
Quick Decline and Stem Pitting Citrus tristeza virus Isolates Induce a Distinct Metabolomic Profile and Antioxidant Enzyme Activity in the Phloem Sap of Two Citrus Species
 ,
,  , and
, and
Abstract
:1. Introduction
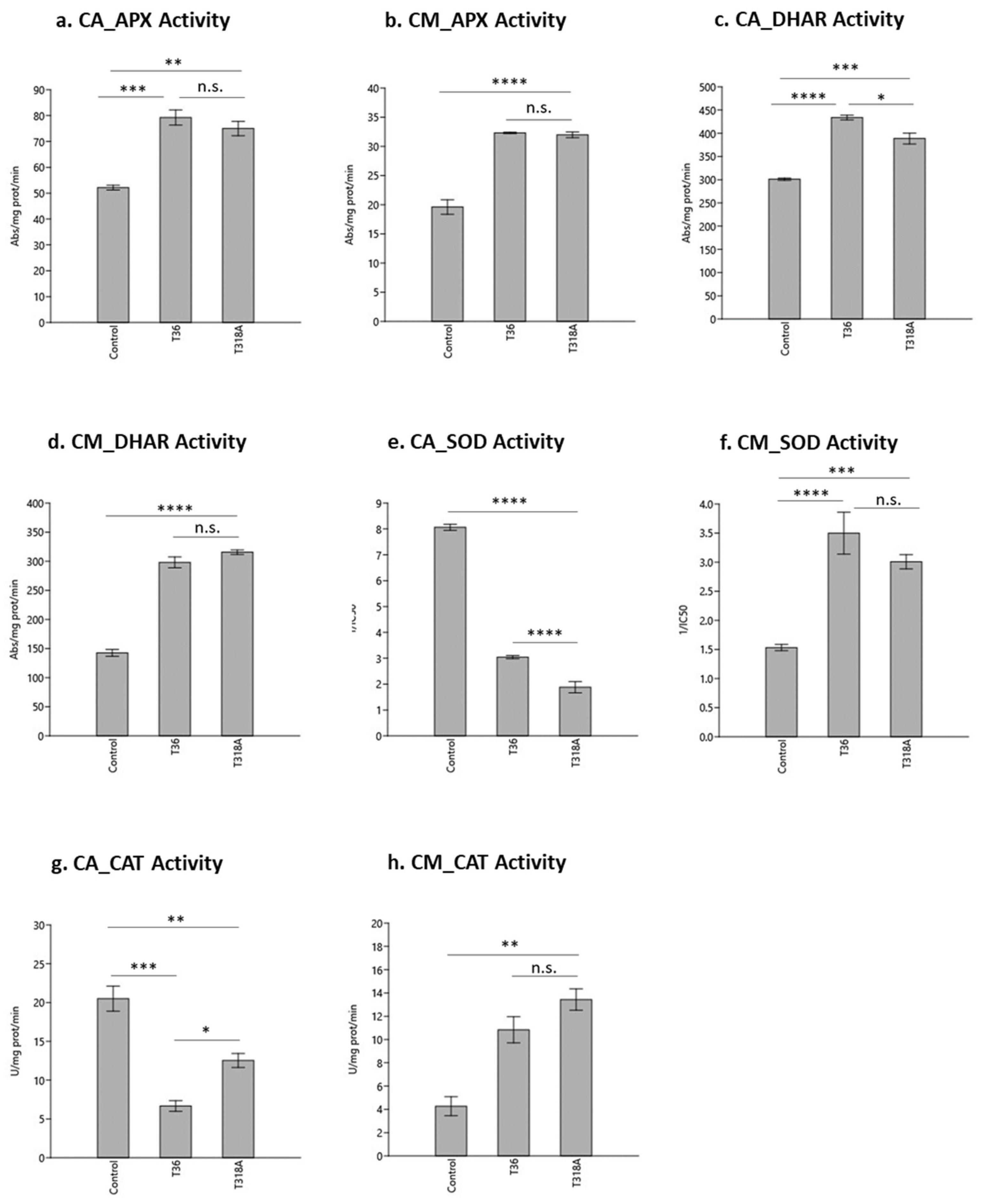
2. Results and Discussion
2.1. Enzymatic Antioxidant Profiling
2.2. Phloem Metabolites
3. Materials and Methods
3.1. Plant Material and Virus Isolates
3.2. Total RNA Extraction and CTV Detection by Reverse Transcription-PCR (RT-PCR)
3.3. Phloem Sap Collection
3.4. Enzymatic Antioxidant Assays
3.4.1. Enzymatic Antioxidant Assays of SOD, CAT, DHAR and APX Enzymes
3.4.2. Statistical Analysis
3.5. Analysis by UHPLC-HRMS and Data Processing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bar-Joseph, M.; Che, X.; Mawassi, M.; Gowda, S.; Satyanarayana, T.; Ayllon, M.A.; Albiach-Martí, M.R.; Garnsey, S.M.; Dawson, W.O. The Continuous Challenge of Citrus tristeza virus Molecular Research. In International Organization of Citrus Virologists Conference Proceedings (1957–2010); eScholarship: Oakland, CA, USA, 2002; Volume 15. [Google Scholar]
- Folimonova, S.Y. Citrus Tristeza Virus: A Large RNA Virus with Complex Biology Turned into a Valuable Tool for Crop Protection. PLoS Pathog. 2020, 16, e1008416. [Google Scholar] [CrossRef] [PubMed]
- Roistacher, C.N.; Moreno, P. The Worldwide Threat from Destructive Isolates of Citrus Tristeza Virus-A Review. International Organization of Citrus Virologists Conference Proceedings (1957–2010); eScholarship: Oakland, CA, USA, 1991; Volume 11. [Google Scholar]
- Folimonova, S.Y.; Folimonov, A.S.; Tatineni, S.; Dawson, W.O. Citrus Tristeza Virus: Survival at the Edge of the Movement Continuum. J. Virol. 2008, 82, 6546–6556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Muñoz, N.; Velázquez, K.; Vives, M.C.; Ruiz-Ruiz, S.; Pina, J.A.; Flores, R.; Moreno, P.; Guerri, J. The Resistance of Sour Orange to Citrus tristeza virus Is Mediated by Both the Salicylic Acid and RNA Silencing Defence Pathways. Mol. Plant Pathol. 2017, 18, 1253–1266. [Google Scholar] [CrossRef]
- Dória, M.S.; de Sousa, A.O.; Barbosa, C.D.J.; Costa, M.G.C.; Gesteira, A.D.S.; Souza, R.M.; Freitas, A.C.O.; Pirovani, C.P. Citrus tristeza virus (CTV) Causing Proteomic and Enzymatic Changes in Sweet Orange Variety “Westin”. PLoS ONE 2015, 10, e0130950. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.D.M.; Varanda, C.M.R.; Félix, M.R.F. Induced Resistance during the Interaction Pathogen x Plant and the Use of Resistance Inducers. Phytochem. Lett. 2016, 15, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Hernández, J.A.; Gullner, G.; Clemente-Moreno, M.J.; Künstler, A.; Juhász, C.; Díaz-Vivancos, P.; Király, L. Oxidative Stress and Antioxidative Responses in Plant–Virus Interactions. Physiol. Mol. Plant Pathol. 2016, 94, 134–148. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Hančević, K.; Radić, T.; Pasković, I.; Urlić, B. Biochemical and Physiological Responses to Long-Term Citrus tristeza virus Infection in Mexican Lime Plants. Plant Pathol. 2017, 67, 987–994. [Google Scholar] [CrossRef]
- Walz, C.; Juenger, M.; Schad, M.; Kehr, J. Evidence for the Presence and Activity of a Complete Antioxidant Defence System in Mature Sieve Tubes. Plant J. 2002, 31, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Racchi, M. Antioxidant Defenses in Plants with Attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef]
- Pérez-Clemente, R.M.; Montoliu, A.; Vives, V.; López-Climent, M.F.; Gómez-Cadenas, A. Photosynthetic and Antioxidant Responses of Mexican Lime (Citrus aurantifolia) Plants to Citrus tristeza virus Infection. Plant Pathol. 2015, 64, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Serra-Soriano, M.; Navarro, J.A.; Genoves, A.; Pallás, V. Comparative Proteomic Analysis of Melon Phloem Exudates in Response to Viral Infection. J. Proteomics. 2015, 124, 11–24. [Google Scholar] [CrossRef]
- Gaupels, F.; Durner, J.; Kogel, K. Production, Amplification and Systemic Propagation of Redox Messengers in Plants? The Phloem Can Do It All! New Phytol. 2017, 214, 554–560. [Google Scholar] [CrossRef] [Green Version]
- Ádám, A.; Nagy, Z.; Kátay, G.; Mergenthaler, E.; Viczián, O. Signals of Systemic Immunity in Plants: Progress and Open Questions. Int. J. Mol. Sci. 2018, 19, 1146. [Google Scholar] [CrossRef] [Green Version]
- Kappagantu, M.; Collum, T.D.; Dardick, C.; Culver, J.N. Viral Hacks of the Plant Vasculature: The Role of Phloem Alterations in Systemic Virus Infection. Annu Rev. Virol. 2020, 7, 351–370. [Google Scholar] [CrossRef]
- Ham, B.-K.; Lucas, W.J. Phloem-Mobile RNAs as Systemic Signaling Agents. Annu. Rev. Plant Biol. 2017, 68, 173–195. [Google Scholar] [CrossRef]
- Killiny, N. Metabolomic Comparative Analysis of the Phloem Sap of Curry Leaf Tree (Bergerac koenegii), Orange Jasmine (Murraya paniculata), and Valencia Sweet Orange (Citrus sinensis) Supports Their Differential Responses to Huanglongbing. Plant Signal Behav. 2016, 11, e1249080. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Zhang, Y.; Zhong, Y.; Yang, J.; Yan, S. Gene Expression Changes in Leaves of Citrus sinensis (L.) Osbeck Infected by Citrus Tristeza Virus. J. Hortic. Sci. Biotechnol. 2016, 91, 466–475. [Google Scholar] [CrossRef]
- Killiny, N. Metabolite Signature of the Phloem Sap of Fourteen Citrus Varieties with Different Degrees of Tolerance to Candidatus Liberibacter asiaticus. Physiol. Mol. Plant Pathol. 2017, 97, 20–29. [Google Scholar] [CrossRef]
- Killiny, N.; Nehela, Y. Metabolomic Response to Huanglongbing: Role of Carboxylic Compounds in Citrus sinensis Response to ‘Candidatus Liberibacter asiaticus’ and Its Vector, Diaphorina citri. Mol. Plant-Microbe Interact. 2017, 30, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Maksoud, S.; Abdel-Massih, R.M.; Rajha, H.N.; Louka, N.; Chemat, F.; Barba, F.J.; Debs, E. Citrus aurantium L. Active Constituents, Biological Effects and Extraction Methods. An Updated Review. Molecules 2021, 26, 5832. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Hijaz, F.; Harper, S.J.; Dawson, W.O. Effects of Citrus Tristeza closterovirus Infection on Phloem Sap and Released Volatile Organic Compounds in Citrus macrophylla. Physiol. Mol. Plant Pathol. 2017, 98, 25–36. [Google Scholar] [CrossRef]
- Dawson, W.O.; Garnsey, S.M.; Tatineni, S.; Folimonova, S.Y.; Harper, S.J.; Gowda, S. Citrus Tristeza Virus-Host Interactions. Front. Microbiol. 2013, 4, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Laino, P.; Russo, M.P.; Guardo, M.; Reforgiato-Recupero, G.; Valè, G.; Cattivelli, L.; Moliterni, V.M.C. Rootstock-Scion Interaction Affecting Citrus Response to CTV Infection: A Proteomic View. Physiol. Plant 2016, 156, 444–467. [Google Scholar] [CrossRef]
- Kuźnia, E.; Kopczewski, T.; Chojak-Koźniewska, J. Ascorbate-Glutathione Cycle and Biotic Stress Tolerance in Plants. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 201–231. [Google Scholar]
- Clarke, S.F.; Guy, P.L.; Burritt, D.J.; Jameson, P.E. Changes in the Activities of Antioxidant Enzymes in Response to Virus Infection and Hormone Treatment. Physiol. Plant 2002, 114, 157–164. [Google Scholar] [CrossRef]
- Zhuang, B.; Xu, B.; Liao, L. Change of Superoxide Dismutase, Peroxidase and Storage Protein in Soybean Leaves after Infection with Soybean Mosaic Virus. Acta Phytopathol. Sinica 1993, 23, 261–265. [Google Scholar]
- Hernández, J.A.; Talavera, J.M.; Martínez-Gómez, P.; Dicenta, F.; Sevilla, F. Response of Antioxidative Enzymes to Plum Pox Virus in Two Apricot Cultivars. Physiol. Plant 2001, 111, 313–321. [Google Scholar] [CrossRef]
- Inaba, J.; Kim, B.M.; Shimura, H.; Masuta, C. Virus-Induced Necrosis Is a Consequence of Direct Protein-Protein Interaction between a Viral RNA-Silencing Suppressor and a Host Catalase. Plant Physiol. 2011, 156, 2026–2036. [Google Scholar] [CrossRef] [Green Version]
- Rizhsky, L.; Hallak-Herr, E.; van Breusegem, F.; Rachmilevitch, S.; Barr, J.E.; Rodermel, S.; Inzé, D.; Mittler, R. Double Antisense Plants Lacking Ascorbate Peroxidase and Catalase Are Less Sensitive to Oxidative Stress than Single Antisense Plants Lacking Ascorbate Peroxidase or Catalase. Plant J. 2002, 32, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Sousa, R.H.V.; Carvalho, F.E.L.; Lima-Melo, Y.; Alencar, V.T.C.B.; Daloso, D.M.; Margis-Pinheiro, M.; Komatsu, S.; Silveira, J.A.G. Impairment of Peroxisomal APX and CAT Activities Increases Protection of Photosynthesis under Oxidative Stress. J. Exp. Bot. 2019, 70, 627–639. [Google Scholar] [CrossRef]
- Viant, M.R.; Kurland, I.J.; Jones, M.R.; Dunn, W.B. How Close Are We to Complete Annotation of Metabolomes? Curr. Opin. Chem. Biol. 2017, 36, 64–69. [Google Scholar] [CrossRef]
- Suntar, I.; Khan, H.; Patel, S.; Celano, R.; Rastrelli, L. An Overview on Citrus aurantium L.: Its Functions as Food Ingredient and Therapeutic Agent. Oxid. Med. Cell. Longev. 2018, 2018, 7864269. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.; Wang, K.; Chen, B.; Liu, M.; Yang, W.; Ning, J.; Cai, Y.; Wei, M. The Enhanced Solubility and Anti-Lipase Activity of Citrus Peel Polymethoxyflavonoids Extracts with Liposomal Encapsulation. LWT 2022, 161, 113395. [Google Scholar] [CrossRef]
- Wang, S.; Yang, C.; Tu, H.; Zhou, J.; Liu, X.; Cheng, Y.; Luo, J.; Deng, X.; Zhang, H.; Xu, J. Characterization and Metabolic Diversity of Flavonoids in Citrus Species. Sci. Rep. 2017, 7, 10549. [Google Scholar] [CrossRef]
- Shen, C.-Y.; Jiang, J.-G.; Huang, C.-L.; Zhu, W.; Zheng, C.-Y. Polyphenols from Blossoms of Citrus aurantium L. Var. amara Engl. Show Significant Anti-Complement and Anti-Inflammatory Effects. J. Agric. Food Chem. 2017, 65, 9061–9068. [Google Scholar] [CrossRef]
- Li, S.; Lo, C.-Y.; Ho, C.-T. Hydroxylated Polymethoxyflavones and Methylated Flavonoids in Sweet Orange (Citrus sinensis) Peel. J. Agric. Food Chem. 2006, 54, 4176–4185. [Google Scholar] [CrossRef]
- Gao, Z.; Gao, W.; Zeng, S.-L.; Li, P.; Liu, E.-H. Chemical Structures, Bioactivities and Molecular Mechanisms of Citrus Polymethoxyflavones. J. Funct. Foods 2018, 40, 498–509. [Google Scholar] [CrossRef]
- Ballistreri, G.; Fabroni, S.; Romeo, F.V.; Timpanaro, N.; Amenta, M.; Rapisarda, P. Anthocyanins and Other Polyphenols in Citrus Genus: Biosynthesis, Chemical Profile, and Biological Activity. In Polyphenols in Plants; Watson, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 191–215. [Google Scholar]
- Khalid, M.; Saeed-ur-Rahman; Bilal, M.; Huang, D. Role of Flavonoids in Plant Interactions with the Environment and against Human Pathogens—A Review. J. Integr. Agric. 2019, 18, 211–230. [Google Scholar] [CrossRef]
- Asai, T.; Matsukawa, T.; Kajiyama, S. Metabolomic Analysis of Primary Metabolites in Citrus Leaf during Defense Responses. J. Biosci. Bioeng. 2017, 123, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.Y.; Thapa, S.P.; Pang, Z.; Gurung, F.B.; Liebrand, T.W.H.; Stevens, D.M.; Ancona, V.; Wang, N.; Coaker, G. Citrus Vascular Proteomics Highlights the Role of Peroxidases and Serine Proteases during Huanglongbing Disease Progression. Mol. Cell. Proteom. 2020, 19, 1936–1952. [Google Scholar] [CrossRef] [PubMed]
- Song, G.C.; Choi, H.K.; Ryu, C.-M. The Folate Precursor Para-Aminobenzoic Acid Elicits Induced Resistance against Cucumber Mosaic Virus and Xanthomonas axonopodis. Ann. Bot. 2013, 111, 925–934. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H.; Sakihama, Y.; Lkehara, N. Flavonoid-Peroxidase Reaction as a Detoxification Mechanism of Plant Cells against H2O2. Plant Physiol. 1997, 11, 1405–1412. [Google Scholar] [CrossRef] [Green Version]
- Bolton, M.D. Primary Metabolism and Plant Defense—Fuel for the Fire. Mol. Plant-Microbe Interact. 2009, 22, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Xu, X.; Li, Y.; Wang, Y.; Li, M.; Wang, Y.; Ding, X.; Chu, Z. Rutin-Mediated Priming of Plant Resistance to Three Bacterial Pathogens Initiating the Early SA Signal Pathway. PLoS ONE 2016, 11, e0146910. [Google Scholar] [CrossRef]
- Almagro, L.; Gómez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreño, M.A. Class III Peroxidases in Plant Defence Reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, R.; Lim, G.-H.; de Lorenzo, L.; Yu, K.; Zhang, K.; Hunt, A.G.; Kachroo, A.; Kachroo, P. Pipecolic Acid Confers Systemic Immunity by Regulating Free Radicals. Sci. Adv. 2018, 4, eaar4509. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Kawano, T. Salicylic Acid-Induced Superoxide Generation Catalyzed by Plant Peroxidase in Hydrogen Peroxide-Independent Manner. Plant Signal Behav. 2015, 10, e1000145. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Luo, Q.; Yang, W.; Ahammed, G.J.; Ding, S.; Chen, X.; Wang, J.; Xia, X.; Shi, K. A Novel Role of Pipecolic Acid Biosynthetic Pathway in Drought Tolerance through the Antioxidant System in Tomato. Antioxidants 2021, 10, 1923. [Google Scholar] [CrossRef]
- Wang, S.; Alseekh, S.; Fernie, A.R.; Luo, J. The Structure and Function of Major Plant Metabolite Modifications. Mol. Plant 2019, 12, 899–919. [Google Scholar] [CrossRef]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising Natural Compounds against Viral Infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Kim, S.H.; Bahk, S.; Vuong, U.T.; Nguyen, N.T.; Do, H.L.; Kim, S.H.; Chung, W.S. Naringenin Induces Pathogen Resistance Against Pseudomonas syringae Through the Activation of NPR1 in Arabidopsis. Front. Plant Sci. 2021, 12, 672552. [Google Scholar] [CrossRef]
- Wang, L.; Chen, M.; Lam, P.Y.; Dini-Andreote, F.; Dai, L.; Wei, Z. Multifaceted Roles of Flavonoids Mediating Plant-Microbe Interactions. Microbiome 2022, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Olfa, T.; Gargouri, M.; Akrouti, A.; Brits, M.; Gargouri, M.; ben Ameur, R.; Pieters, L.; Foubert, K.; Magné, C.; Soussi, A.; et al. A Comparative Study of Phytochemical Investigation and Antioxidative Activities of Six Citrus Peel Species. Flavour Fragr. J. 2021, 36, 564–575. [Google Scholar] [CrossRef]
- Dawson, W.O.; Bar-Joseph, M.; Garnsey, S.M.; Moreno, P. Citrus tristeza virus: Making an Ally from an Enemy. Annu. Rev. Phytopathol. 2015, 53, 137–155. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, S.; Moreno, P.; Guerri, J.; Ambrós, S. The Complete Nucleotide Sequence of a Severe Stem Pitting Isolate of Citrus tristeza virus from Spain: Comparison with Isolates from Different Origins. Arch. Virol. 2006, 151, 387–398. [Google Scholar] [CrossRef]
- Afonso, A.M.; Guerra, R.; Cavaco, A.M.; Pinto, P.; Andrade, A.; Duarte, A.; Power, D.M.; Marques, N.T. Identification of Asymptomatic Plants Infected with Citrus tristeza virus from a Time Series of Leaf Spectral Characteristics. Comput. Electron. Agric. 2017, 141, 340–350. [Google Scholar] [CrossRef]
- Hijaz, F.; Killiny, N. Collection and Chemical Composition of Phloem Sap from Citrus sinensis L. Osbeck (Sweet Orange). PLoS ONE 2014, 9, e101830. [Google Scholar] [CrossRef] [Green Version]
- Killiny, N. Collection of the Phloem Sap, Pros and Cons. Plant Signal Behav. 2019, 14, 1618181. [Google Scholar] [CrossRef]
- Chandlee, J.M.; Scandalios, J.G. Analysis of Variants Affecting the Catalase Developmental Program in Maize Scutellum. Theor. Appl. Genet. 1984, 69, 71–77. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef] [PubMed]
- de Gara, L.; Paciolla, C.; de Tullio, M.C.; Motto, M.; Arrigoni, O. Ascorbate-Dependent Hydrogen Peroxide Detoxification and Ascorbate Regeneration during Germination of a Highly Productive Maize Hybrid: Evidence of an Improved Detoxification Mechanism against Reactive Oxygen Species. Physiol. Plant 2000, 109, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Hammerschmidt, R.; Nuckles, E.M.; Kuć, J. Association of Enhanced Peroxidase Activity with Induced Systemic Resistance of Cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]


{kind=link}
| N° | Name | Formula | m/z (+/−) | mzCloud Match | Ratio CA/CM |
|---|---|---|---|---|---|
| 1 | ♦ 5,3′-Dihydroxy-3,6,7,4′ tetramethoxyflavone | C19 H18 O8 | 373.0911 (−) | 95.1 | 994 |
| 2 | Eriodictyol | C15 H12 O6 | 289.0695 (+) | 98.6 | 858 |
| 3 | ♦ Naringin | C27 H32 O14 | 579.1726 (−) | 95.5 | 792 |
| 4 | Rhoifolin | C27 H30 O14 | 579.1727 (+) | 98.3 | 745 |
| 5 | 5,2′-Dihydroxy-6,7,8,6′ tetramethoxyflavone | C19 H18 O8 | 375.1062 (+) | 91.1 | 419 |
| 6 | 5,7-Dihydroxy-6,3′,4′-trimethoxyflavone | C18 H16 O7 | 345.0953 (+) | 95.0 | 204 |
| 7 | Luteolin 7-rutinoside | C27 H30 O15 | 593.1530 (−) | 91.5 | 152 |
| 8 | ♦ Quercetin | C15 H10 O7 | 303.0490 (+) | 95.1 | 147 |
| 9 | 5,6,7,8,3′,4′-Hexamethoxyflavone | C21 H22 O8 | 403.1368 (+) | 98.6 | 141 |
| 10 | Apigenin | C15 H10 O5 | 271.0593 (+) | 93.1 | 140 |
| 11 | Naringenin | C15 H12 O5 | 273.0746 (+) | 98.4 | 135 |
| 12 | 3-Methylkaempferol | C16 H12 O6 | 301.0700 (+) | 99.3 | 132 |
| 15 | Hesperetin | C16 H14 O6 | 303.0849 (+) | 99.9 | 104 |
| 13 | Neodiosmin | C28 H32 O15 | 609.1803 (+) | 97.9 | 90 |
| 14 | ♦ Isoferulic acid | C10 H10 O4 | 195.0646 (+) | 99.9 | 80 |
| 16 | 3-O-Feruloylquinic acid | C17 H20 O9 | 367.1019 (−) | 95.5 | 84 |
| 17 | Prunin | C21 H22 O10 | 435.1268 (+) | 91.3 | 56 |
| 18 | Isorhamnetin | C16 H12 O7 | 317.0647 (+) | 97.4 | 51 |
| 19 | ♦ Neohesperidin | C28 H34 O15 | 609.1805 (−) | 97.6 | 42 |
| 20 | Apigenin 6,8-di-C-glucoside | C27 H30 O15 | 593.1532 (−) | 93.0 | 40 |
| 21 | Naringenin 4′-glucoside 7-rutinoside | C33 H42 O19 | 741.2214 (−) | 100.0 | 32 |
| 22 | ♦ Biorobin | C27 H30 O15 | 593.1525 (−) | 89.4 | 32 |
| 23 | Vitexin | C21 H20 O10 | 423.1115 (+) | 95.9 | 16 |
| 24 | Methyl caffeate | C10 H10 O4 | 195.0646 (+) | 98.3 | 9 |
| 25 | 4-Hydroxybenzaldehyde | C7 H6 O2 | 123.0436 (+) | 99.4 | 0.50 |
| 26 | Quercetin-3β-D-glucoside | C21 H20 O12 | 465.1017 (+) | 98.2 | 0.34 |
| 27 | Methyl cinnamate | C10 H10 O2 | 163.0755 (+) | 96.7 | 0.25 |
| 28 | 4-Coumaric acid | C9 H8 O3 | 165.0540 (+) | 99.8 | 0.18 |
| 29 | 3-[3-(beta-D-Glucopyranosyloxy)-2-methoxyphenyl] propanoic acid | C16 H22 O9 | 357.1173 (−) | 96.9 | 0.13 |
| 30 | Cnidioside A | C17 H20 O9 | 367.1019 (−) | 99.6 | 0.09 |
| 31 | Dihydrocaffeic acid | C9 H10 O4 | 181.0502 (-) | 90.7 | 0.08 |
| 32 | Asparagin | C4 H8 N2 O3 | 133.0603 (+) | 96.1 | 0.07 |
| 33 | Scopoletin | C10 H8 O4 | 193.0487 (+) | 92.6 | 0.06 |
| 34 | 2,4-Dimethylbenzaldehyde | C9 H10 O | 135.0799 (+) | 87.2 | 0.06 |
| 35 | 7-Hydroxycoumarine | C9 H6 O3 | 163.0385 (+) | 98.4 | 0.05 |
| 36 | ♦ 4-Aminobenzoic acid | C7 H7 N O2 | 136.0402 (−) | 97.3 | 0.05 |
| 37 | ♦ Rutin | C27 H30 O16 | 609.1442 (−) | 98.8 | 0.02 |
| 38 | ♦ Ferulic acid | C10 H10 O4 | 193.0509 (−) | 99.1 | 0.01 |
| 39 | L-(-)-Methionine | C5 H11 N O2 S | 150.0671 (+) | 99.8 | 0.007 |
| 40 | Eriocitrin | C27 H32 O15 | 595.1650 (−) | 99.5 | 0.006 |
| 41 | ♦ 2-Hydroxycinnamic acid | C9 H8 O3 | 163.0398 (−) | 97.1 | 0.003 |
| N° | Name | Formula | m/z | mzCloud Match | Ratio INF_CA T36/N-INF_CA |
| 1 | 3-[4-(beta-D-Glucopyranosyloxy)-6-methoxy-1-benzofuran-5-yl]propanoic acid | C17 H20 O9 | 367.1022 (−) | 99.5 | 43 |
| 2 | ♦ Biorobin | C27 H30 O15 | 583.1520 (−) | 89.4 | 0.147 |
| N° | Name | Formula | m/z | mzCloud Match | Ratio INF_CM T36/N-INF_CM |
| 1 | 2-[[6-hydroxy-4-(4-hydroxy-3,5-dimethoxyphenyl)-3-(hydroxymethyl)-5,7-dimethoxy-1,2,3,4-tetrahydronaphthalen-2-yl]methoxy]-6-(hydroxymethyl)oxane-3,4,5-triol | C28 H38 O13 | 581.2223 (−) | 98.1 | 200 |
| 2 | 5,6′-Dihydroxy-6,7,8,2′-tetramethoxyflavone | C19 H18 O8 | 375.1061 (+) | 91.1 | 16 |
| 3 | Isovanillic acid | C8 H8 O4 | 169.0489 (+) | 99.4 | 12 |
| 4 | cis-Aconitic acid | C6 H6 O6 | 173.0086 (−) | 89.2 | 9 |
| 5 | Nobiletin | C21 H22 O8 | 403.1378 (+) | 95.6 | 6 |
| 6 | 3,4-Dihydroxybenzaldehyde | C7 H6 O3 | 139.0387 (+) | 97.4 | 5 |
| 7 | ♦ Quercetin | C15 H10 O7 | 303.0486 (+) | 95.1 | 5 |
| 8 | ♦ Naringin | C27 H32 O14 | 579.1729 (−) | 95.5 | 5 |
| 9 | ♦ 4-Aminobenzoic acid | C7 H7 N O2 | 136.0402 (−) | 97.2 | 0.2 |
| 10 | ♦ Isoferulic acid | C10 H10 O4 | 195.0648 (+) | 94.7 | 0.07 |
| 11 | Glucose-1,6-bisphosphate | C6 H14 O12 P2 | 338.9875 (−) | 99.1 | 0.03 |
| 12 | Guanosine | C10 H13 N5 O5 | 338.9875 (−) | 99.9 | 0.003 |
| N° | Name | Formula | m/z | mzCloud Match | Ratio INF_CA_T318A/N-INF_CA |
| 1 | 5,3′-Dihydroxy-3,6,7,4′-tetramethoxyflavone | C19 H18 O8 | 373.0994 (−) | 94.4 | 403 |
| 2 | Fumaric acid | C4 H4 O4 | 115.0035 (−) | 99.5 | 55 |
| 3 | cis-Aconitic acid | C6 H6 O6 | 173.0085 (−) | 89.2 | 17 |
| 4 | Isovanillic acid | C8 H8 O4 | 167.0346 (−) | 96.6 | 0.4 |
| 5 | 3-[2-(β-D-Glucopyranosyloxy)-4-methoxyphenyl]propanoic acid | C16 H22 O9 | 357.1173 (−) | 99.6 | 0.4 |
| 6 | ♦ 2-Hydroxycinnamic acid | C9 H8 O3 | 163.0398 (−) | 91.5 | 0.18 |
| 7 | ♦ Neohesperidin | C28 H34 O15 | 609.1805 (−) | 97.6 | 0.17 |
| 8 | Hesperidin | C28 H34 O15 | 609.1825 (−) | 98.5 | 0.002 |
| N° | Name | Formula | m/z | mzCloud Match | Ratio INF_CM_T318A/N-INF_CM |
| 1 | 3-[2-(β-D-Glucopyranosyloxy)-4-methoxyphenyl]propanoic acid | C16 H22 O9 | 357.1166 (−) | 99.1 | 1977 |
| 2 | Pipecolic acid | C6 H11 N O2 | 130.0862 (+) | 98.1 | 109 |
| 3 | Salicylic acid | C7 H6 O3 | 137.0233 (−) | 99.8 | 22 |
| 4 | Proline | C5 H9 N O2 | 116.0704 (+) | 100.0 | 21 |
| 5 | 3,4-Dihydroxyphenylpropionic acid | C9 H10 O4 | 181.0509 (−) | 90.7 | 0.023 |
| 6 | Hesperidin | C28 H34 O15 | 609.1824 (−) | 98.5 | 0.019 |
| 7 | ♦ Ferulic acid | C10 H10 O4 | 193.0513 (−) | 98.9 | 0.013 |
| 8 | ♦ 4-Aminobenzoic acid | C7 H7 N O2 | 136.0402 (−) | 97.3 | 0.011 |
| 9 | Isovanillic acid | C8 H8 O4 | 167.0345 (−) | 96.1 | 0.008 |
| 10 | Guanosine | C10 H13 N5 O5 | 338.9874 (−) | 99.9 | 0.005 |
| 11 | ♦ Rutin | C27 H30 O16 | 609.1443 (−) | 98.8 | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dandlen, S.A.; Da Silva, J.P.; Miguel, M.G.; Duarte, A.; Power, D.M.; Marques, N.T. Quick Decline and Stem Pitting Citrus tristeza virus Isolates Induce a Distinct Metabolomic Profile and Antioxidant Enzyme Activity in the Phloem Sap of Two Citrus Species. Plants 2023, 12, 1394. https://doi.org/10.3390/plants12061394
Dandlen SA, Da Silva JP, Miguel MG, Duarte A, Power DM, Marques NT. Quick Decline and Stem Pitting Citrus tristeza virus Isolates Induce a Distinct Metabolomic Profile and Antioxidant Enzyme Activity in the Phloem Sap of Two Citrus Species. Plants. 2023; 12(6):1394. https://doi.org/10.3390/plants12061394
Chicago/Turabian StyleDandlen, Susana A., José P. Da Silva, Maria Graça Miguel, Amílcar Duarte, Deborah M. Power, and Natália Tomás Marques. 2023. "Quick Decline and Stem Pitting Citrus tristeza virus Isolates Induce a Distinct Metabolomic Profile and Antioxidant Enzyme Activity in the Phloem Sap of Two Citrus Species" Plants 12, no. 6: 1394. https://doi.org/10.3390/plants12061394