyte Suaeda maritima: The role of antioxidants. Environ. Exp. Bot. 2013, 87, 120–125. [Google Scholar] [CrossRef]
- Miyagi, A.; Noguchi, K.; Tokida, T.; Usui, Y.; Nakamura, H.; Sakai, H.; Hasegawa, T.; Kawai-Yamada, M. Oxalate contents in leaves of two rice cultivars grown at a free-air CO2 enrichment (FACE) site. Plant Prod. Sci. 2019, 22, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Brooks, G.L.; Whittaker, J.B. Responses of multiple generations of Gastrophysa viridula, feeding on Rumex obtusifolius, to elevated CO2. Glob. Chang. Biol. 1998, 4, 63–75. [Google Scholar] [CrossRef]
- Khan, M.A.; Aziz, S. Some aspects of salinity, plant density, and nutrient effects on Cressa cretica L. J. Plant Nutr. 1998, 21, 769–784. [Google Scholar] [CrossRef] [Green Version]
- Austenfeld, F.-A. Correlation of Substrate Salinity and Ion Concentration in Salicornia europaea L. with Special Reference to Oxalate. Biochem. Und Physiol. Der Pflanz. 1974, 165, 303–316. [Google Scholar] [CrossRef]
- Shi, R.; Liang, L.; Liu, W.; Zeb, A. Kochia scoparia L., a newfound candidate halophyte, for phytoremediation of cadmium-contaminated saline soils. Environ. Sci. Pollut. Res. 2022, 29, 44759–44768. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mesa-Marín, J.; Pérez-Romero, J.A.; López-Jurado, J.; García-López, J.V.; Mariscal, V.; Molina-Heredia, F.P.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Flowers, T.J.; et al. Consortia of plant-growth-promoting rhizobacteria isolated from halophytes improve response of eight crops to soil salinization and climate change conditions. Agronomy 2021, 11, 1609. [Google Scholar] [CrossRef]
- Forouzi, A.; Ghasemnezhad, A.; Nasrabad, R.G. Phytochemical response of Stevia plant to growth promoting microorganisms under salinity stress. S. Afr. J. Bot. 2020, 134, 109–118. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Iniguez, A.L.; Ahmer, B.M.M.; Triplett, E.W. Kinetics and strain specificity of rhizosphere and endophytic colonization by enteric bacteria on seedlings of Medicago sativa and Medicago truncatula. Appl. Environ. Microbiol. 2003, 69, 1783–1790. [Google Scholar] [CrossRef] [Green Version]
- Gamalero, E.; Glick, B.R. Bacterial Modulation of Plant Ethylene Levels. Plant Physiol. 2015, 169, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Nies, D.; Silver, S. Molecular Microbiology of Heavy Metals; Springer: Berlin/Heidelberg, Germany, 2007; ISBN 9783540697701. [Google Scholar]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Endophytic Cultivable Bacteria of the Metal Bioaccumulator Spartina maritima Improve Plant Growth but Not Metal Uptake in Polluted Marshes Soils. Front. Microbiol. 2015, 6, 1450. [Google Scholar] [CrossRef] [Green Version]
- Andrades-Moreno, L.; Del Castillo, I.; Parra, R.; Doukkali, B.; Redondo-Gómez, S.; Pérez-Palacios, P.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Prospecting metal-resistant plant-growth promoting rhizobacteria for rhizoremediation of metal contaminated estuaries using Spartina densiflora. Environ. Sci. Pollut. Res. Int. 2014, 21, 3713–3721. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Isolation of plant-growth-promoting and metal-resistant cultivable bacteria from Arthrocnemum macrostachyum in the Odiel marshes with potential use in phytoremediation. Mar. Pollut. Bull. 2016, 110, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Scouting contaminated estuaries: Heavy metal resistant and plant growth promoting rhizobacteria in the native metal rhizoaccumulator Spartina maritima. Mar. Pollut. Bull. 2015, 90, 150–159. [Google Scholar] [CrossRef]
- Midway, S.; Robertson, M.; Flinn, S.; Kaller, M. Comparing multiple comparisons: Practical guidance for choosing the best multiple comparisons test. PeerJ 2020, 8, e10387. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irrigation | Optimal (171 mmol NaCl L−1) | |||||||||||
| T and CO2 | 400 ppm | +4 °C, 700 ppm | ||||||||||
| Inoculation | C | 1 | 2 | 3 | 4 | 5 | C | 1 | 2 | 3 | 4 | 5 |
| SFA | 23.8 ± 0.5 | 25.7 ± 2.3 | 23.7 ± 0.4 | 24.3 ± 0.5 | 24.7 ± 0.3 | 24.4 ± 0.4 | 27.0 ± 0.3 | 25.8 ± 0.6 | 26.1 ± 0.6 | 25.7 ± 0.4 | 26.6 ± 1.6 | 26.1 ± 0.2 |
| MUFA | 8.6 ± 0.7 | 7.5 ± 1.1 | 10.0 ± 0.4 | 10.2 ± 0.6 | 10.5 ± 0.9 | 9.9 ± 0.3 | 10.6 ± 0.9 | 12.4 ± 1.0 | 11.4 ± 0.7 | 11.5 ± 0.9 | 15.9 ± 1.5 | 12.9 ± 0.7 |
| PUFA | 67.7 ± 0.7 | 66.8 ± 2.7 | 66.3 ± 0.6 | 65.5 ± 0.4 | 64.7 ± 0.8 | 65.7 ± 0.6 | 62.3 ± 1.2 | 61.8 ± 1.4 | 62.5 ± 1.0 | 62.8 ± 0.8 | 57.5 ± 3.0 | 61.0 ± 0.6 |
| PUFA/SFA | 2.9 ± 0.1 | 2.7 ± 0.3 | 2.8 ± 0.1 | 2.7 ± 0.1 | 2.6 ± 0.0 | 2.7 ± 0.1 | 2.3 ± 0.1 | 2.4 ± 0.1 | 2.4 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.2 | 2.3 ± 0.0 |
| 18:2/18:3 | 0.8 ± 0.04 | 0.7 ± 0.05 | 0.8 ± 0.04 | 0.9 ± 0.01 | 0.7 ± 0.04 | 0.8 ± 0.05 | 1.14 ± 0.12 | 0.87 ± 0.03 | 1.06 ± 0.07 | 1.00 ± 0.03 | 1.10 ± 0.06 | 1.27 ± 0.13 |
| Cox | 11.4 ± 0.2 | 11.4 ± 0.5 | 11.0 ± 0.2 | 10.7 ± 0.1 | 11.0 ± 0.1 | 10.9 ± 0.2 | 9.8 ± 0.3 | 10.2 ± 0.2 | 10.0 ± 0.2 | 10.1 ± 0.1 | 8.6 ± 0.9 | 9.5 ± 0.2 |
| OS | 5171.4 ± 100.5 | 5170.4 ± 245.0 | 5005.4 ± 87.3 | 4853.7 ± 28.6 | 5017.9 ± 70.4 | 4972.5 ± 89.7 | 4441.5 ± 158.5 | 4618.9 ± 113.0 | 4499.2 ± 105.0 | 4566.6 ± 47.4 | 3852.1 ± 444.8 | 4256.0 ± 106.2 |
| h/H | 3.4 ± 0.1 | 3.2 ± 0.2 | 3.5 ± 0.1 | 3.3 ± 0.1 | 3.3 ± 0.1 | 3.4 ± 0.1 | 2.9 ± 0.1 | 3.1 ± 0.1 | 3.0 ± 0.1 | 3.0 ± 0.1 | 3.0 ± 0.2 | 3.0 ± 0.0 |
| PI | 83.3 ± 2.1 | 83.7 ± 4.2 | 80.6 ± 1.9 | 76.8 ± 0.5 | 83.1 ± 1.6 | 80.2 ± 2.1 | 67.9 ± 3.3 | 75.1 ± 1.6 | 70.0 ± 2.3 | 71.9 ± 0.5 | 59.6 ± 8.8 | 65.4 ± 2.4 |
| AI | 0.30 ± 0.01 | 0.35 ± 0.05 | 0.31 ± 0.01 | 0.30 ± 0.01 | 0.30 ± 0.01 | 0.30 ± 0.01 | 0.37 ± 0.01 | 0.35 ± 0.01 | 0.35 ± 0.01 | 0.35 ± 0.01 | 0.36 ± 0.03 | 0.35 ± 0.00 |
| TI | 0.2 ± 0.01 | 0.19 ± 0.03 | 0.18 ± 0.01 | 0.19 ± 0.00 | 0.18 ± 0.00 | 0.2 ± 0.01 | 0.25 ± 0.01 | 0.21 ± 0.01 | 0.23 ± 0.01 | 0.22 ± 0.00 | 0.38 ± 0.14 | 0.25 ± 0.01 |
| Omega 3 | 38.5 ± 1.3 | 39.2 ± 2.4 | 36.6 ± 1.1 | 34.5 ± 0.3 | 38.1 ± 0.9 | 36.5 ± 1.3 | 29.6 ± 2.0 | 33.2 ± 1.0 | 30.5 ± 1.3 | 31.5 ± 0.4 | 22.7 ± 5.7 | 27.3 ± 1.5 |
| Omega 6 | 29.1 ± 0.8 | 27.5 ± 1.3 | 29.7 ± 0.6 | 31.1 ± 0.3 | 26.7 ± 1.0 | 29.2 ± 1.0 | 32.8 ± 1.3 | 28.7 ± 0.7 | 32.0 ± 1.0 | 31.3 ± 0.8 | 34.9 ± 2.9 | 33.7 ± 1.2 |
| Irrigation | Salinity (510 mmol NaCl L−1) | |||||||||||
| T and CO2 | 400 ppm | +4 °C, 700 ppm | ||||||||||
| Inoculation | C | 1 | 2 | 3 | 4 | 5 | C | 1 | 2 | 3 | 4 | 5 |
| SFA | 24.4 ± 0.6 | 23.9 ± 0.4 | 24.1 ± 0.4 | 24.8 ± 0.3 | 25.8 ± 1.6 | 25.5 ± 0.8 | 22.6 ± 2.5 | 25.9 ± 0.4 | 27.2 ± 0.9 | 24.9 ± 0.5 | 26.4 ± 0.2 | 26.1 ± 0.3 |
| MUFA | 9.2 ± 0.8 | 9.4 ± 0.5 | 9.9 ± 0.5 | 11.7 ± 0.3 | 11.3 ± 1.1 | 9.7 ± 1.7 | 21.5 ± 8.5 | 12.1 ± 0.5 | 13.4 ± 1.4 | 11.1 ± 0.8 | 14.4 ± 1.5 | 13.0 ± 1.0 |
| PUFA | 66.4 ± 1.3 | 66.7 ± 0.8 | 66.0 ± 0.7 | 63.6 ± 0.2 | 62.8 ± 2.6 | 64.8 ± 1.9 | 55.9 ± 6.0 | 62.0 ± 0.9 | 59.4 ± 2.3 | 64.1 ± 1.2 | 59.2 ± 1.6 | 60.9 ± 0.8 |
| PUFA/SFA | 2.7 ± 0.1 | 2.8 ± 0.1 | 2.7 ± 0.1 | 2.6 ± 0.0 | 2.5 ± 0.2 | 2.6 ± 0.1 | 2.5 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.1 | 2.6 ± 0.1 | 2.2 ± 0.1 | 2.3 ± 0.0 |
| 18:2/18:3 | 0.9 ± 0.1 | 0.8 ± 0.08 | 0.9 ± 0.03 | 0.9 ± 0.02 | 0.9 ± 0.10 | 0.8 ± 0.03 | 1.14 ± 0.10 | 0.91 ± 0.03 | 1.05 ± 0.11 | 0.99 ± 0.06 | 1.07 ± 0.08 | 0.99 ± 0.05 |
| Cox | 10.9 ± 0.3 | 11.1 ± 0.3 | 10.7 ± 0.1 | 10.4 ± 0.1 | 10.4 ± 0.6 | 10.8 ± 0.3 | 8.9 ± 0.9 | 10.1 ± 0.2 | 9.6 ± 0.5 | 10.3 ± 0.3 | 9.5 ± 0.4 | 9.8 ± 0.1 |
| OS | 4955.5 ± 157.5 | 5028.9 ± 131.9 | 4855.1 ± 69.3 | 4720.1 ± 39.4 | 4711.4 ± 281.8 | 4884.6 ± 138.0 | 3992.4 ± 444.1 | 4591.0 ± 73.6 | 4310.4 ± 236.7 | 4677.5 ± 140.1 | 4263.3 ± 171.3 | 4441.2 ± 44.8 |
| h/H | 3.3 ± 0.1 | 3.4 ± 0.1 | 3.4 ± 0.1 | 3.2 ± 0.1 | 3.1 ± 0.2 | 3.2 ± 0.1 | 4.0 ± 0.8 | 3.0 ± 0.1 | 2.8 ± 0.1 | 3.2 ± 0.1 | 2.9 ± 0.1 | 3.0 ± 0.1 |
| PI | 78.4 ± 3.1 | 80.5 ± 3.0 | 76.1 ± 1.2 | 75.9 ± 1.1 | 76.3 ± 5.1 | 78.2 ± 1.8 | 66.3 ± 4.3 | 74.0 ± 1.1 | 69.1 ± 3.8 | 73.6 ± 2.6 | 68.6 ± 2.5 | 71.1 ± 1.1 |
| AI | 0.32 ± 0.01 | 0.3 ± 0.01 | 0.3 ± 0.01 | 0.3 ± 0.01 | 0.3 ± 0.03 | 0.3 ± 0.01 | 0.30 ± 0.04 | 0.35 ± 0.01 | 0.37 ± 0.02 | 0.33 ± 0.01 | 0.36 ± 0.00 | 0.35 ± 0.00 |
| TI | 0.19 ± 0.01 | 0.2 ± 0.01 | 0.2 ± 0.01 | 0.2 ± 0.01 | 0.2 ± 0.03 | 0.2 ± 0.01 | 0.21 ± 0.02 | 0.22 ± 0.01 | 0.25 ± 0.03 | 0.21 ± 0.01 | 0.24 ± 0.01 | 0.23 ± 0.00 |
| Omega 3 | 35.6 ± 1.9 | 36.7 ± 1.8 | 34.1 ± 0.8 | 33.6 ± 0.5 | 34.0 ± 3.0 | 35.6 ± 1.1 | 26.5 ± 3.5 | 32.5 ± 0.7 | 29.5 ± 2.5 | 32.4 ± 1.6 | 28.8 ± 1.9 | 30.7 ± 0.6 |
| Omega 6 | 30.8 ± 1.1 | 30.0 ± 1.2 | 31.9 ± 0.6 | 29.9 ± 0.3 | 28.8 ± 0.6 | 29.1 ± 1.2 | 29.4 ± 3.2 | 29.5 ± 0.6 | 29.8 ± 0.6 | 31.6 ± 0.7 | 30.3 ± 0.5 | 30.2 ± 1.1 |
| Irrigation | Drought | |||||||||||
| T and CO2 | 400 ppm | +4 °C, 700 ppm | ||||||||||
| Inoculation | C | 1 | 2 | 3 | 4 | 5 | C | 1 | 2 | 3 | 4 | 5 |
| SFA | 24.2 ± 0.6 | 25.3 ± 1.2 | 24.8 ± 0.1 | 28.5 ± 2.4 | 24.9 ± 0.3 | 24.4 ± 0.5 | 26.0 ± 0.5 | 25.9 ± 0.7 | 26.8 ± 0.6 | 26.6 ± 0.7 | 26.3 ± 0.5 | 25.5 ± 0.5 |
| MUFA | 9.1 ± 0.6 | 8.4 ± 0.6 | 10.8 ± 1.0 | 10.7 ± 1.4 | 9.6 ± 0.4 | 10.7 ± 0.8 | 9.6 ± 0.7 | 12.4 ± 1.4 | 9.1 ± 1.3 | 9.3 ± 0.5 | 13.3 ± 0.5 | 11.5 ± 0.9 |
| PUFA | 66.7 ± 0.9 | 66.3 ± 1.6 | 64.4 ± 1.0 | 60.9 ± 3.6 | 65.5 ± 0.7 | 64.9 ± 1.1 | 64.4 ± 0.9 | 61.7 ± 2.1 | 51.0 ± 3.3 | 64.0 ± 1.0 | 60.4 ± 0.9 | 63.0 ± 1.3 |
| PUFA/SFA | 2.8 ± 0.1 | 2.7 ± 0.2 | 2.6 ± 0.1 | 2.2 ± 0.3 | 2.6 ± 0.1 | 2.7 ± 0.1 | 2.5 ± 0.1 | 2.4 ± 0.1 | 1.9 ± 0.2 | 2.4 ± 0.1 | 2.3 ± 0.1 | 2.5 ± 0.1 |
| 18:2/18:3 | 0.9 ± 0.03 | 0.8 ± 0.07 | 0.9 ± 0.06 | 0.9 ± 1.16 | 0.7 ± 0.04 | 0.8 ± 0.05 | 1.04 ± 0.10 | 1.15 ± 0.17 | 2.01 ± 0.25 | 1.08 ± 0.15 | 1.46 ± 0.12 | 1.03 ± 0.09 |
| Cox | 11.0 ± 0.2 | 11.2 ± 0.4 | 10.6 ± 0.3 | 9.8 ± 1.1 | 11.1 ± 0.1 | 10.9 ± 0.3 | 10.3 ± 0.1 | 9.8 ± 0.5 | 7.3 ± 0.6 | 10.2 ± 0.3 | 9.1 ± 0.3 | 10.1 ± 0.4 |
| OS | 4988.5 ± 89.1 | 5069.5 ± 190.4 | 4819.3 ± 136.3 | 4459.0 ± 503.6 | 5026.3 ± 59.1 | 4951.7 ± 131.6 | 4654.4 ± 63.4 | 4415.0 ± 248.2 | 3284.4 ± 295.5 | 4620.1 ± 166.6 | 4092.5 ± 123.9 | 4571.8 ± 174.8 |
| h/H | 3.3 ± 0.1 | 3.2 ± 0.1 | 3.3 ± 0.0 | 2.7 ± 0.3 | 3.2 ± 0.0 | 3.3 ± 0.1 | 3.0 ± 0.1 | 3.0 ± 0.1 | 2.4 ± 0.2 | 2.9 ± 0.1 | 3.0 ± 0.1 | 3.0 ± 0.1 |
| PI | 78.9 ± 1.5 | 81.5 ± 3.9 | 77.6 ± 2.4 | 70.4 ± 10.7 | 81.9 ± 1.3 | 81.0 ± 2.5 | 71.3 ± 2.2 | 69.2 ± 4.4 | 43.2 ± 5.9 | 70.6 ± 3.9 | 61.4 ± 2.5 | 71.9 ± 3.5 |
| AI | 0.3 ± 0.01 | 0.3 ± 0.02 | 0.3 ± 0.00 | 0.4 ± 0.05 | 0.3 ± 0.01 | 0.3 ± 0.01 | 0.35 ± 0.01 | 0.35 ± 0.01 | 0.46 ± 0.04 | 0.36 ± 0.01 | 0.36 ± 0.01 | 0.34 ± 0.01 |
| TI | 0.2 ± 0.01 | 0.2 ± 0.02 | 0.2 ± 0.01 | 0.3 ± 0.12 | 0.2 ± 0.00 | 0.2 ± 0.01 | 0.22 ± 0.01 | 0.24 ± 0.03 | 0.38 ± 0.04 | 0.23 ± 0.02 | 0.27 ± 0.02 | 0.22 ± 0.01 |
| Omega 3 | 36.0 ± 0.9 | 37.7 ± 2.2 | 34.7 ± 1.7 | 31.1 ± 6.3 | 37.6 ± 0.8 | 36.7 ± 1.6 | 31.8 ± 1.2 | 29.6 ± 2.9 | 17.8 ± 2.7 | 31.5 ± 2.3 | 24.8 ± 1.5 | 31.4 ± 2.2 |
| Omega 6 | 30.7 ± 0.4 | 28.6 ± 0.8 | 29.6 ± 0.7 | 29.8 ± 2.8 | 27.9 ± 0.8 | 28.2 ±0.9 | 32.6 ± 1.7 | 32.1 ± 1.0 | 33.2 ± 0.8 | 32.6 ± 1.5 | 35.6 ± 0.7 | 31.6 ± 1.0 |
| Treatment | FA 16:0 | FA 18:0 | FA 16:1t | FA 18:1 | FA 18:2 | FA 18:3 | Phenols | Oxalate |
|---|---|---|---|---|---|---|---|---|
| T/CO2 | 0.0000 *** | 0.0590 | 0.0000 *** | 0.0000 *** | 0.0000 *** | 0.0000 *** | 0.4109 | 0.0000 *** |
| Irrigation | 0.1267 | 0.7633 | 0.0000 *** | 0.1763 | 0.2043 | 0.0221 ** | 0.0000 *** | 0.0000 *** |
| Inoculation | 0.5454 | 0.3962 | 0.5359 | 0.0315 ** | 0.2587 | 0.0869 | 0.0966 | 0.0000 *** |
| T/CO2 × Irrigation | 0.4179 | 0.0871 | 0.0005 *** | 0.0262 ** | 0.0037 ** | 0.1464 | 0.0000 *** | 0.3307 |
| T/CO2 × Inoculation | 0.4104 | 0.6916 | 0.0002 *** | 0.0740 | 0.0011 ** | 0.0137 ** | 0.0075 ** | 0.0000 *** |
| Irrigation × Inoculation | 0.1679 | 0.3482 | 0.1805 | 0.4864 | 0.5135 | 0.1859 | 0.0000 *** | 0.0000 *** |
| T/CO2 × Irrigation × Inoculation | 0.1088 | 0.7653 | 0.0174 * | 0.3431 | 0.7529 | 0.0858 | 0.0000 *** | 0.0000 *** |
| CO2 and Temperature | Irrigation | Rhizobacteria Inoculation |
|---|---|---|
| 400 ppm CO2 25/14 °C | Optimal salinity (171 mmol L−1 NaCl) | None, 1, 2, 3, 4, and 5 |
| Salt stress (510 mmol L−1 NaCl) | None, 1, 2, 3, 4, and 5 | |
| Drought | None, 1, 2, 3, 4, and 5 | |
| 700 ppm CO2 29/18 °C | Optimal salinity (171 mmol L−1 NaCl) | None, 1, 2, 3, 4, and 5 |
| Salt stress (510 mmol L−1 NaCl) | None, 1, 2, 3, 4, and 5 | |
| Drought | None, 1, 2, 3, 4, and 5 |
| Consortia | Halophyte | Estuary and Year | Rhizobacteria Strain | Plant-Growth-Promoting Traits | Salt Tolerance (M NaCl) | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N Fixation | P Solubilization | Siderophores | IAA | Biofilm | ACC Deaminase | ||||||
| 1 | Spartina densiflora | Tinto 2010 | Pseudomonas composti SDT3 | • | • | 1.5 | [83] | ||||
| Aeromonas aquariorum SDT13 | • | • | • | 0.5 | |||||||
| Bacillus thuringiensis SDT14 | • | • | 1 | ||||||||
| 2 | Arthrocnemum macrostachyum | Odiel 2015 | Vibrio kanaloae RA1 | • | • | • | 0.6 | [84] | |||
| Pseudoalteromonas prydzensis RA15 | • | • | • | 1 | |||||||
| Staphylococcus warneri RA18 | • | 1.5 | |||||||||
| 3 | Spartina maritima | Tinto 2013 | Bacillus methylotrophicus SMT38 | • | • | • | 2 | [85] | |||
| Bacillus aryabhattai SMT48 | • | • | • | • | 1.5 | ||||||
| Bacillus licheniformis SMT51 | • | • | • | • | • | 2 | |||||
| 4 | Atriplex portulacoides | Piedras 2017 | Vibrio spartinae HPJ2 | • | • | • | • | • | • | 1 | [37] |
| Marinobacter sediminum HPJ15 | • | • | 1.7 | ||||||||
| Vibrio parahaemolyticus HPJ50 | • | • | • | • | 1 | ||||||
| 5 | Salicornia ramosissima | Tinto 2016 | Vibrio neocaledonicus SRT1 | • | • | • | • | • | 1.7 | [37] | |
| Thalassospira australica SRT8 | • | • | 1 | ||||||||
| Pseudarthrobacter oxydans SRT15 | • | • | • | 1 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesa-Marín, J.; Mateos-Naranjo, E.; Carreiras, J.; Feijão, E.; Duarte, B.; Matos, A.R.; Betti, M.; Del Rio, C.; Romero-Bernal, M.; Montaner, J.; et al. Interactive Temperature and CO2 Rise, Salinity, Drought, and Bacterial Inoculation Alter the Content of Fatty Acids, Total Phenols, and Oxalates in the Edible Halophyte Salicornia ramosissima. Plants 2023, 12, 1395. https://doi.org/10.3390/plants12061395
Mesa-Marín J, Mateos-Naranjo E, Carreiras J, Feijão E, Duarte B, Matos AR, Betti M, Del Rio C, Romero-Bernal M, Montaner J, et al. Interactive Temperature and CO2 Rise, Salinity, Drought, and Bacterial Inoculation Alter the Content of Fatty Acids, Total Phenols, and Oxalates in the Edible Halophyte Salicornia ramosissima. Plants. 2023; 12(6):1395. https://doi.org/10.3390/plants12061395
Chicago/Turabian StyleMesa-Marín, Jennifer, Enrique Mateos-Naranjo, João Carreiras, Eduardo Feijão, Bernardo Duarte, Ana Rita Matos, Marco Betti, Carmen Del Rio, Marina Romero-Bernal, Joan Montaner, and et al. 2023. "Interactive Temperature and CO2 Rise, Salinity, Drought, and Bacterial Inoculation Alter the Content of Fatty Acids, Total Phenols, and Oxalates in the Edible Halophyte Salicornia ramosissima" Plants 12, no. 6: 1395. https://doi.org/10.3390/plants12061395