

Seed Transmission of Begomoviruses: A Potential Threat for Bitter Gourd Cultivation
Abstract
:1. Introduction
2. Results
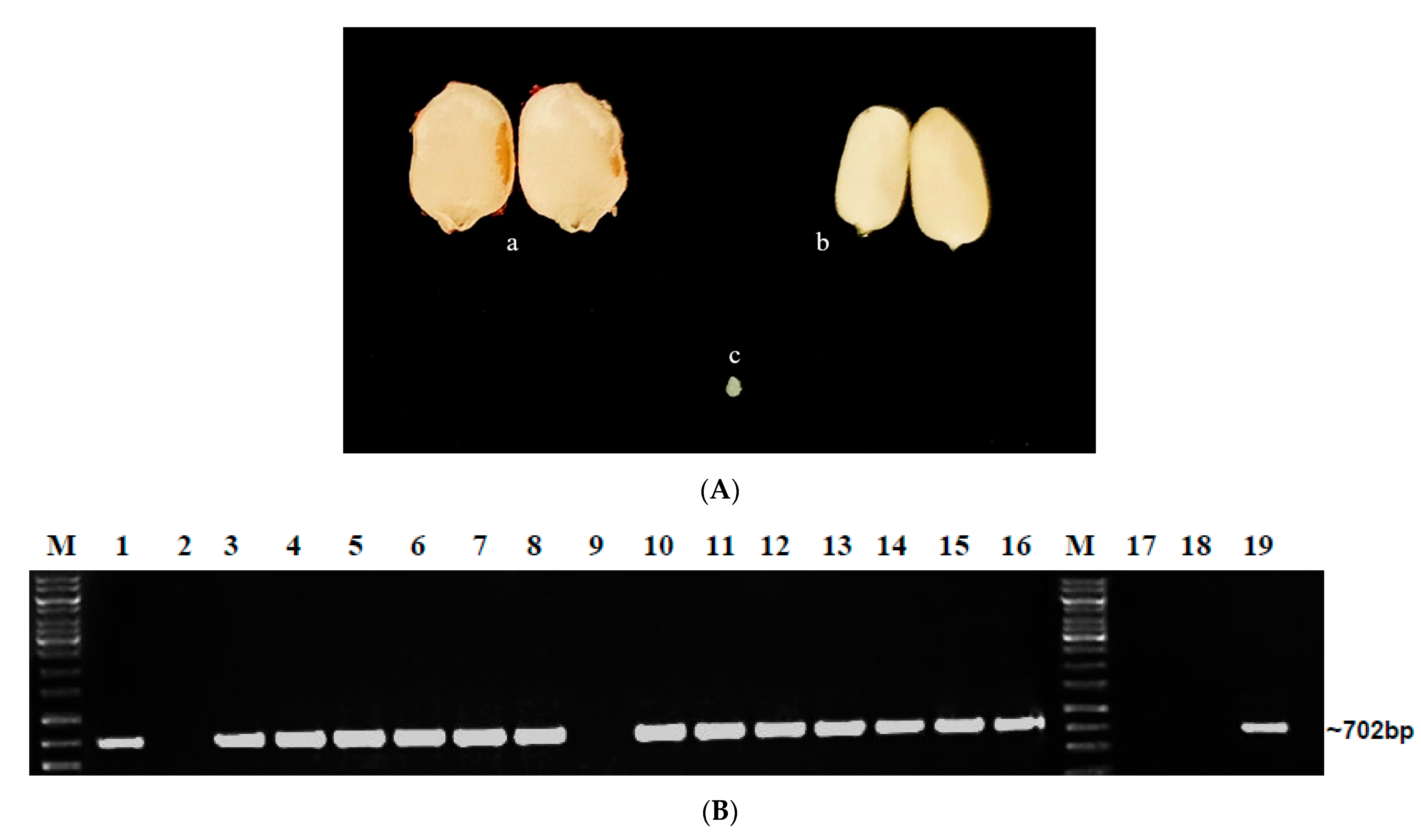
2.1. Detection of Begomoviruses in Market Seeds
2.1.1. DAS-ELISA
2.1.2. PCR Using ToLCNDV and BgYMV Specific Primers
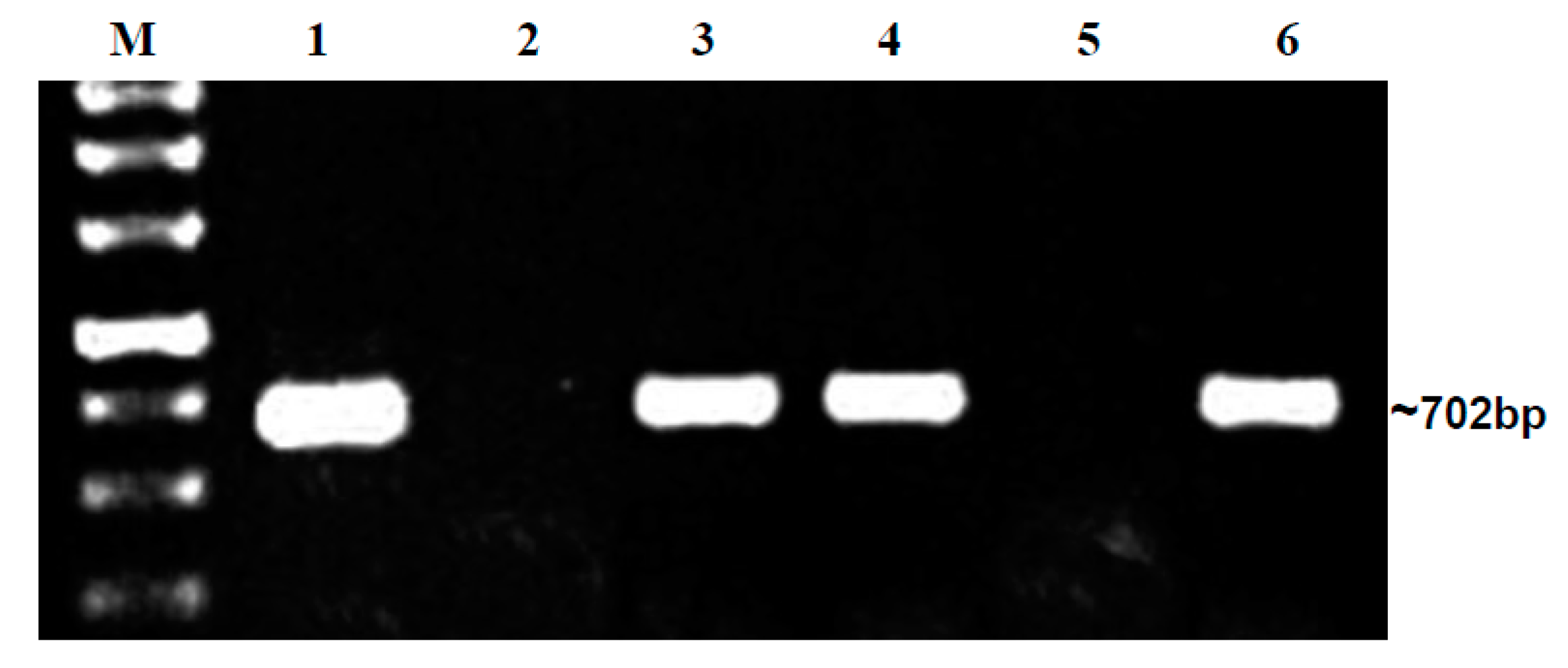
2.2. Detection of Begomoviruses in Seeds Collected from Infected Bitter Gourd Plants in the Field
2.2.1. DAS-ELISA
2.2.2. PCR Using BgYMV- and ToLCNDV-Specific Primers
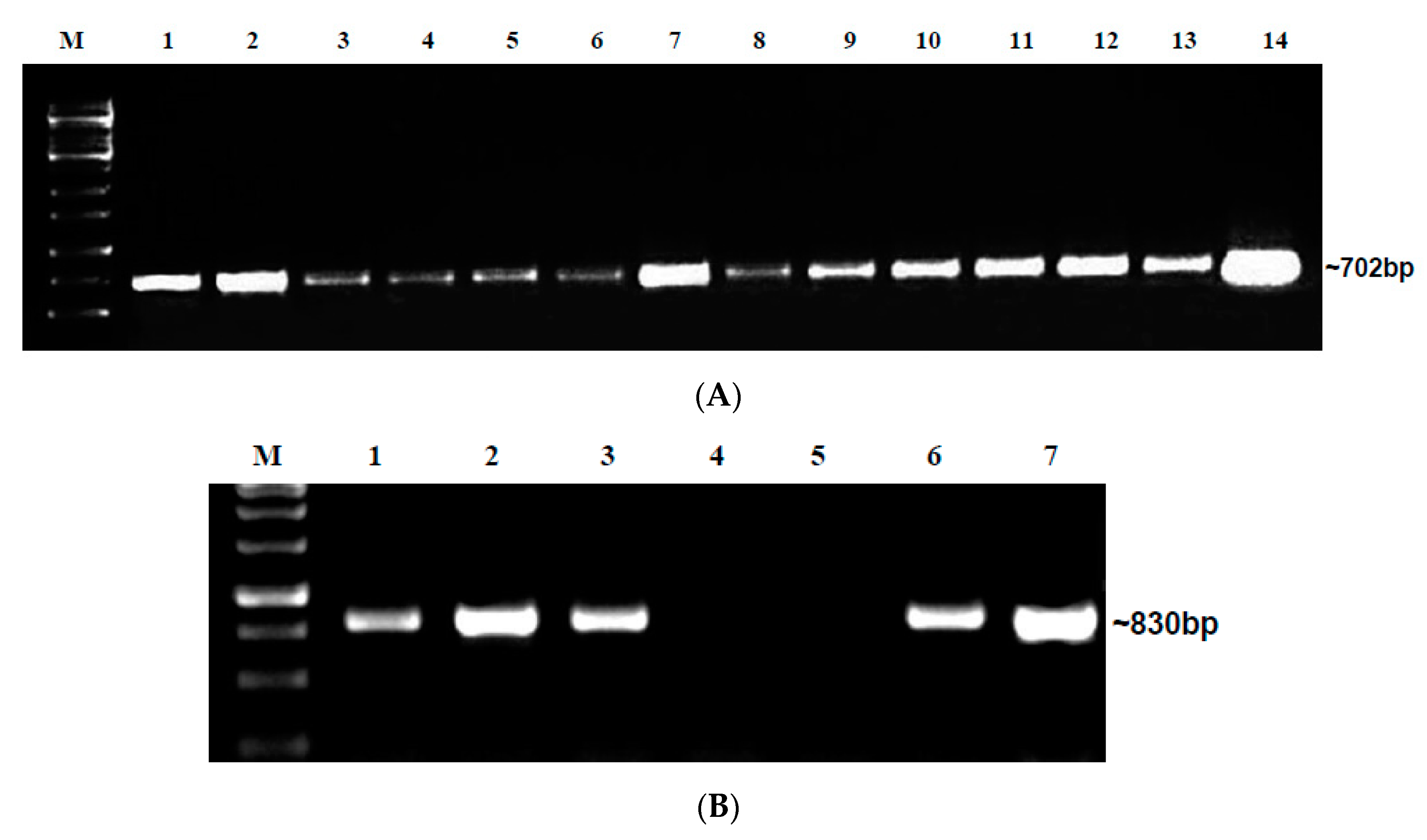
2.3. Assessing Seed Transmission by Grow-Out Test for Market Seeds in Glasshouse
2.4. Detection of Begomovirus in Grow-Out Test Plants in Microplot Study I
2.5. Tracking of Begomoviruses in Floral and Fruit Parts of Grow-Out Test Plants in Microplot through PCR
Grow-Out Test Plants Derived from Market and Field-Collected Seeds
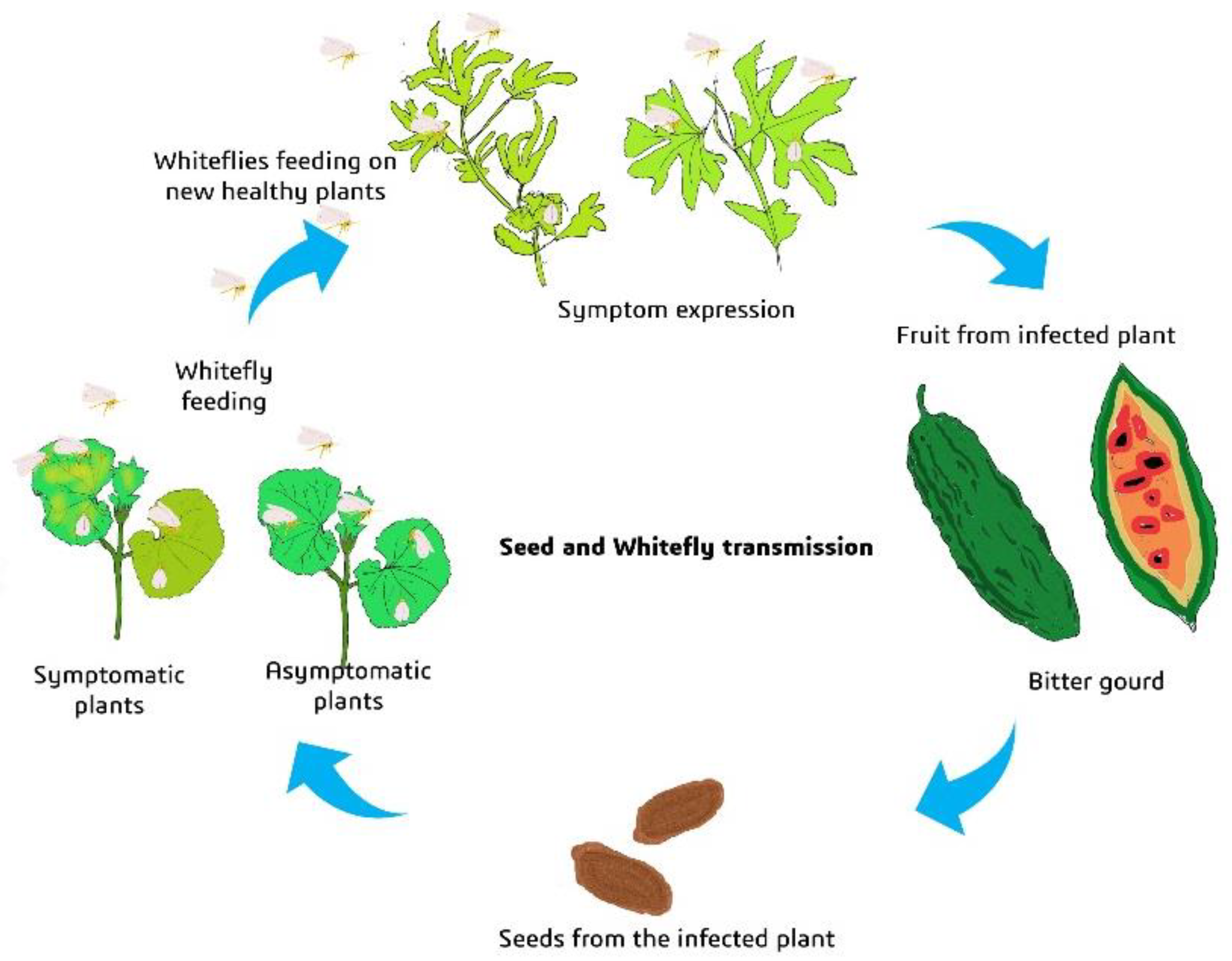
2.6. Transmissibility of Begomovirus from Grow-Out Test Plants of Microplot by Whiteflies
2.7. Seed as Potential Source of Inoculum for Active Spread of the Disease by Whitefly (Microplot Study II)
3. Discussion
4. Materials and Methods
4.1. Detection of Begomoviruses in Different Seed Parts through DAS-ELISA and PCR
4.1.1. Collection of Market Seeds
4.1.2. Collection of Seeds from Infected Fields
4.1.3. DAS-ELISA
4.1.4. PCR Using BgYMV and ToLCNDV Primers
4.2. Grow-Out Test for Seed Transmission
4.3. Detection of Begomovirus in Grow-Out Test Plants in Microplot
4.4. Tracking of Begomoviruses in Floral and Fruit Parts in Microplot
4.5. Transmissibility of Viral Inoculum from Grow-Out Test Plants of Microplot
4.5.1. Establishment of Whitefly Culture
4.5.2. Whitefly Transmission
4.6. Seed as Potential Source of Inoculum for Whitefly (Microplot Study II)
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Varsani, A.; Roumagnac, P.; Fuchs, M.; Navas-Castillo, J.; Moriones, E.; Idris, A.; Briddon, R.W.; Rivera-Bustamante, R.; Zerbini, F.M.; Martin, D.P. Capulavirus and Grablovirus: Two new genera in the family Geminiviridae. Arch. Virol. 2017, 162, 1819–1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch. Virol. 2020, 162, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A.; Consortium, I.R. ICTV virus taxonomy profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131. [Google Scholar] [CrossRef]
- Roumagnac, P.; Lett, J.M.; Fiallo-Olivé, E.; Navas-Castillo, J.; Zerbini, F.M.; Martin, D.P.; Varsani, A. Establishment of five new genera in the family Geminiviridae: Citlodavirus, Maldovirus, Mulcrilevirus, Opunvirus, and Topilevirus. Arch. Virol. 2022, 167, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.K.; Sharma, P.K.; Khan, M.S.; Snehi, S.K.; Raj, S.K.; Rao, G.P. Molecular detection and identification of Tomato leaf curl New Delhi virus isolate causing yellow mosaic disease in Bitter gourd (Momordica charantia), a medicinally important plant in India. Med. Plants 2010, 2, 117–123. [Google Scholar] [CrossRef]
- Raj, S.; Snehi, S.; Khan, M.; Tiwari, A.; Rao, G. First report of Pepper leaf curl Bangladesh virus strain associated with bitter gourd (Momordica charantia L.) yellow mosaic disease in India. Australas. Plant Dis. Notes 2010, 5, 14–16. [Google Scholar] [CrossRef] [Green Version]
- Nagendran, K.; Mohan Kumar, S.; Manoranjitham, S.K.; Karthikeyan, G. Molecular detection and characterization of Tomato leaf curl New Delhi virus causing mosaic disease on bitter gourd in Tamil Nadu, India. Biosci. Trends 2014, 7, 3925–3931. [Google Scholar]
- Naik, S.O.; Venkataravanappa, V.; Chakravarthy, A.K.; Reddy, C.L.; Krishnareddy, M. Survey, host range and characterization of begomovirus infecting bitter gourd and its vector whitefly cryptic species. Indian Phytopathol. 2022, 75, 1111–1127. [Google Scholar] [CrossRef]
- Manivannan, K.; Renukadevi, P.; Malathi, V.G.; Karthikeyan, G.; Balakrishnan, N. A new seed-transmissible begomovirus in bitter gourd (Momordica charantia L.). Microb. Pathog. 2019, 128, 82–89. [Google Scholar] [CrossRef]
- Renukadevi, P.; Sangeetha, B.; Malathi, V.G.; Nakkeeran, S.; Satya, V.K. Satya. Enigmatic emergence of seed transmission of geminiviruses. In Geminivirus: Detection, Diagnosis and Management; Academic Press: Cambridge, MA, USA, 2022; pp. 285–306. [Google Scholar]
- Kim, J.; Kil, E.J.; Kim, S.; Seo, H.; Byun, H.S.; Park, J.; Chung, M.N.; Kwak, H.R.; Kim, M.K.; Kim, C.S.; et al. Seed transmission of Sweet potato leaf curl virus in sweet potato (Ipomoea batatas). Plant Pathol. 2015, 64, 1284–1291. [Google Scholar] [CrossRef]
- Kil, E.J.; Park, J.; Choi, H.S.; Kim, C.S.; Lee, S. Lee. Seed transmission of Tomato yellow leaf curl virus in white soybean (Glycine max). Plant Pathol. J. 2017, 33, 424–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kil, E.J.; Park, J.; Choi, E.Y.; Byun, H.S.; Lee, K.Y.; An, C.G.; Lee, J.H.; Lee, G.S.; Choi, H.S.; Kim, C.S.; et al. Seed transmission of Tomato yellow leaf curl virus in sweet pepper (Capsicum annuum). Eur. J. Plant Pathol. 2018, 150, 759–764. [Google Scholar] [CrossRef]
- Kothandaraman, S.V.; Devadason, A.; Ganesan, M.V. Seed-borne nature of a begomovirus, Mung bean yellow mosaic virus in black gram. Appl. Microbial. Biotechnol. 2016, 100, 1925–1933. [Google Scholar] [CrossRef]
- Sangeetha, B.; Malathi, V.; Alice, D.; Suganthy, M.; Renukadevi, P. A distinct seed-transmissible strain of tomato leaf curl New Delhi virus infecting Chayote in India. Virus Res. 2018, 258, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.J.; Vo, T.T.B.; Fadhila, C.; Ho, P.T.; Lal, A.; Troiano, E.; Parrella, G.; Lee, S. Seed transmission of tomato leaf curl New Delhi virus from zucchini squash in Italy. Plant 2020, 9, 563. [Google Scholar] [CrossRef]
- Suruthi, V.; Nakkeeran, S.; Renukadevi, P.; Malathi, V.G.; Rajasree, V. Evidence of seed transmission of dolichos yellow mosaic virus, a begomovirus infecting lablab-bean in India. VirusDisease 2018, 29, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Zhang, Z.; Zhao, X.; Wang, Y.; Wang, S.; Qin, Y.; Zhang, D.; Tian, Y.; Zhao, F. Evidence for seed transmission of sweet potato symptomless virus 1 in sweet potato (Ipomoeabatatas). J. Plant Pathol. 2020, 102, 299–303. [Google Scholar] [CrossRef]
- Fadhila, C.; Lal, A.; Vo, T.T.; Ho, P.T.; Hidayat, S.H.; Lee, J.; Kil, E.J.; Lee, S. The threat of seed-transmissible pepper yellow leaf curl Indonesia virus in chili pepper. Microb. Pathog. 2020, 143, 104–132. [Google Scholar] [CrossRef]
- Ortega-Acosta, C.; Ochoa-Martínez, D.L.; Hernández-Morales, J.; Ramírez-Rojas, S.; Gutiérrez-Gallegos, J.A. Evaluation of seed transmission of begomoviruses in roselle and roselle-associated weeds. Rev. Mex. Fitopatol. 2019, 37, 135–146. [Google Scholar]
- Pagán, I. Transmission through seeds: The unknown life of plant viruses. PLoS Pathog. 2022, 18, 1010707. [Google Scholar] [CrossRef]
- Hull, R. Plant Virology; Elsevier: London, UK, 2014; pp. 1–979. [Google Scholar]
- Quainoo, A.; Wetten, A.; Allainguillaume, J. Transmission of cocoa swollen shoot virus by seeds. J. Virol. Methods 2008, 150, 45–49. [Google Scholar] [CrossRef]
- Keur, J.Y. Studies of the occurrence and transmission of virus diseases in the genus Abutilon. Bull. Torrey Bot. Club 1934, 61, 53–70. [Google Scholar] [CrossRef]
- Bennett, C.W.; Esau, K. Further studies on the relation of the curly top virus to plant tissues. J. Agric. Res. 1936, 13, 595–620. [Google Scholar]
- Andreason, S.A.; Olaniyi, O.G.; Gilliard, A.C.; Wadl, P.A.; Williams, L.H.; Jackson, D.M.; Simmons, A.M.; Ling, K.S. Large-scale seedling grow-out experiments do not support seed transmission of sweet potato leaf curl virus in sweet potato. Plants 2021, 10, 139. [Google Scholar] [CrossRef]
- Kil, E.-J.; Kim, S.; Lee, Y.-J.; Byun, H.-S.; Park, J.; Seo, H.; Kim, C.-S.; Shim, J.-K.; Lee, J.-H.; Kim, J.-K. Tomato yellow leaf curl virus (TYLCV-IL): A seed-transmissible geminivirus in tomatoes. Sci. Rep. 2016, 6, 19013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Padilla, V.; Fortes, I.M.; Romero-Rodríguez, B.; Arroyo-Mateos, M.; Castillo, A.G.; Moyano, C.; De León, L.; Moriones, E. Revisiting Seed Transmission of the Type Strain of Tomato yellow leaf curl virus in Tomato Plants. Phytopathology 2020, 110, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Fortes, I.; Pérez-Padilla, V.; Romero-Rodríguez, B.; Fernandez-Muñoz, R.; Moyano, C.; Castillo, A.G.; De León, L.; Moriones, E. The begomovirus tomato leaf curl New Delhi virus is seed-borne but not seed-transmitted in melon. Plant Dis. 2022, 107. [Google Scholar] [CrossRef]
- Chang, H.H.; Ku, H.M.; Tsai, W.S.; Chien, R.C.; Jan, F.J. Identification and characterization of a mechanical transmissible begomovirus causing leaf curl on oriental melon. Eur. J. Plant Pathol. 2010, 127, 219–228. [Google Scholar] [CrossRef]
- Chellappan, P.; Masona, M.; Vanitharani, R.; Taylor, N.; Fauquet, C. Broad spectrum resistance to ssDNA viruses associated with transgene-induced gene silencing in cassava. Plant Mol. Biol. 2004, 56, 601–611. [Google Scholar] [CrossRef]
- Wang, D.; Maule, A.J. Early embryo invasion as a determinant in pea of the seed transmission of pea seed-borne mosaic virus. J. Gen. Virol. 1992, 73, 1615–1620. [Google Scholar] [CrossRef]
- Wang, D.; Maule, A.J. Maule.A model for seed transmission of a plant virus: Genetic and structural analyses of pea embryo invasion by pea seed-borne mosaic virus. Plant Cell 1994, 6, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Maule, A.J. Contrasting patterns in the spread of two seed-borne viruses in pea embryos. Plant J. 1997, 11, 1333–1340. [Google Scholar] [CrossRef]
- Karmann, J.; Müller, B.; Hammes, U.Z. The long and winding road: Transport pathways for amino acids in Arabidopsis seeds. Plant Reprod. 2018, 31, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Rouhibakhsh, A.; Priya, J.; Periasamy, M.; Haq, Q.M.I.; Malathi, V.G. An improved DNA isolation method and PCR protocol for efficient detection of multicomponents of begomovirus in legumes. J. Virol. Methods 2008, 147, 37–42. [Google Scholar] [CrossRef]
- Virk, J.K.; Mandal, B.; Rao, V.R.; Kundu, T. Detection of Tomato Leaf Curl New Delhi Virus DNA Using U-Bent Optical Fiber-Based Lspr Probes. Opt. Fiber Technol. 2022, 74. [Google Scholar] [CrossRef]
- Zeidan, M.; Czosnek, H. Acquisition of tomato yellow leaf curl virus by the whitefly Bemisia tabaci. J. Gen. Virol. 1991, 72, 2607–2614. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample. No | Hybrid/Variety | Percentage of Embryo Infection (%) |
|---|---|---|
| Hybrid/variety seeds collected from market | ||
| 1. | H1 | 26.6 |
| 2. | H2 | 62.96 |
| 3. | H3 | 20.00 |
| 4. | H4 | 10.00 |
| 5. | Variety Co1 | 0.00 |
| Hybrid seeds collected from begomoviruses-infected plant from field | ||
| 6. | H1 | 26.6 |
| 7. | H2 | 33.33 |
| 8. | H3 | 20.0 |
| Sample. No | Hybrid/Variety | No of Samples Positive/Total No of Samples | |
|---|---|---|---|
| ToLCNDV Alone | Both BgYMV and ToLCNDV | ||
| Hybrid seeds from market | |||
| 1. | H1 | 4/8 | 2/8 |
| 2. | H2 | 13/17 | 4/17 |
| 3. | H3 | 6/6 | - |
| 4. | H4 | 3/3 | - |
| Hybrid seeds collected from begomoviruses-infected field | |||
| 5. | H1 | 3/4 | 1/4 |
| 6. | H2 | 3/5 | 2/5 |
| 7. | H3 | 3/3 | - |
| Sample. No | Hybrid | No of Samples Positive/Total No of Samples Tested | No of Symptomatic Plants | No of Asymptomatic Plants | Percentage of Seed Transmission (%) | |
|---|---|---|---|---|---|---|
| BgYMV | ToLCNDV | |||||
| 1 | H1 | 0/100 | 5/100 | 4/5 | 1/5 | 5.0 |
| 2 | H2 | 0/100 | 5/100 | 3/5 | 2/5 | 5.0 |
| 3 | H3 | 0/100 | 3/100 | 3/3 | 0/3 | 3.0 |
| Detection of Begomoviruses Using PCR with Specific Primers (170 Plants) | Percentage of Seed Transmission of Both Combined (%) | ||||
|---|---|---|---|---|---|
| ToLCNDV Alone Detected (%) | Percentage of Total Seed Transmission of ToLCNDV Alone (%) | Both BgYMV and ToLCNDV Detected (%) | |||
| Symptomatic Plants | Asymptomatic Plants | Symptomatic Plants | Asymptomatic Plants | ||
| Grow-out test plants (H1) of seeds collected from market | |||||
| 4.8 | 38.4 | 43.2 | 3.2 | 0.8 | 4.0 |
| Grow-out test plants (H1) of seeds derived from infected fields | |||||
| 2.2 | 0.0 | 2.2 | 6.6 | 2.2 | 8.8 |
| Inoculum Source (Microplot Study) | Acquisition Access Period (AAP) | Inoculation Feeding Period (IAP) | No of Plants Showing Symptoms/No of Plants Used for Transmission | Days Taken to Express Symptoms | Type of Symptoms | PCR Reaction |
|---|---|---|---|---|---|---|
| Symptomatic plants raised from market seeds | ||||||
| 1 | 24 h | 12 h | 2/15 | 20 | Mosaic and mottling | + |
| Asymptomatic plants raised from market seeds | ||||||
| 2 | 24 h | 12 h | 3/15 | 20 | Puckering, crinkling and mottling | + |
| Healthy plants | ||||||
| 3 | 24 h | 12 h | 0/15 | Nil | No symptom | − |
| Particulars | No of Samples Positive/No of Samples Tested in PCR BgYMV | No of Samples Positive/No of Samples Tested in PCR ToLCNDV | Extent of Transmission (%) |
|---|---|---|---|
| Before releasing of whitefly in microplot | 22/150 | 43/150 | 43.33 |
| After releasing whitefly in microplot | +14/150 | +29/150 | +28.66 |
| Total | 36/150 | 72/150 | 72.0 |
| Begomovirus detected in whitefly DNA sample | 2/15 | 5/15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomathi Devi, R.; Jothika, C.; Sankari, A.; Lakshmi, S.; Malathi, V.G.; Renukadevi, P. Seed Transmission of Begomoviruses: A Potential Threat for Bitter Gourd Cultivation. Plants 2023, 12, 1396. https://doi.org/10.3390/plants12061396
Gomathi Devi R, Jothika C, Sankari A, Lakshmi S, Malathi VG, Renukadevi P. Seed Transmission of Begomoviruses: A Potential Threat for Bitter Gourd Cultivation. Plants. 2023; 12(6):1396. https://doi.org/10.3390/plants12061396
Chicago/Turabian StyleGomathi Devi, Ravisankar, Chinnaraj Jothika, Arjunan Sankari, Sethuraman Lakshmi, Varagur Ganesan Malathi, and Perumal Renukadevi. 2023. "Seed Transmission of Begomoviruses: A Potential Threat for Bitter Gourd Cultivation" Plants 12, no. 6: 1396. https://doi.org/10.3390/plants12061396