
Genome-Wide Identification and Evolutionary Analysis of Gossypium YTH Domain-Containing RNA-Binding Protein Family and the Role of GhYTH8 in Response to Drought Stress
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. YTH Gene Family in Four Gossypium Species
2.2. Phylogenetic Analysis of YTH Genes
2.3. Motifs and Gene Structure Analysis
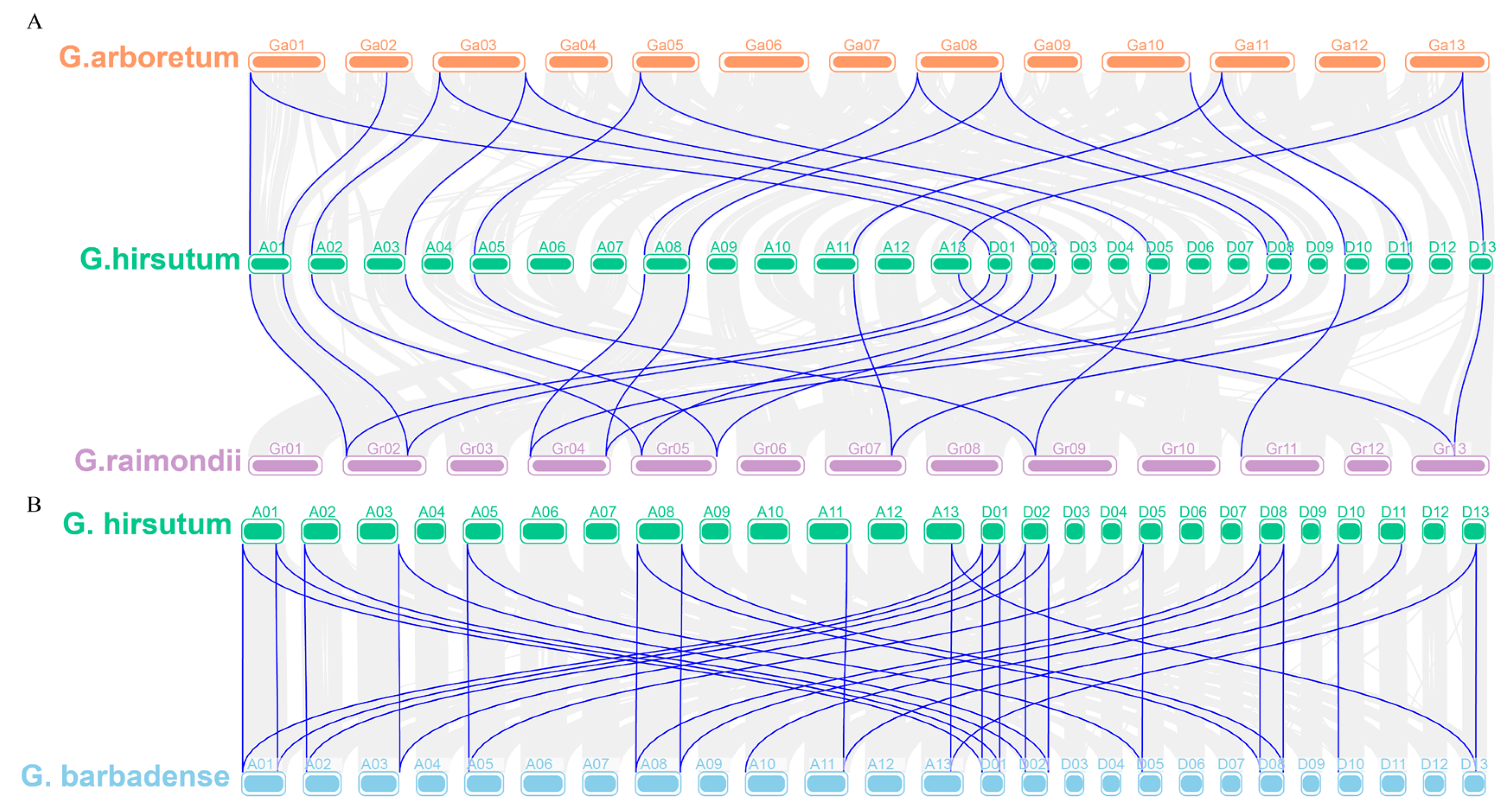
2.4. Chromosome Distribution and Synteny Analysis of YTH Genes
2.5. Cis-Regulatory Element Analysis of YTH Genes
2.6. Prediction of YTH Genes Targeted by MiRNAs in G. Hirsutum
2.7. Tissue Expression Patterns of YTH Genes in G. hirsutum
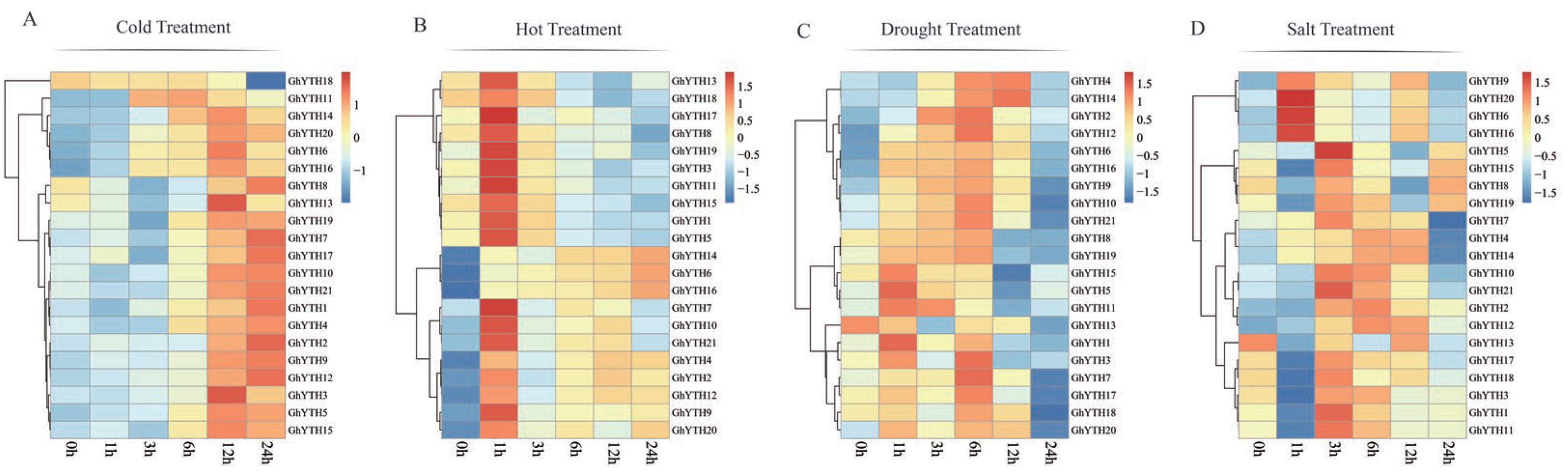
2.8. Expression Patterns of YTH Genes in G. hirsutum under Stress Conditions
2.9. Subcellular Localization of GhYTH8 and GhYTH16
2.10. Functional Verification of GhYTH8 in Cotton under Drought Stress
3. Discussion
4. Materials and Methods
4.1. Identification and Physiochemical Properties Analysis of YTH Gene Family Members in Four Gossypium Species
4.2. Evolutionary, Gene Structure, and Conserved Motif Analysis of YTH Genes
4.3. Chromosomal Distribution and Synteny Analysis of YTH Genes
4.4. Regulatory Elements Analysis of YTH Genes
4.5. Subcellular Localization of GhYTH8 and GhYTH16 Proteins
4.6. Expression Profiles Analysis of YTH Genes
4.7. MiRNA Prediction of GhYTH Genes
4.8. Virus-induced Gene Silencing (VIGS) Experiment and Drought Treatment
4.9. Quantitative Real-Time PCR (qPCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Glisovic, T.; Bachorik, J.L.; Yong, J.; Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008, 582, 1977–1986. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Rio, D.C. Mechanisms and Regulation of Alternative Pre-mRNA Splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [Green Version]
- Mittal, N.; Scherrer, T.; Gerber, A.P.; Janga, S.C. Interplay between posttranscriptional and posttranslational interactions of RNA-binding proteins. J. Mol. Biol. 2011, 409, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Lorkovic, Z.J. Role of plant RNA-binding proteins in development, stress response and genome organization. Trends Plant Sci. 2009, 14, 229–236. [Google Scholar] [CrossRef]
- Ambrosone, A.; Costa, A.; Leone, A.; Grillo, S. Beyond transcription: RNA-binding proteins as emerging regulators of plant response to environmental constraints. Plant Sci. 2012, 182, 12–18. [Google Scholar] [CrossRef]
- Imai, Y.; Matsuo, N.; Ogawa, S.; Tohyama, M.; Takagi, T. Cloning of a gene, YT521, for a novel RNA splicing-related protein induced by hypoxia/reoxygenation. Mol. Brain Res. 1998, 53, 33–40. [Google Scholar] [CrossRef]
- Hartmann, A.M.; Nayler, O.; Schwaiger, F.W.; Obermeier, A.; Stamm, S. The interaction and colocalization of Sam68 with the splicing-associated factor YT521-B in nuclear dots is regulated by the Src family kinase p59fyn. Mol. Biol. Cell 1999, 10, 3909–3926. [Google Scholar] [CrossRef] [Green Version]
- Stoilov, P.; Rafalska, I.; Stamm, S. YTH: A new domain in nuclear proteins. Trends Biochem. Sci. 2002, 27, 495–497. [Google Scholar] [CrossRef]
- Addepalli, B.; Hunt, A.G. A novel endonuclease activity associated with the Arabidopsis ortholog of the 30-kDa subunit of cleavage and polyadenylation specificity factor. Nucleic. Acids Res. 2007, 35, 4453–4463. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 176. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Hsu, P.J.; Chen, Y.S.; Yang, Y.G. Dynamic transcriptomic m(6)A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, X.; Liu, K.; Roundtree, I.A.; Tempel, W.; Li, Y.; Lu, Z.; He, C.; Min, J. Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat. Chem. Biol. 2014, 10, 927–929. [Google Scholar] [CrossRef]
- Li, D.; Zhang, H.; Hong, Y.; Huang, L.; Li, X.; Zhang, Y.; Ouyang, Z.; Song, F. Genome-Wide Identification, Biochemical Characterization, and Expression Analyses of the YTH Domain-Containing RNA-Binding Protein Family in Arabidopsis and Rice. Plant Mol. Biol. Report. 2014, 32, 1169–1186. [Google Scholar] [CrossRef]
- Wang, N.; Yue, Z.; Liang, D.; Ma, F. Genome-wide identification of members in the YTH domain-containing RNA-binding protein family in apple and expression analysis of their responsiveness to senescence and abiotic stresses. Gene 2014, 538, 292–305. [Google Scholar] [CrossRef]
- Yin, S.; Ao, Q.; Tan, C.; Yang, Y. Genome-wide identification and characterization of YTH domain-containing genes, encoding the m(6)A readers, and their expression in tomato. Plant Cell Rep. 2021, 40, 1229–1245. [Google Scholar] [CrossRef]
- Sun, J.; Bie, X.M.; Wang, N.; Zhang, X.S.; Gao, X.Q. Genome-wide identification and expression analysis of YTH domain-containing RNA-binding protein family in common wheat. BMC Plant Biol. 2020, 20, 351. [Google Scholar] [CrossRef]
- Arribas-Hernandez, L.; Bressendorff, S.; Hansen, M.H.; Poulsen, C.; Erdmann, S.; Brodersen, P. An m(6)A-YTH Module Controls Developmental Timing and Morphogenesis in Arabidopsis. Plant Cell 2018, 30, 952–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scutenaire, J.; Deragon, J.M.; Jean, V.; Benhamed, M.; Raynaud, C.; Favory, J.J.; Merret, R.; Bousquet-Antonelli, C. The YTH Domain Protein ECT2 Is an m(6)A Reader Required for Normal Trichome Branching in Arabidopsis. Plant Cell 2018, 30, 986–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, S.; Ao, Q.; Qiu, T.; Tan, C.; Tu, Y.; Kuang, T.; Yang, Y. Tomato SlYTH1 encoding a putative RNA m(6)A reader affects plant growth and fruit shape. Plant Sci. 2022, 323, 111417. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Guo, T.; Wang, P.; Sun, X.; Shao, Y.; Jia, X.; Liang, B.; Gong, X.; Ma, F. MhYTP1 and MhYTP2 from Apple Confer Tolerance to Multiple Abiotic Stresses in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1367. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Hu, L.; Jiang, L.; Liu, S. Genome-wide identification and expression analysis of YTH domain-containing RNA-binding protein family in cucumber (Cucumis sativus). Genes Genom. 2018, 40, 579–589. [Google Scholar] [CrossRef]
- Biłas, R.; Szafran, K.; Hnatuszko-Konka, K.; Kononowicz, A.K. Cis-regulatory elements used to control gene expression in plants. Plant Cell Tissue Organ Cult. 2016, 127, 269–287. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Li, Y.; Zhao, W.; Song, X.; Lu, M.; Li, X.; Li, X.; Liu, R.; Yan, L.; Zhang, X. Integration of Hormonal and Nutritional Cues Orchestrates Progressive Corolla Opening. Plant Physiol. 2016, 171, 1209–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Fan, G.; Wang, K.; Sun, F.; Yuan, Y.; Song, G.; Li, Q.; Ma, Z.; Lu, C.; Zou, C.; et al. Genome sequence of the cultivated cotton Gossypium arboreum. Nat. Genet. 2014, 46, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Chen, J.; Fang, L.; Zhang, Z.; Ma, W.; Niu, Y.; Ju, L.; Deng, J.; Zhao, T.; Lian, J.; et al. Gossypium barbadense and Gossypium hirsutum genomes provide insights into the origin and evolution of allotetraploid cotton. Nat. Genet. 2019, 51, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Lv, W.; Li, T.; Zhang, S.; Wang, H.; Li, X.; Wang, L.; Ma, D.; Zang, Y.; Shen, J.; et al. Dynamic regulation and functions of mRNA m6A modification. Cancer Cell Int. 2022, 22, 48. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Sun, H.; Xu, C. YTH Domain: A Family of N(6)-methyladenosine (m(6)A) Readers. Genom. Proteom. Bioinform. 2018, 16, 99–107. [Google Scholar] [CrossRef]
- Patil, D.P.; Pickering, B.F.; Jaffrey, S.R. Reading m(6)A in the Transcriptome: M(6)A-Binding Proteins. Trends Cell Biol. 2018, 28, 113–127. [Google Scholar] [CrossRef]
- Chen, Z.J.; Scheffler, B.E.; Dennis, E.; Triplett, B.A.; Zhang, T.; Guo, W.; Chen, X.; Stelly, D.M.; Rabinowicz, P.D.; Town, C.D.; et al. Toward sequencing cotton (Gossypium) genomes. Plant Physiol. 2007, 145, 1303–1310. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Huang, G.; He, S.; Yang, Z.; Sun, G.; Ma, X.; Li, N.; Zhang, X.; Sun, J.; Liu, M.; et al. Resequencing of 243 diploid cotton accessions based on an updated A genome identifies the genetic basis of key agronomic traits. Nat. Genet. 2018, 50, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Morton, J.A.; Pellicer, J.; Leitch, I.J.; Leitch, A.R. Genome downsizing after polyploidy: Mechanisms, rates and selection pressures. Plant J. 2021, 107, 1003–1015. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014, 14, 93. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Heidari, P.; Faraji, S.; Ahmadizadeh, M.; Ahmar, S.; Mora-Poblete, F. New Insights Into Structure and Function of TIFY Genes in Zea mays and Solanum lycopersicum: A Genome-Wide Comprehensive Analysis. Front. Genet. 2021, 12, 657970. [Google Scholar] [CrossRef]
- Abdullah; Faraji, S.; Mehmood, F.; Malik, H.M.T.; Ahmed, I.; Heidari, P.; Poczai, P. The GASA Gene Family in Cacao (Theobroma cacao, Malvaceae): Genome Wide Identification and Expression Analysis. Agronomy 2021, 11, 1425. [Google Scholar] [CrossRef]
- Rejeb, I.B.; Pastor, V.; Mauch-Mani, B. Plant Responses to Simultaneous Biotic and Abiotic Stress: Molecular Mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef] [Green Version]
- Bhargava, S.; Sawant, K.; Tuberosa, R. Drought stress adaptation: Metabolic adjustment and regulation of gene expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
- Sheshadri, S.A.; Nishanth, M.J.; Simon, B. Stress-Mediated cis-Element Transcription Factor Interactions Interconnecting Primary and Specialized Metabolism in Planta. Front. Plant Sci. 2016, 7, 1725. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.; Liang, C.; Meng, Z.; Sun, G.; Meng, Z.; Guo, S.; Zhang, R. CottonFGD: An integrated functional genomics database for cotton. BMC Plant Biol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Wang, N.; Xue, Y.; Guan, Q.; van Nocker, S.; Liu, C.; Ma, F. Overexpression of the RNA binding protein MhYTP1 in transgenic apple enhances drought tolerance and WUE by improving ABA level under drought condition. Plant Sci. 2019, 280, 397–407. [Google Scholar] [CrossRef]
- Niu, D.; Zhang, X.; Song, X.; Wang, Z.; Li, Y.; Qiao, L.; Wang, Z.; Liu, J.; Deng, Y.; He, Z.; et al. Deep Sequencing Uncovers Rice Long siRNAs and Its Involvement in Immunity against Rhizoctonia solani. Phytopathology 2018, 108, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. Drought coping strategies in cotton: Increased crop per drop. Plant Biotechnol. J. 2017, 15, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Wang, Q.; Sun, R.; Zhang, B. Deep sequencing reveals important roles of microRNAs in response to drought and salinity stress in cotton. J. Exp. Bot. 2015, 66, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Pan, A.; He, S.; Su, P.; Yuan, X.; Zhu, S.; Liu, Z. Different MicroRNA Families Involved in Regulating High Temperature Stress Response during Cotton (Gossypium hirsutum L.) Anther Development. Int. J. Mol. Sci. 2020, 21, 1280. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Ying, S.; Li, Y.; Zhu, L.; Wang, X.; Jin, H. Linking the YTH domain to cancer: The importance of YTH family proteins in epigenetics. Cell Death Dis. 2021, 12, 346. [Google Scholar] [CrossRef]
- Hirschfeld, M.; Zhang, B.; Jaeger, M.; Stamm, S.; Erbes, T.; Mayer, S.; Tong, X.; Stickeler, E. Hypoxia-dependent mRNA expression pattern of splicing factor YT521 and its impact on oncological important target gene expression. Mol. Carcinog. 2014, 53, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Qin, W.; Li, Y.; Zhang, C.; Wang, Y.; Yang, Z.; Ge, X.; Li, F. Red light promotes cotton embryogenic callus formation by influencing endogenous hormones, polyamines and antioxidative enzyme activities. Plant Growth Regul. 2018, 87, 187–199. [Google Scholar] [CrossRef]
- Arabidopsis Genome, I. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Sharma, H.; Sharma, A.; Rajput, R.; Sidhu, S.; Dhillon, H.; Verma, P.C.; Pandey, A.; Upadhyay, S.K. Molecular Characterization, Evolutionary Analysis, and Expression Profiling of BOR Genes in Important Cereals. Plants 2022, 11, 911. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [Green Version]
- Otasek, D.; Morris, J.H.; Boucas, J.; Pico, A.R.; Demchak, B. Cytoscape Automation: Empowering workflow-based network analysis. Genome. Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Amino Acid Length | P.I. | MW (da) | Location |
|---|---|---|---|---|---|
| Ga01G0324.1 | GaYTH1 | 654 | 6.19 | 72224.25 | Chr01:2562085–2564534(-) |
| Ga02G0908.1 | GaYTH2 | 625 | 6.83 | 68261.89 | Chr02:60033655–60039884(-) |
| Ga03G0644.1 | GaYTH3 | 654 | 5.83 | 72141.35 | Chr03:9431937–9434567(-) |
| Ga03G2762.1 | GaYTH4 | 572 | 6.08 | 63500.31 | Chr03:135380859–135385216(-) |
| Ga08G0210.1 | GaYTH5 | 635 | 7.22 | 70257.79 | Chr08:1723259–1728028(-) |
| Ga08G2483.1 | GaYTH6 | 577 | 5.9 | 63763.03 | Chr08:124586293–124589173(+) |
| Ga10G3082.1 | GaYTH7 | 648 | 5.32 | 71361.54 | Chr10:129290100–129295676(-) |
| Ga11G0956.1 | GaYTH8 | 677 | 5.56 | 74319.92 | Chr11:16221680–16226718(+) |
| Ga11G0958.1 | GaYTH9 | 605 | 6.15 | 66573.51 | Chr11:16288205–16291213(+) |
| Ga13G1351.1 | GaYTH10 | 596 | 6.2 | 65509.66 | Chr13:83803610–83807221(-) |
| Gbar_A01G002830.1 | GbYTH1 | 566 | 8.48 | 62324.52 | A01:2568244–2572387(-) |
| Gbar_A01G015130.1 | GbYTH2 | 686 | 6.56 | 75281.04 | A01:91702309–91709506(+) |
| Gbar_A02G005740.2 | GbYTH3 | 654 | 5.92 | 72218.39 | A02:8400243–8404612(-) |
| Gbar_A03G023470.1 | GbYTH4 | 572 | 5.96 | 63511.37 | A03:105084247–105089973(-) |
| Gbar_A05G011330.1 | GbYTH5 | 653 | 5.2 | 71651.11 | A05:10378466–10385178(-) |
| Gbar_A08G001890.1 | GbYTH6 | 666 | 6.09 | 73438.17 | A08:1669319–1673433(-) |
| Gbar_A08G022910.1 | GbYTH7 | 577 | 6.01 | 63687.94 | A08:115214249–115217627(+) |
| Gbar_A10G000210.1 | GbYTH8 | 650 | 5.32 | 71521.71 | A10:226672–232918(+) |
| Gbar_A11G027540.5 | GbYTH9 | 605 | 6.15 | 66528.45 | A11:98904170–98908097(-) |
| Gbar_A11G027580.1 | GbYTH10 | 612 | 5.96 | 66900.74 | A11:98946062–98949702(-) |
| Gbar_A13G011910.1 | GbYTH11 | 607 | 6.08 | 66295.32 | A13:73997171–74001401(-) |
| Gbar_D01G003000.1 | GbYTH12 | 654 | 7.22 | 72240.22 | D01:2591629–2595737(-) |
| Gbar_D01G016180.1 | GbYTH13 | 625 | 6.72 | 68475.31 | D01:47823428–47830617(+) |
| Gbar_D02G006470.1 | GbYTH14 | 689 | 8.27 | 76200.42 | D02:8464041–8470880(-) |
| Gbar_D02G025310.1 | GbYTH15 | 572 | 6.21 | 63284.11 | D02:67385264–67390933(-) |
| Gbar_D05G011770.9 | GbYTH16 | 660 | 5.04 | 72234.67 | D05:9785369–9791757(-) |
| Gbar_D08G001930.1 | GbYTH17 | 670 | 6 | 73907.66 | D08:1553392–1557467(-) |
| Gbar_D08G023590.1 | GbYTH18 | 559 | 6.81 | 62082.17 | D08:61689230–61692672(+) |
| Gbar_D10G000170.2 | GbYTH19 | 650 | 5.38 | 71582.77 | D10:164189–170278(+) |
| Gbar_D11G035010.1 | GbYTH20 | 622 | 5.72 | 68122.11 | Scaffold118:164271–170796(+) |
| Gbar_D11G035040.1 | GbYTH21 | 605 | 6.26 | 66560.48 | Scaffold118:211996–217213(+) |
| Gbar_D13G011640.1 | GbYTH22 | 606 | 6.08 | 66234.25 | D13:33076872–33081145(-) |
| Gh_A01G032100.1 | GhYTH1 | 654 | 6.19 | 72210.18 | A01:2855623–2858072(-) |
| Gh_A01G173200.1 | GhYTH2 | 625 | 6.55 | 68290.89 | A01:91578115–91584668(+) |
| Gh_A02G065800.1 | GhYTH3 | 654 | 6 | 72278.51 | A02:8801670–8804658(-) |
| Gh_A03G245900.1 | GhYTH4 | 720 | 8.25 | 81093.52 | A03:109862488–109870541(-) |
| Gh_A05G102800.1 | GhYTH5 | 651 | 5.2 | 71407.77 | A05:10898607–10904838(-) |
| Gh_A08G019200.1 | GhYTH6 | 666 | 6.09 | 73404.17 | A08:1725549–1729279(-) |
| Gh_A08G241100.1 | GhYTH7 | 568 | 6 | 62764.8 | A08:120898607–120901810(+) |
| Gh_A11G291200.1 | GhYTH8 | 605 | 6.15 | 66501.43 | A11:105609984–105613426(-) |
| Gh_A11G291500.1 | GhYTH9 | 612 | 5.96 | 66900.74 | A11:105651056–105655185(-) |
| Gh_A13G121400.1 | GhYTH10 | 607 | 6.08 | 66322.34 | A13:73017791–73021817(-) |
| Gh_D01G029500.1 | GhYTH11 | 656 | 7.22 | 72458.49 | D01:2546081–2549106(-) |
| Gh_D01G171300.1 | GhYTH12 | 625 | 6.72 | 68445.22 | D01:49030052–49036273(+) |
| Gh_D02G069800.1 | GhYTH13 | 654 | 6.05 | 71898.2 | D02:8345891–8350344(-) |
| Gh_D02G261200.2 | GhYTH14 | 572 | 6.21 | 63400.27 | D02:70853189–70858785(-) |
| Gh_D05G115100.3 | GhYTH15 | 652 | 5.08 | 71170.49 | D05:9802311–9808568(-) |
| Gh_D08G018500.1 | GhYTH16 | 583 | 7.24 | 64432.55 | D08:1638425–1641033(-) |
| Gh_D08G232400.1 | GhYTH17 | 577 | 6.41 | 64071.27 | D08:64381471–64384820(+) |
| Gh_D10G001600.3 | GhYTH18 | 650 | 5.25 | 71466.59 | D10:196546–202609(+) |
| Gh_D11G291400.1 | GhYTH19 | 605 | 6.26 | 66560.48 | D11:60000676–60004152(-) |
| Gh_D11G291600.1 | GhYTH20 | 613 | 5.86 | 67164.03 | D11:60044437–60048569(-) |
| Gh_D13G123000.1 | GhYTH21 | 606 | 6.12 | 66218.25 | D13:36489013–36493188(-) |
| Gorai.002G031800.1 | GrYTH1 | 654 | 6.9 | 72227.26 | Chr02:2370851–2374924(-) |
| Gorai.002G182900.1 | GrYTH2 | 625 | 6.49 | 68392.05 | Chr02:48153196–48160671(+) |
| Gorai.004G020600.1 | GrYTH3 | 670 | 6 | 73973.81 | Chr04:1530886–1534693(-) |
| Gorai.004G244600.1 | GrYTH4 | 599 | 6.41 | 66812.77 | Chr04:58179219–58182629(+) |
| Gorai.005G070500.1 | GrYTH5 | 654 | 5.96 | 71958.21 | Chr05:7641980–7646032(-) |
| Gorai.005G265300.1 | GrYTH6 | 572 | 6.21 | 63296.12 | Chr05:63865504–63871029(-) |
| Gorai.007G288900.1 | GrYTH7 | 605 | 6.26 | 66610.57 | Chr07:49501978–49505697(-) |
| Gorai.007G289200.1 | GrYTH8 | 622 | 5.69 | 68135.13 | Chr07:49545172–49551745(-) |
| Gorai.009G120700.1 | GrYTH9 | 654 | 5.14 | 71573.99 | Chr09:8935247–8942392(-) |
| Gorai.011G001700.1 | GrYTH10 | 648 | 5.37 | 71416.54 | Chr11:161861–168102(+) |
| Gorai.013G123200.1 | GrYTH11 | 606 | 6.12 | 66162.19 | Chr13:31722245–31726598(-) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, W.; Wang, W.; Xiao, X.; Sun, J.; Wu, B.; Zhao, Y.; Pei, S.; Fan, W.; Xu, D.; Qin, T. Genome-Wide Identification and Evolutionary Analysis of Gossypium YTH Domain-Containing RNA-Binding Protein Family and the Role of GhYTH8 in Response to Drought Stress. Plants 2023, 12, 1198. https://doi.org/10.3390/plants12051198
Hao W, Wang W, Xiao X, Sun J, Wu B, Zhao Y, Pei S, Fan W, Xu D, Qin T. Genome-Wide Identification and Evolutionary Analysis of Gossypium YTH Domain-Containing RNA-Binding Protein Family and the Role of GhYTH8 in Response to Drought Stress. Plants. 2023; 12(5):1198. https://doi.org/10.3390/plants12051198
Chicago/Turabian StyleHao, Wei, Weipeng Wang, Xiangfen Xiao, Jialiang Sun, Bingjie Wu, Yuping Zhao, Shuaishuai Pei, Wenjia Fan, Dongbei Xu, and Tengfei Qin. 2023. "Genome-Wide Identification and Evolutionary Analysis of Gossypium YTH Domain-Containing RNA-Binding Protein Family and the Role of GhYTH8 in Response to Drought Stress" Plants 12, no. 5: 1198. https://doi.org/10.3390/plants12051198