
Evaluation of Amino Acid Profiles of Rice Genotypes under Different Salt Stress Conditions
 , ,
, , 
Abstract
:1. Introduction
2. Results
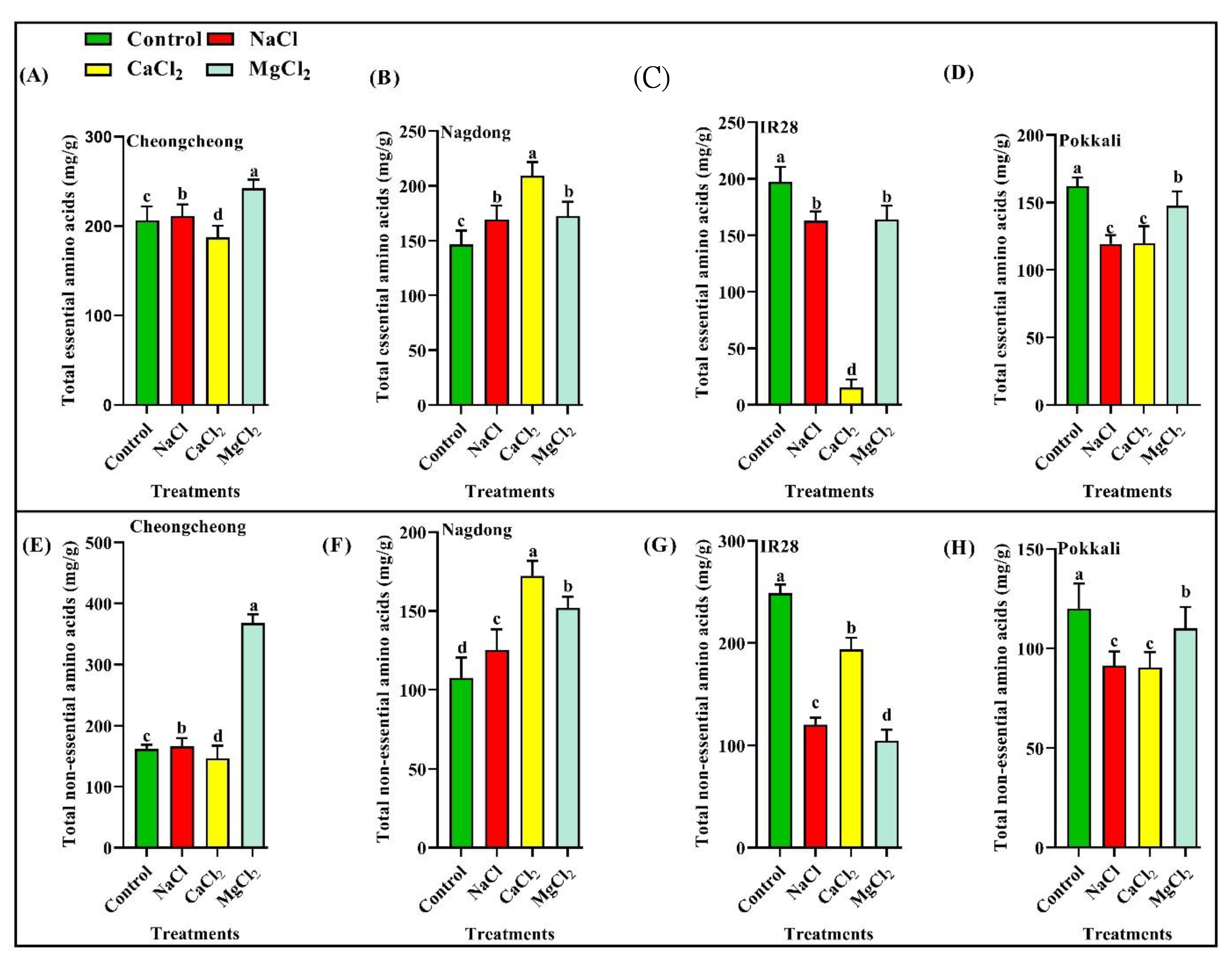
2.1. Effect of Different Salt Types on Essential Amino Acid Profiles
2.2. Effect of Different Salts on Non-Essential Amino Acids
2.3. Effect of Different Salt Types on Total Essential and Non-Essential Amino Acid Profiles
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Preparation and Application of Salt Solutions
3.3. Analysis of Amino Acid Profiles
3.4. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ahanger, M.A.; Aziz, U.; Alsahli, A.A.; Alyemeni, M.N.; Ahmad, P. Influence of exogenous salicylic acid and nitric oxide on growth, photosynthesis, and ascorbate-glutathione cycle in salt stressed Vigna angularis. Biomolecules 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahanger, M.A.; Mir, R.A.; Alyemeni, M.N.; Ahmad, P. Combined effects of brassinosteroid and kinetin mitigates salinity stress in tomato through the modulation of antioxidant and osmolyte metabolism. Plant Physiol. Biochem. 2020, 147, 31–42. [Google Scholar] [CrossRef]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T.J.B.l. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Amir, R.J.A.a. Current understanding of the factors regulating methionine content in vegetative tissues of higher plants. Amino Acids 2010, 39, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.P.s. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Ashraf, M.J.B.P. Organic substances responsible for salt tolerance inEruca sativa. Biol. Plant. 1994, 36, 255–259. [Google Scholar] [CrossRef]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Hülsenfrüchte. Lehrbuch der Lebensmittelchemie; Springer: Berlin/Heidelberg, Germany, 2001; pp. 731–756. [Google Scholar]
- Chen, Y.-C.; Holmes, E.C.; Rajniak, J.; Kim, J.-G.; Tang, S.; Fischer, C.R.; Mudgett, M.B.; Sattely, E.S.J.P.o.t.N.A.o.S. N-hydroxy-pipecolic acid is a mobile metabolite that induces systemic disease resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E4920–E4929. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Huang, W.; Zhang, F.; Luo, X.; Hu, B.; Xie, J. Metabolomic profiling of dongxiang wild rice under salinity demonstrates the significant role of amino acids in rice salt stress. Front. Plant Sci. 2021, 12, 729004. [Google Scholar] [CrossRef]
- Da Silva, C.J.; Fontes, E.P.B.; Modolo, L.V.J. Salinity-induced accumulation of endogenous H2S and NO is associated with modulation of the antioxidant and redox defense systems in Nicotiana tabacum L. cv. Havana. Plant Sci. 2017, 256, 148–159. [Google Scholar] [CrossRef]
- Farooq, M.; Asif, S.; Jang, Y.-H.; Park, J.-R.; Zhao, D.-D.; Kim, E.-G.; Kim, K.-M.J.F.i.P.S. Effect of Different Salts on Nutrients Uptake, Gene Expression, Antioxidant, and Growth Pattern of Selected Rice Genotypes. Front. Plant Sci. 2022, 13, 173–178. [Google Scholar] [CrossRef]
- Fougere, F.; Le Rudulier, D.; Streeter, J.G. Effects of salt stress on amino acid, organic acid, and carbohydrate composition of roots, bacteroids, and cytosol of alfalfa (Medicago sativa L.). Plant Physiol. 1991, 96, 1228–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gay, F.; Maraval, I.; Roques, S.; Gunata, Z.; Boulanger, R.; Audebert, A.; Mestres, C. Effect of salinity on yield and 2-acetyl-1-pyrroline content in the grains of three fragrant rice cultivars (Oryza sativa L.) in Camargue (France). Field Crops Res. 2010, 117, 154–160. [Google Scholar] [CrossRef]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, M.; Zeier, T.; Bernsdorff, F.; Reichel-Deland, V.; Kim, D.; Hohmann, M.; Scholten, N.; Schuck, S.; Bräutigam, A.; Hölzel, T.J.C. Flavin monooxygenase-generated N-hydroxypipecolic acid is a critical element of plant systemic immunity. Cell 2018, 173, 456–469.e416. [Google Scholar] [CrossRef] [Green Version]
- Hartzendorf, T.; Rolletschek, H.J.A.B. Effects of NaCl-salinity on amino acid and carbohydrate contents of Phragmites australis. Aquat. Bot. 2001, 69, 195–208. [Google Scholar] [CrossRef]
- Huang, L.; Wu, D.-z.; Zhang, G.-p. Advances in studies on ion transporters involved in salt tolerance and breeding crop cultivars with high salt tolerance. J. Zhejiang Univ. -Sci. B 2020, 21, 426–441. [Google Scholar] [CrossRef]
- Ji, H.; Yang, G.; Zhang, X.; Zhong, Q.; Qi, Y.; Wu, K.; Shen, T.J.F.i.P.S. Regulation of salt tolerance in the roots of Zea mays by L-histidine through transcriptome analysis. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Joseph, E.; Radhakrishnan, V.; Mohanan, K. A study on the accumulation of proline-an osmoprotectant amino acid under salt stress in some native rice cultivars of North Kerala, India. Univers. J. Agric. Res. 2015, 3, 15–22. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E.J.F.c. Contribution of amino acids to strawberry fruit quality and their relevance as stress indicators under NaCl salinity. Food Chem. 2008, 111, 642–647. [Google Scholar] [CrossRef]
- Kobayashi, H.; Sato, S.; Masaoka, Y.J.P.P.S. Tolerance of grasses to calcium chloride, magnesium chloride and sodium chloride. Plant Prod. Sci. 2004, 7, 30–35. [Google Scholar] [CrossRef]
- Latz, A.; Mehlmer, N.; Zapf, S.; Mueller, T.D.; Wurzinger, B.; Pfister, B.; Csaszar, E.; Hedrich, R.; Teige, M.; Becker, D. Salt stress triggers phosphorylation of the Arabidopsis vacuolar K+ channel TPK1 by calcium-dependent protein kinases (CDPKs). Mol. Plant 2013, 6, 1274–1289. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-C.; Pyeon, H.-Y.; Park, Y.-J.; Oh, T.-S.; Jang, M.-J.J.J.o.M. Effect of sodium chloride on the growth, amino acid content, and fragrance patterns of Pleurotus ostreatus. J. Mushroom 2021, 19, 310–315. [Google Scholar]
- Lindemann, B.J.N. Receptors and transduction in taste. Nature 2001, 413, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, B.; Li, J.; Song, Z.; Lu, B.; Chi, M.; Yang, B.; Liu, J.; Lam, Y.-W.; Li, J. Salt-response analysis in two rice cultivars at seedling stage. Acta Physiol. Plant. 2017, 39, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, F.J.; Amtmann, A.J.A.o.b. K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Mansour, M.J. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant. 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Protection of plasma membrane of onion epidermal cells by glycinebetaine and proline against NaCl stress. Plant Physiol. Biochem. 1998, 36, 767–772. [Google Scholar] [CrossRef]
- Mouhamad, R.S.; Iqbal, M.; Qamar, M.A.; Mutlag, L.A.; Razaq, I.B.; Abbas, M.; Hussain, F. Effect of gravistimulation on amino acid profile of pea, rice, corn, wheat during early growth stages. Inf. Process. Agric. 2016, 3, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Nan, G.; Zhang, Y.; Li, S.; Lee, I.; Takano, T.; Liu, S. NaCl stress-induced transcriptomics analysis of Salix linearistipularis (syn. Salix mongolica). J. Biol. Res. -Thessalon. 2016, 23, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Nedjimi, B.J.C.S. How NaCl, Na2SO4, MgCl2 and CaCl2 salts affect the germinability of Pinus halepensis mill. Curr. Sci. 2017, 2031–2035. [Google Scholar] [CrossRef]
- Nedjimi, B.J.J.o.t.S.S.o.A. S Is salinity tolerance related to osmolytes accumulation in Lygeum spartum L. seedlings? J. Saudi Soc. Agric. Sci. 2011, 10, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Nongpiur, R.; Soni, P.; Karan, R.; Singla-Pareek, S.L.; Pareek, A.J.P.s. Behavior. Histidine kinases in plants: Cross talk between hormone and stress responses. Plant Signal. Behav. 2012, 7, 1230–1237. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.K.; Kumar, M.; Li, W.; Luo, Y.; Burritt, D.J.; Alkan, N.; Tran, L.-S.P. Enhancing salt tolerance of plants: From metabolic reprogramming to exogenous chemical treatments and molecular approaches. Cells 2020, 9, 2492. [Google Scholar] [CrossRef] [PubMed]
- Pires, M.V.; Pereira Júnior, A.A.; Medeiros, D.B.; Daloso, D.M.; Pham, P.A.; Barros, K.A.; Engqvist, M.K.; Florian, A.; Krahnert, I.; Maurino, V.G.J.P.; et al. The influence of alternative pathways of respiration that utilize branched-chain amino acids following water shortage in Arabidopsis. Plant Cell Environ. 2016, 39, 1304–1319. [Google Scholar] [CrossRef] [Green Version]
- Rare, E.J.J.o.H.S. Stress physiology: The functional significance of the accumulation of nitrogen-containing compounds. J. Hortic. Sci. 1990, 65, 231–243. [Google Scholar] [CrossRef]
- Silveira, J.A.G.; de Almeida Viégas, R.; da Rocha, I.M.A.; Moreira, A.C.d.O.M.; de Azevedo Moreira, R.; Oliveira, J.T.A. Proline accumulation and glutamine synthetase activity are increased by salt-induced proteolysis in cashew leaves. J. Plant Physiol. 2003, 160, 115–123. [Google Scholar] [CrossRef]
- Simon-Sarkadi, L.; Kocsy, G.; Sebestyén, Z.J.A.B.S. Effect of salt stress on free amino acid and polyamine content in cereals. Acta Biol. Szeged. 2002, 46, 73–75. [Google Scholar]
- Storey, R.; Walker, R.R. Citrus and salinity. Sci. Hortic. 1999, 78, 39–81. [Google Scholar] [CrossRef]
- Su, Q.; Zheng, X.; Tian, Y.; Wang, C. Exogenous brassinolide alleviates salt stress in Malus hupehensis Rehd. by regulating the transcription of NHX-Type Na+ (K+)/H+ antiporters. Front. Plant Sci. 2020, 11, 38. [Google Scholar] [CrossRef]
- Teakle, N.L.; Tyerman, S.D.J.P. Mechanisms of Cl-transport contributing to salt tolerance. Plant Cell Environ. 2010, 33, 566–589. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Davenport, R.J.A.o.B. Na⁺ tolerance and Na⁺ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Toderich, K.N.; Mamadrahimov, A.A.; Khaitov, B.B.; Karimov, A.A.; Soliev, A.A.; Nanduri, K.R.; Shuyskaya, E.V.J. Differential impact of salinity stress on seeds minerals, storage proteins, fatty acids, and squalene composition of new Quinoa genotype, grown in hyper-arid desert environments. Front. Plant Sci. 2020, 11, 607102. [Google Scholar] [CrossRef]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Xie, E.; Wei, X.; Ding, A.; Zheng, L.; Wu, X.; Anderson, B. Short-term effects of salt stress on the amino acids of Phragmites australis root exudates in constructed wetlands. Water 2020, 12, 569. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Park, J.-R.; Jang, Y.-H.; Kim, E.-G.; Kim, K.-M.J. Rice cultivars under salt stress Show differential expression of genes related to the regulation of Na+/K+ balance. Front. Plant Sci. 2021, 12, 680131. [Google Scholar] [CrossRef]
- Saranga, Y.; Rhodes, D.; Janick, J. Changes in amino acid composition associated with tolerance to partial desiccation of celery somatic embryos. J. Am. Soc. Hortic. Sci. 1992, 117, 337–341. [Google Scholar] [CrossRef]
- Okuma, E.; Soeda, K.; Tada, M.; Murata, Y. Exogenous proline mitigates the inhibition of growth of Nicotiana tabacum cultured cells under saline conditions. Soil Sci. Plant Nutr. 2000, 46, 257–263. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Amino Acids Concentration (mg/g) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Amino Acids | Control | NaCl | CaCl2 | MgCl2 | |||||
| histidine | 21.9320 | ±0.0481 a | 22.3443 | ±0.0572 a | 18.6910 | ±0.0530 b | 26.2183 | ±0.0780 b | |
| isoleucine | 21.5630 | ±0.0278 a | 23.0637 | ±0.0405 a | 20.8033 | ±0.13930 b | 20.8923 | ±0.0810 a | |
| Cheongcheong | leucine | 47.5327 | ±0.0476 d | 49.0367 | ±0.0449 d | 42.1950 | ±0.0040 b | 56.7487 | ±0.0366 d |
| lysine | 47.5327 | ±0.0476 a | 49.0367 | ±0.0449 a | 42.1950 | ±0.0040 b | 56.7487 | ±0.0366 a | |
| methionine | 11.4197 | ±0.0830 a | 13.0113 | ±0.0080 a | 13.6030 | ±0.0538 b | 10.6220 | ±0.0726 a | |
| phenylalanine | 50.9810 | ±0.0135 a | 49.5547 | ±0.06824 a | 46.3273 | ±0.0479 b | 62.3823 | ±0.0910 a | |
| valine | 31.4523 | ±0.0605 a | 32.3437 | ±0.4789 a | 27.0763 | ±0.05525 b | 38.2525 | ±0.2092 a | |
| histidine | 12.4530 | ±0.0416 d | 13.0277 | ±0.0670 b | 20.2657 | ±0.0606 a | 18.4313 | ±0.0504 b | |
| isoleucine | 42.1503 | ±0.0513 d | 47.3487 | ±0.1185 c | 11.6133 | ±0.0671 a | 23.1907 | ±0.0082 b | |
| Nagdong | leucine | 28.3263 | ±0.0971 c | 30.7597 | ±0.0577 a | 48.7727 | ±0.0193 c | 40.1517 | ±0.0504 b |
| lysine | 28.3263 | ±0.097 d | 30.7597 | ±0.0577 c | 48.7727 | ±0.0193 a | 40.1517 | ±0.0504 b | |
| methionine | 6.1750 | ±0.01840 b | 8.8327 | ±0.0499 b | 14.3833 | ±0.4374 a | 5.5293 | ±0.0510 c | |
| phenylalanine | 27.4770 | ±0.0558 d | 36.6877 | ±0.0279 b | 62.4527 | ±0.0435 a | 40.8457 | ±0.0409 b | |
| valine | 17.7271 | ±0.0826 d | 18.5057 | ±0.0813 c | 28.9400 | ±0.0519 a | 26.3620 | ±0.1285 b | |
| histidine | 17.3273 | ±0.0918 b | 13.5823 | ±0.4245 c | 0.1331 | ±0.1484 d | 17.9267 | ±0.0526 c | |
| isoleucine | 58.6427 | ±0.0816 b | 46.9240 | ±0.0580 b | 8.5097 | ±0.0875 d | 20.4000 | ±0.0523 c | |
| IR28 | leucine | 38.6567 | ±0.0405 a | 31.2697 | ±0.0448 b | 0.1057 | ±0.0024 d | 38.8330 | ±0.0480 c |
| lysine | 38.6567 | ±0.0405 b | 31.2697 | ±0.0448 b | 0.1057 | ±0.0024 d | 38.8330 | ±0.0480 c | |
| methionine | 4.8377 | ±0.0604 c | 7.5853 | ±0.0221 c | 0.3053 | ±0.0026 d | 7.1460 | ±0.0388 b | |
| phenylalanine | 35.6660 | ±0.0340 b | 30.5953 | ±0.0244 c | 0.9770 | ±0.0128 d | 37.3160 | ±0.0742 c | |
| valine | 25.3173 | ±0.0696 b | 20.1277 | ±0.2022 b | 1.3500 | ±0.3846 d | 25.6160 | ±0.0638 c | |
| histidine | 14.4810 | ±0.0216 c | 10.1870 | ±0.0992 d | 10.4380 | ±0.0736 c | 12.4347 | ±0.0691 d | |
| isoleucine | 48.5057 | ±0.0813 c | 34.4333 | ±0.0689 d | 35.0070 | ±0.0016 c | 43.2627 | ±0.1078 d | |
| Pokkali | leucine | 31.7527 | ±0.0205 b | 23.6393 | ±0.0436 c | 23.3773 | ±0.0469 a | 28.4547 | ±0.0406 a |
| lysine | 31.7527 | ±0.0205 c | 23.6393 | ±0.0436 d | 23.3773 | ±0.0466 c | 28.4547 | ±0.0406 d | |
| methionine | 2.6607 | ±0.0372 d | 1.8703 | ±0.0389 d | 2.7887 | ±0.1329 c | 3.6467 | ±0.0428 d | |
| phenylalanine | 30.1867 | ±0.0099 c | 24.1847 | ±0.0108 d | 22.9813 | ±0.0256 c | 29.4773 | ±0.0167 d | |
| valine | 20.9020 | ±0.1092 c | 15.0550 | ±0.1042 d | 15.0603 | ±0.0998 c | 17.9817 | ±0.0146 d | |
| Amino Acids Concentration (mg/g) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Amino Acids | Control | NaCl | CaCl2 | MgCl2 | |||||
| alanine | 3.8897 | ±0.09458 b | 5.8680 | ±0.1182 a | 6.3473 | ±0.0849 a | 8.8637 | ±0.1219 a | |
| aspartic acid | 0.0000 | ±0.0000 d | 0.0120 | ±0.0008 d | 0.0280 | ±0.0008 c | 12.6287 | ±0.0542 a | |
| arginine | 18.5940 | ±0.0077 d | 18.8767 | ±0.0205 d | 13.6407 | ±0.0073 b | 20.2863 | ±0.0383 b | |
| Cheongcheong | cystine | 6.0297 | ±0.0499 b | 6.8760 | ±0.1232 a | 5.7897 | ±0.1353 c | 17.7533 | ±0.2000 a |
| glutamic acid | 2.0557 | ±0.0328 b | 1.9530 | ±0.0751 b | 1.0663 | ±0.0407 c | 79.3577 | ±0.4542 a | |
| proline | 67.4487 | ±0.3886 b | 68.3933 | ±0.0062 a | 60.4013 | ±0.0849 b | 180.5450 | ±0.0666 a | |
| serine | 30.5667 | ±0.0262 a | 32.5203 | ±0.2946 a | 26.5350 | ±0.1382 b | 5.6793 | ±0.1636 c | |
| tyrosine | 32.7213 | ±0.0886 a | 30.8024 | ±0.0853 a | 31.3510 | ±0.0753 c | 41.6530 | ±0.0654 a | |
| alanine | 2.7193 | ±0.1678 c | 3.9213 | ±0.0575 b | 4.4190 | ±0.0686 b | 1.8937 | ±0.0869 d | |
| aspartic acid | 0.0943 | ±0.0030 c | 0.0297 | ±0.0012 c | 0.0400 | ±0.0008 b | 0.3170 | ±0.0008 b | |
| arginine | 26.8177 | ±0.1422 c | 31.1870 | ±0.0220 a | 22.9353 | ±0.0719 a | 16.9047 | ±0.0977 c | |
| Nagdong | cystine | 3.4220 | ±0.1742 c | 4.6633 | ±0.1347 c | 8.0563 | ±0.1023 b | 7.9863 | ±0.0344 b |
| glutamic acid | 0.8600 | ±0.0445 d | 0.8253 | ±0.0592 c | 1.9667 | ±0.0286 b | 50.7533 | ±0.2684 b | |
| proline | 37.1363 | ±0.0310 d | 42.9877 | ±0.0167 b | 67.5527 | ±0.1063 a | 0.4480 | ±0.0490 c | |
| serine | 17.7547 | ±0.0937 c | 17.7397 | ±0.2226 c | 28.4620 | ±0.0602 a | 46.1547 | ±0.2519 a | |
| tyrosine | 17.7763 | ±0.0459 d | 22.8867 | ±0.0294 b | 38.0600 | ±0.0432 a | 26.9487 | ±0.0365 b | |
| alanine | 6.3427 | ±0.3871 a | 1.5003 | ±0.3229 d | 0.0600 | ±0.0163 d | 0.7853 | ±0.1222 c | |
| aspartic acid | 7.5447 | ±0.0054 a | 0.3880 | ±0.0008 b | 0.0022 | ±0.0001 d | 0.2353 | ±0.0036 c | |
| arginine | 37.9410 | ±0.0820 a | 30.3133 | ±0.0981 b | 0.2370 | ±0.0453 c | 13.4747 | ±0.0850 d | |
| IR28 | cystine | 7.4310 | ±0.0488 a | 5.0977 | ±0.0036 b | 64.0577 | ±0.1019 a | 5.1280 | ±0.0523 c |
| glutamic acid | 51.8383 | ±0.1194 a | 2.5927 | ±0.2237 a | 23.0097 | ±0.0725 a | 12.8890 | ±0.0954 c | |
| proline | 114.3420 | ±0.1197 a | 39.5077 | ±0.0825 c | 46.3053 | ±0.0849 c | 0.0073 | ±0.0012 d | |
| serine | 0.0480 | ±0.0021 d | 19.5293 | ±0.3328 b | 23.0097 | ±0.0725 c | 46.1023 | ±0.0761 a | |
| tyrosine | 22.4483 | ±0.0767 b | 20.9170 | ±0.0794 c | 36.3613 | ±0.0704 b | 25.0637 | ±0.0209 c | |
| alanine | 1.8300 | ±0.0973 d | 2.9263 | ±0.0547 c | 2.9950 | ±0.0063 c | 4.4933 | ±0.1069 b | |
| aspartic acid | 1.3100 | ±0.0488 b | 2.3160 | ±0.0024 a | 0.1417 | ±0.0012 a | 0.3000 | ±0.0008 b | |
| arginine | 31.7940 | ±0.1709 b | 22.3023 | ±0.0964 c | 22.9140 | ±0.1037 a | 27.8323 | ±0.0985 a | |
| Pokkali | cystine | 1.4320 | ±0.0662 d | 1.9943 | ±0.0172 d | 2.3770 | ±0.0910 d | 1.7170 | ±0.0608 d |
| glutamic acid | 1.5097 | ±0.0440 c | 0.7767 | ±0.1247 c | 0.4673 | ±0.0529 d | 0.7697 | ±0.0865 d | |
| proline | 41.5357 | ±0.0484 c | 31.6633 | ±0.0262 d | 31.3440 | ±0.0910 d | 38.3803 | ±0.0873 b | |
| serine | 21.3560 | ±0.0881 b | 13.8670 | ±0.1142 d | 14.5700 | ±0.0778 d | 17.0800 | ±0.0081 b | |
| tyrosine | 18.2497 | ±0.0406 c | 15.0227 | ±0.0165 d | 14.9750 | ±0.0094 d | 18.7513 | ±0.0433 d | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, M.; Jang, Y.-H.; Kim, E.-G.; Park, J.-R.; Eom, G.-H.; Zhao, D.-D.; Kim, K.-M. Evaluation of Amino Acid Profiles of Rice Genotypes under Different Salt Stress Conditions. Plants 2023, 12, 1315. https://doi.org/10.3390/plants12061315
Farooq M, Jang Y-H, Kim E-G, Park J-R, Eom G-H, Zhao D-D, Kim K-M. Evaluation of Amino Acid Profiles of Rice Genotypes under Different Salt Stress Conditions. Plants. 2023; 12(6):1315. https://doi.org/10.3390/plants12061315
Chicago/Turabian StyleFarooq, Muhammad, Yoon-Hee Jang, Eun-Gyeong Kim, Jae-Ryoung Park, Gyu-Hyeon Eom, Dan-Dan Zhao, and Kyung-Min Kim. 2023. "Evaluation of Amino Acid Profiles of Rice Genotypes under Different Salt Stress Conditions" Plants 12, no. 6: 1315. https://doi.org/10.3390/plants12061315