
Transcriptomic Analysis Reveals Panicle Heterosis in an Elite Hybrid Rice ZZY10 and Its Parental Lines
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Characterization of ZZY10 and Its Two Parental Lines
2.2. Mapping Short Reads to the Rice Genome
2.3. Differentially Expressed Genes (DEGs) Identified by RNA-Seq
2.4. Gene Ontology (GO) Analysis
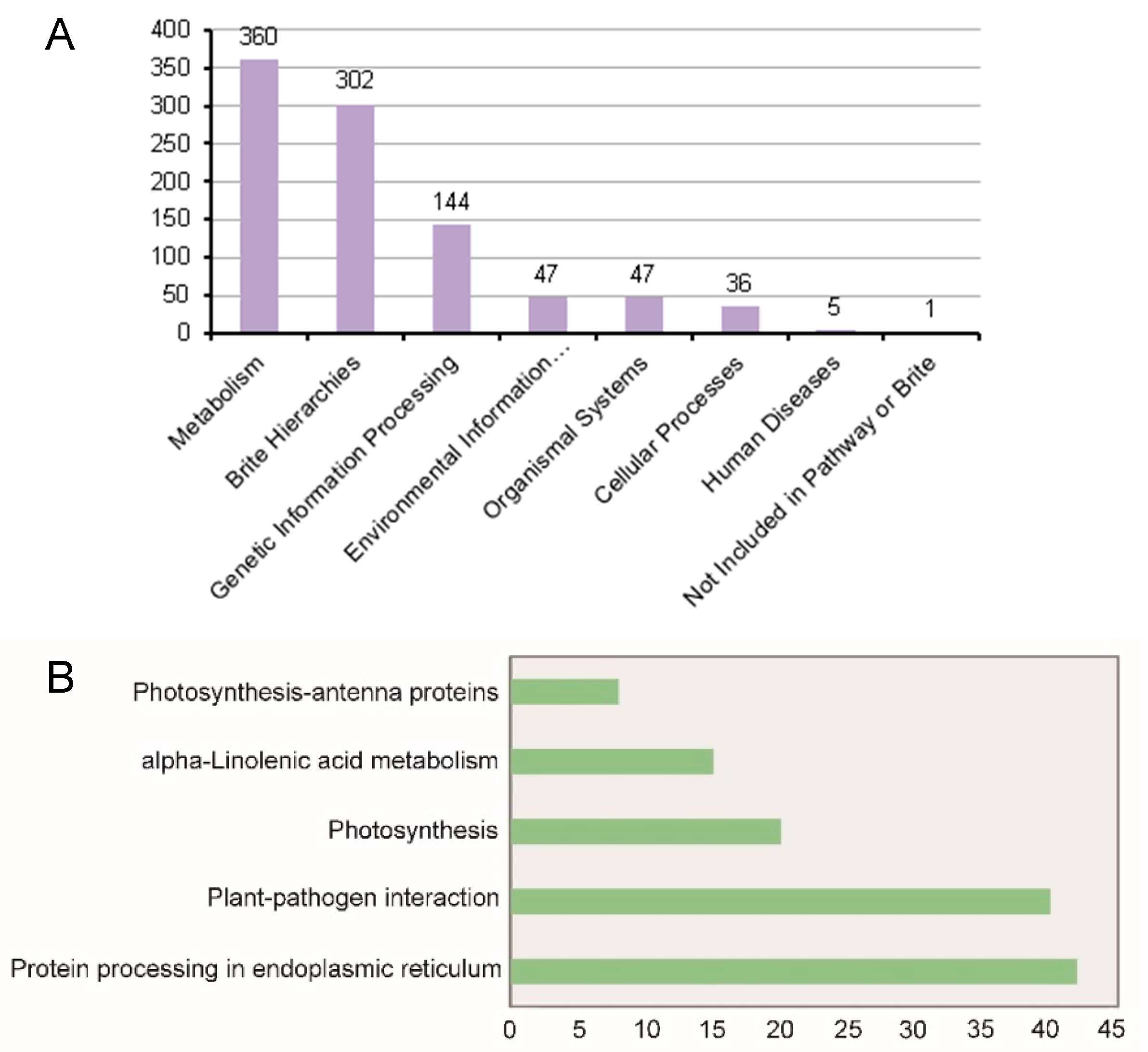
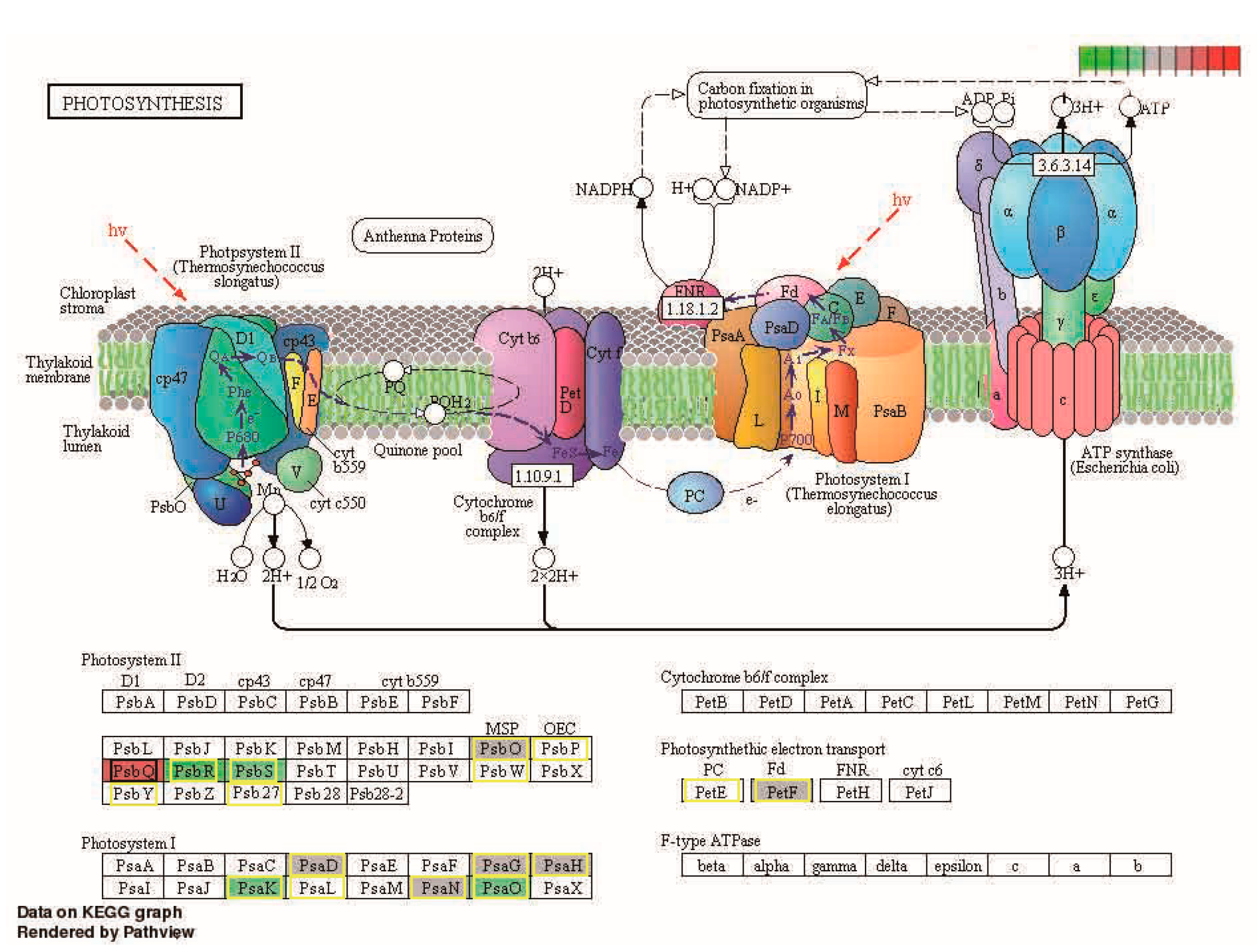
2.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR)
3. Discussion
3.1. Comparative Analysis of the Annotated DGHP
3.2. Photosynthesis Regulation in Heterosis
3.3. Implications for the Mechanism of Heterosis
4. Materials and Methods
4.1. Plants and Growing Conditions
4.2. Agronomic Trait Evaluation
4.3. Transcriptome Analysis
4.4. qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, S.H.; Zhuang, J.Y.; Fan, Y.Y.; Du, J.H.; Cao, L.Y. Progress in research and development on hybrid rice: A super-domesticate in China. Ann. Bot. 2007, 100, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.H.; Yuan, L.P. Hybrid rice achievements, development and prospect in China. J. Integr. Agric. 2015, 14, 197–205. [Google Scholar] [CrossRef]
- Khush, G.S. Breaking the yield frontier of rice. Geojournal 1995, 35, 329–332. [Google Scholar] [CrossRef]
- Lawson, T.; Flexas, J. Fuelling life: Recent advances in photosynthesis research. Plant J. 2020, 101, 753–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, S.P.; Marshall-Colon, A.; Zhu, X.G. Meeting the Global Food Demand of the Future by Engineering Crop Photosynthesis and Yield Potential. Cell 2015, 161, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Aschan, G.; Pfanz, H. Non-foliar photosynthesis and its contribution to the overall carbon balance of plants. Ann. Des Sci. 2003, 25, 3–23. [Google Scholar] [CrossRef] [Green Version]
- Simkin, A.J.; Faralli, M.; Ramamoorthy, S.; Lawson, T. Photosynthesis in non-foliar tissues: Implications for yield. Plant J. 2020, 101, 1001–1015. [Google Scholar] [CrossRef]
- Nelson, N.; Ben-Shem, A. The complex architecture of oxygenic photosynthesis. Nat. Rev. Mol. Cell Biol. 2004, 5, 971–982. [Google Scholar] [CrossRef]
- Wu, G.X.; Ma, L.; Yuan, C.; Dai, J.H.; Luo, L.; Poudyal, R.S.; Sayre, R.T.; Lee, C.H. Formation of light-harvesting complex II aggregates from LHCII–PSI–LHCI complexes in rice plants under high light. J. Exp. Bot. 2021, 72, 4938–4948. [Google Scholar] [CrossRef]
- Gao, P.; Xia, H.Q.; Li, Q.; Li, Z.Z.; Zhai, C.; Weng, L.; Mi, H.L.; Yan, S.; Datla, R.; Wang, H.; et al. PALE-GREEN LEAF 1, a rice cpSRP54 protein, is essential for the assembly of the PSI-LHCI supercomplex. Plant Direct 2022, 6, e451. [Google Scholar] [CrossRef]
- Reddy, M.R.; Mangrauthia, S.K.; Reddy, S.V.; Manimaran, P.; Yugandhar, P.; Naresh Babu, P.; Vishnukiran, T.; Subrahmanyam, D.; Sundaram, R.M.; Balachandran, S.M. PAP90, a novel rice protein plays a critical role in regulation of D1 protein stability of PSII. J. Adv. Res. 2021, 30, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Maydup, M.L.; Antonietta, M.; Guiamet, J.J.; Graciano, C.; Lopez, J.R.; Tambussi, E.A. The contribution of ear photosynthesis to grain filling in bread wheat (Triticum aestivum L.). Field Crops Res. 2010, 119, 48–58. [Google Scholar] [CrossRef]
- Sanchez-Bragado, R.; Molero, G.; Reynolds, M.P.; Araus, J.L. Photosynthetic contribution of the ear to grain filling in wheat: A comparison of different methodologies for evaluation. J. Exp. Bot. 2016, 67, 2787–2798. [Google Scholar] [CrossRef]
- Maydup, M.L.; Antonietta, M.; Graciano, C.; Guiamet, J.J.; Tambussi, E.A. The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: Responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field Crops Res. 2014, 167, 102–111. [Google Scholar] [CrossRef]
- Coolen, S.; Proietti, S.; Hickman, R.; Davila Olivas, N.H.; Huang, P.P.; Van Verk, M.C.; Van Pelt, J.A.; Wittenberg, A.H.; De Vos, M.; Prins, M.; et al. Transcriptome dynamics of Arabidopsis during sequential biotic and abiotic stresses. Plant J. 2016, 86, 249–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digel, B.; Pankin, A.; Korff, M.V. Global Transcriptome Profiling of Developing Leaf and Shoot Apices Reveals Distinct Genetic and Environmental Control of Floral Transition and Inflorescence Development in Barley. Plant Cell 2015, 27, 2318–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.T.; Zarka, D.G.; Van Buskirk, H.A.; Fowler, S.G.; Thomashow, M.F. Roles of the CBF2 and ZAT12 transcription factors in configuring the low temperature transcriptome of Arabidopsis. Plant J. 2005, 41, 195–211. [Google Scholar] [CrossRef]
- Ding, H.P.; Cheng, Q.; Luo, X.R.; Li, L.J.; Chen, Z.; Liu, H.J.; Gao, J.; Lin, H.J.; Shen, Y.O.; Zhao, M.J.; et al. Heterosis in Early Maize Ear Inflorescence Development: A Genome-Wide Transcription Analysis for Two Maize Inbred Lines and Their Hybrid. Int. J. Mol. Sci. 2014, 15, 13892–13915. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Liu, D.C.; Wu, J.; Zhao, X.B.; Hao, M.; Geng, S.F.; Yan, J.; Jiang, X.X.; Zhang, L.Q.; Wu, J.Y.; et al. mRNA and Small RNA Transcriptomes Reveal Insights into Dynamic Homoeolog Regulation of Allopolyploid Heterosis in Nascent Hexaploid Wheat. Plant Cell 2014, 26, 1878–1900. [Google Scholar] [CrossRef] [Green Version]
- Shankar, R.; Bhattacharjee, A.; Jain, M. Transcriptome analysis in different rice cultivars provides novel insights into desiccation and salinity stress responses. Sci. Rep. 2016, 6, 23719. [Google Scholar] [CrossRef] [Green Version]
- Song, G.Y.; Guo, Z.B.; Liu, Z.W.; Qu, X.F.; Jiang, D.M.; Wang, W.; Zhu, Y.G.; Yang, D.C. The phenotypic predisposition of the parent in F1hybrid is correlated with transcriptome preference of the positive general combining ability parent. BMC Genom. 2014, 15, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson-Wagner, R.A.; Jia, Y.; DeCook, R.; Borsuk, L.A.; Nettleton, D.; Schnable, P.S. All possible modes of gene action are observed in a global comparison of gene expression in a maize F-1 hybrid and its inbred parents. Proc. Natl. Acad. Sci. USA 2006, 103, 6805–6810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, G.; Tao, Y.; Liu, G.Z.; Chen, C.; Luo, R.Y.; Xia, H.; Gan, Q.; Zeng, H.P.; Lu, Z.K.; Han, Y.N.; et al. A transcriptomic analysis of superhybrid rice LYP9 and its parents. Proc. Natl. Acad. Sci. USA 2009, 106, 7695–7701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.K.; Luo, L.J.; Mei, H.W.; Wang, D.L.; Shu, Q.Y.; Tabien, R.; Zhong, D.B.; Ying, C.S.; Stansel, J.W.; Khush, G.S.; et al. Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice. I. Biomass and grain yield. Genetics 2001, 158, 1737–1753. [Google Scholar] [CrossRef] [PubMed]
- Shull, G.H. The Composition of a Field of Maize. J. Hered. 1908, 4, 296–301. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Xin, C. Gene location, chromosomal marker and inheritance of coat colour in pigs. Acta Vet. Zootech. Sin. 1997, 28, 97–102. [Google Scholar]
- Pickett, A.A. Hybrid wheat: Results and problems. J. Agric. Sci. 1993, 121, 1–259. [Google Scholar] [CrossRef]
- Ge, X.M.; Chen, W.H.; Song, S.H.; Wang, W.W.; Hu, S.N.; Yu, J. Transcriptomic profiling of mature embryo from an elite super-hybrid rice LYP9 and its parental lines. BMC Plant Biol. 2008, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- He, G.M.; Zhu, X.P.; Elling, A.A.; Chen, L.B.; Wang, X.F.; Guo, L.; Liang, M.Z.; He, H.; Zhang, H.Y.; Chen, F.F.; et al. Global Epigenetic and Transcriptional Trends among Two Rice Subspecies and Their Reciprocal Hybrids. Plant Cell 2010, 22, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Song, G.S.; Zhai, H.L.; Peng, Y.G.; Zhang, L.; Wei, G.; Chen, X.Y.; Xiao, Y.G.; Wang, L.L.; Chen, Y.J.; Wu, B.; et al. Comparative Transcriptional Profiling and Preliminary Study on Heterosis Mechanism of Super-Hybrid Rice. Mol. Plant 2010, 3, 1012–1025. [Google Scholar] [CrossRef]
- Zhang, H.Y.; He, H.; Chen, L.B.; Li, L.; Liang, M.Z.; Wang, X.F.; Liu, X.G.; He, G.M.; Chen, R.S.; Ma, L.G.; et al. A genome-wide transcription analysis reveals a close correlation of promoter INDEL polymorphism and heterotic gene expression in rice hybrids. Mol. Plant 2008, 1, 720–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ifuku, K. The PsbP and PsbQ family proteins in the photosynthetic machinery of chloroplasts. Plant Physiol. Biochem. 2014, 81, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ifuku, K.; Ishihara, S.; Shimamoto, R.; Ido, K.; Fumihiko, S. Structure, function, and evolution of the PsbP protein family in higher plants. Photosynth. Res. 2008, 98, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Pagliano, C.; Saracco, G.; Barber, J. Structural, functional and auxiliary proteins of photosystem II. Photosynth. Res. 2013, 116, 167–188. [Google Scholar] [CrossRef]
- Shen, J.R. The Structure of Photosystem II and the Mechanism of Water Oxidation in Photosynthesis. Annu. Rev. Plant Biol. 2015, 66, 23–48. [Google Scholar] [CrossRef] [Green Version]
- Bruce, A.B. The mendelian theory of heredity and the augmentation of vigor. Science 1910, 32, 627–628. [Google Scholar] [CrossRef]
- Yu, S.B.; Li, J.X.; Xu, C.G.; Tan, Y.F.; Gao, Y.J.; Li, X.H.; Zhang, Q.; Maroof, M.A.S. Importance of epistasis as the genetic basis of heterosis in an elite rice hybrid. Proc. Natl. Acad. Sci. USA 1997, 94, 9226–9231. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Liu, X. The Research on Autotetraploid Rice; Guangzhou South China University of Technology Press: Guangzhou, China, 2014; ISBN 9787562341208. [Google Scholar]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ZZB | Z7-10 | ZZY10 | HPH (%) | MPH (%) | |
|---|---|---|---|---|---|
| PL (cm) | 26.95 ± 0.82 | 29.83 ± 0.94 | 31.45 ± 1.61 | −0.77 | 10.79 ** |
| 1000-GW (g) | 1.54 ± 0.03 | 1.69 ± 0.05 | 1.94 ± 0.04 | 15.03 ** | 20.34 ** |
| PBN | 14.89 ± 0.93 | 13.87 ± 0.78 | 14.36 ± 1.01 | −2.24 | 1.16 |
| SBN | 53.22 ± 5.78 | 66.56 ± 5.55 | 59.22 ± 8.53 | −11.02 * | −1.11 |
| TN | 25 ± 3.16 | 11 ± 2.83 | 13 ± 1.79 | −47.20 ** | −26.67 ** |
| PH (cm) | 123.36 ± 1.55 | 123.02 ± 3.43 | 146.07 ± 1.15 | 16.81 ** | 18.66 ** |
| GL (cm) | 9.62 ± 0.27 | 9.24 ± 0.20 | 9.55 ± 0.09 | −0.80 | 0.80 |
| GW (cm) | 2.32 ± 0.06 | 2.62 ± 0.09 | 2.42 ± 0.07 | −7.49 ** | −1.88 |
| Stage | ZZB (cm) | Z7-10 (cm) | ZZY10 (cm) | HPH (%) | MPH (%) |
|---|---|---|---|---|---|
| meiosis stage | 11.05 ± 0.46 | 12.03 ± 0.41 | 13.84 ± 0.58 | 15.07 | 19.98 |
| uninucleate stage | 13.65 ± 0.61 | 14.94 ± 0.38 | 15.70 ± 0.48 | 5.11 | 9.85 |
| bicellular stage | 20.29 ± 0.86 | 21.08 ± 0.51 | 24.03 ± 0.69 | 14.03 | 16.19 |
| tricellular stage | 23.94 ± 0.67 | 26.58 ± 0.50 | 28.33 ± 0.72 | 6.56 | 12.13 |
| Heading stage | 26.95 ± 0.82 | 29.83 ± 0.94 | 31.45 ± 1.61 | −0.77 | 10.79 ** |
| Ripening stage | 29.84 ± 1.50 | 29.76 ± 0.97 | 30.23 ± 1.14 | −3.05 * | 1.44 |
| Genetic Class | Subclass | Expression Pattern | p-Value (Total Number of DEGs) | |
|---|---|---|---|---|
| 0.05 (9000) | ||||
| Number | % | |||
| Up-regulated | Over-dominance | ZZY10 > Z7 > ZZB | 1261 | 14.1 |
| ZZY10 > ZZB > Z7 | 1329 | 14.8 | ||
| ZZY10 > ZZB = Z7 | 0 | 0 | ||
| High-parent dominance | ZZY10 = ZZB > Z7 | 365 | 4.1 | |
| ZZY10 = Z7 > ZZB | 236 | 2.6 | ||
| Additivity | Additivity | ZZB < ZZY10 < Z7 | 2587 | 28.7 |
| Z7 < ZZY10 < ZZB | 910 | 10.1 | ||
| Down-regulated | Low-parent dominance | ZZY10 = ZZB < Z7 | 58 | 0.6 |
| ZZY10 = Z7 < ZZB | 139 | 1.5 | ||
| Under-dominance | ZZY10 < ZZB < Z7 | 731 | 8.1 | |
| ZZY10 < Z7 < ZZB | 1384 | 15.4 | ||
| ZZY10 < ZZB = Z7 | 0 | 0 | ||
| Category | GO_Term | GO_Term_Annotation | p Value |
|---|---|---|---|
| BP | GO:0015979 | photosynthesis | 2.98 × 10−6 |
| BP | GO:0015074 | DNA integration | 5.93 × 10−5 |
| BP | GO:0042545 | cell wall modification | 0.000219 |
| CC | GO:0009579 | thylakoid | 6.08 × 10−6 |
| CC | GO:0044436 | thylakoid part | 6.08 × 10−6 |
| CC | GO:0009521 | photosystem | 1.31 × 10−5 |
| CC | GO:0034357 | photosynthetic membrane | 2.04 × 10−5 |
| CC | GO:0009522 | photosystem I | 0.000582 |
| CC | GO:0009523 | photosystem II | 0.000645 |
| CC | GO:0005618 | cell wall | 0.001305 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Z.; Wu, Y.; Zhang, P.; Hu, G.; Fu, D.; Yu, G.; Tong, H. Transcriptomic Analysis Reveals Panicle Heterosis in an Elite Hybrid Rice ZZY10 and Its Parental Lines. Plants 2023, 12, 1309. https://doi.org/10.3390/plants12061309
Zhong Z, Wu Y, Zhang P, Hu G, Fu D, Yu G, Tong H. Transcriptomic Analysis Reveals Panicle Heterosis in an Elite Hybrid Rice ZZY10 and Its Parental Lines. Plants. 2023; 12(6):1309. https://doi.org/10.3390/plants12061309
Chicago/Turabian StyleZhong, Zhengzheng, Yawen Wu, Peng Zhang, Guocheng Hu, Dong Fu, Guoping Yu, and Hanhua Tong. 2023. "Transcriptomic Analysis Reveals Panicle Heterosis in an Elite Hybrid Rice ZZY10 and Its Parental Lines" Plants 12, no. 6: 1309. https://doi.org/10.3390/plants12061309