

Surfactin and Spo0A-Dependent Antagonism by Bacillus subtilis Strain UD1022 against Medicago sativa Phytopathogens
 , and
, and 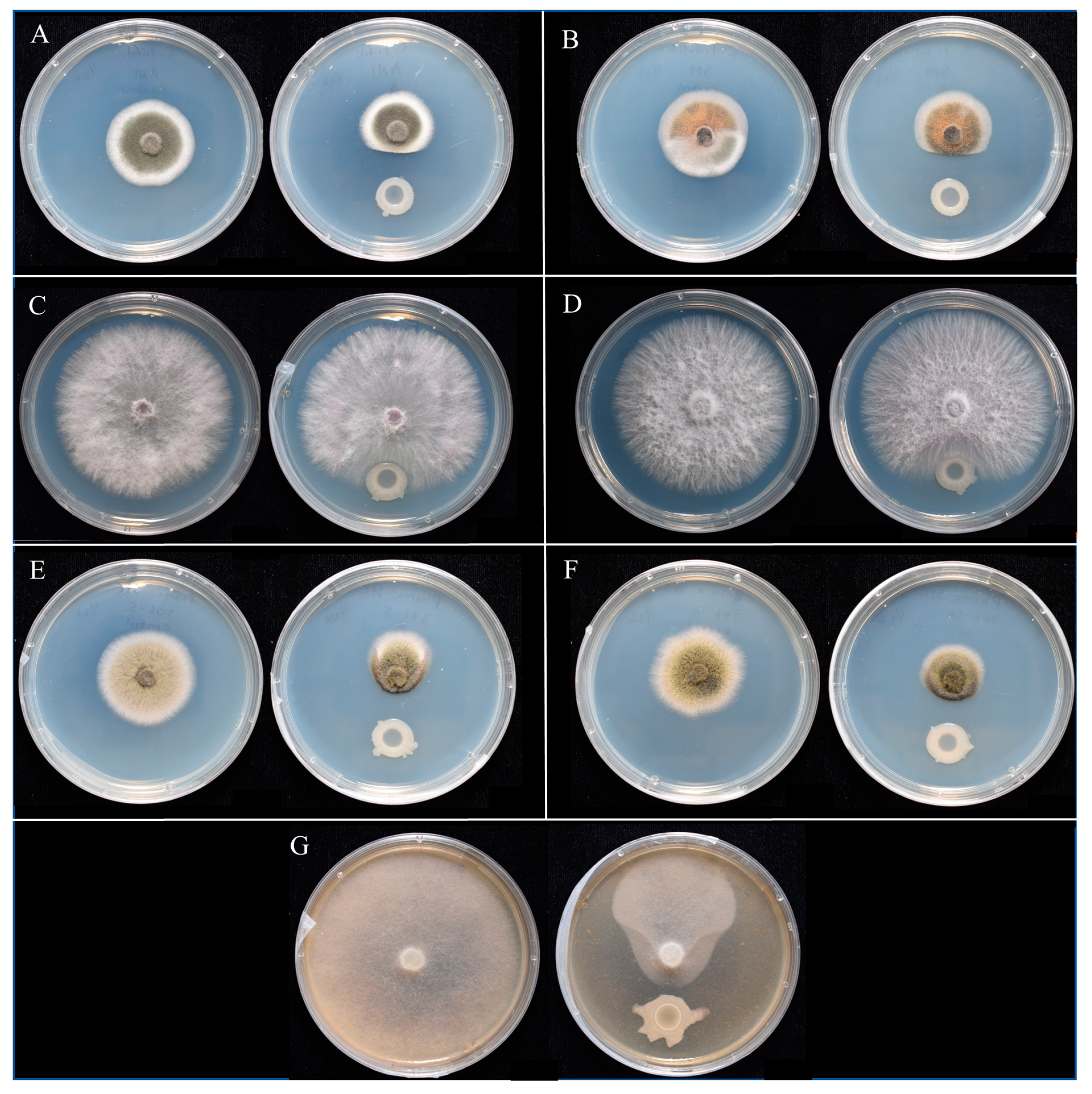{kind=link}
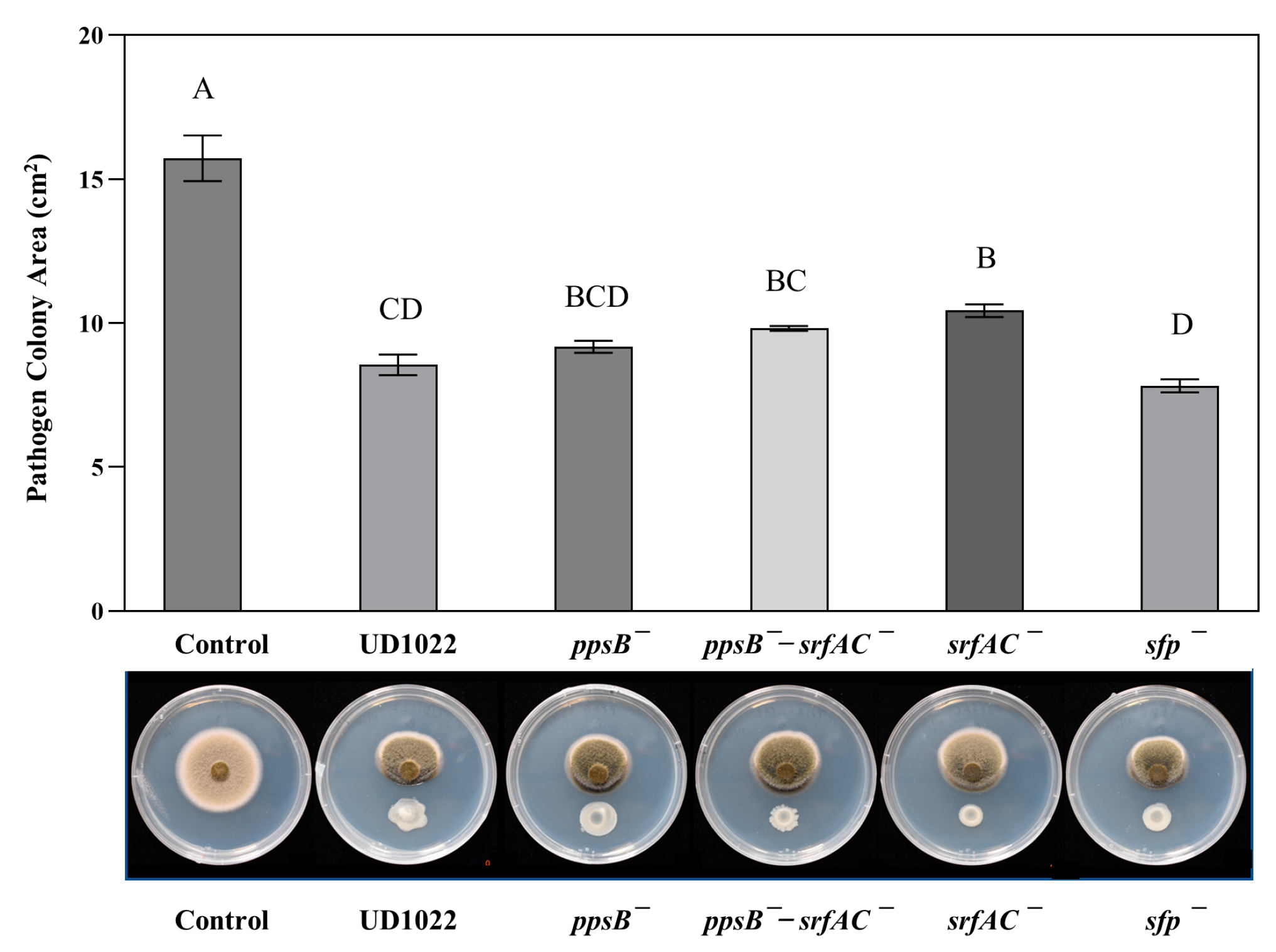{kind=link}
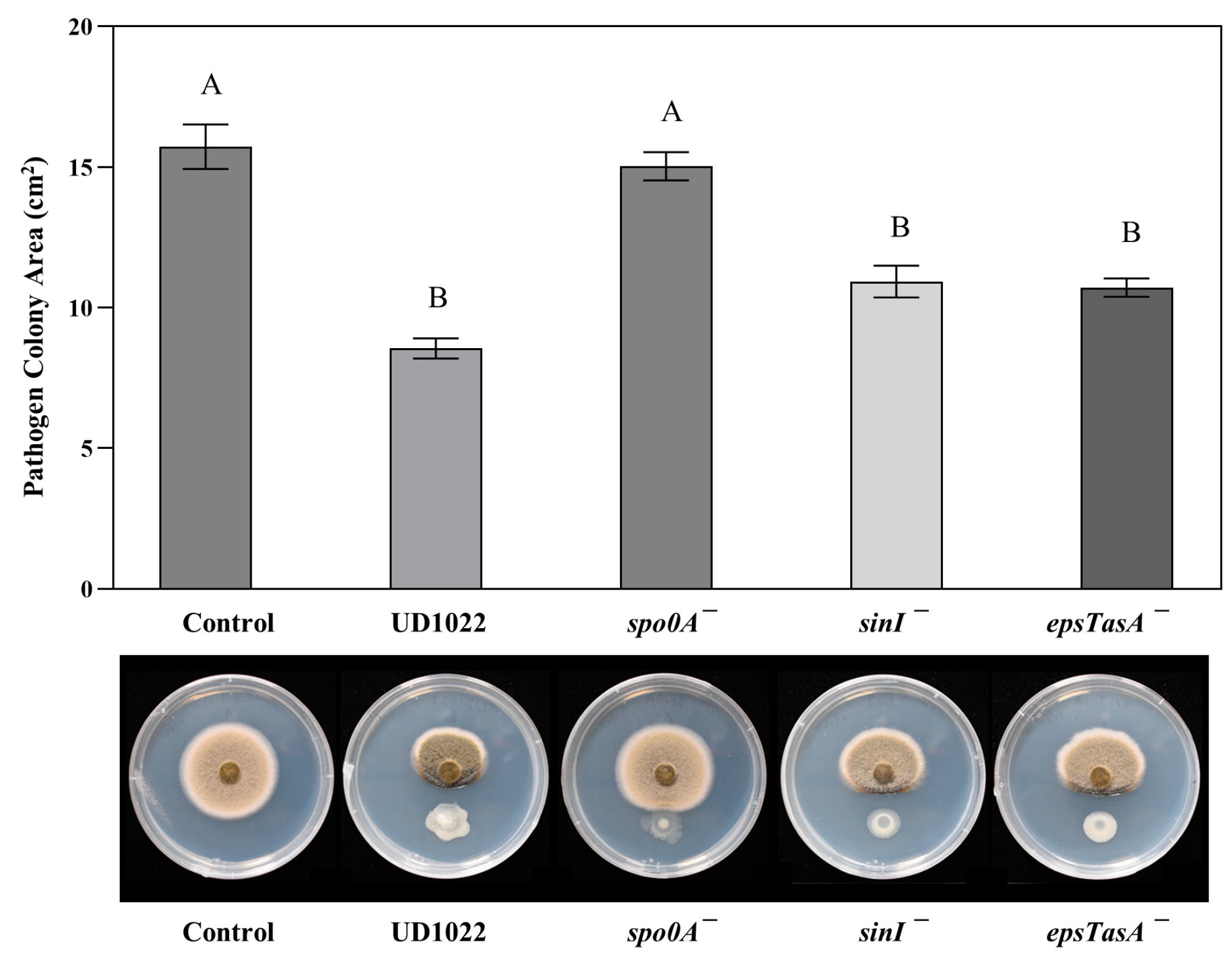{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Antagonistic Plate Assays
4.3. UD1022 Mutant Assays
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kulkarni, K.P.; Tayade, R.; Asekova, S.; Song, J.T.; Shannon, J.G.; Lee, J.-D. Harnessing the Potential of Forage Legumes, Alfalfa, Soybean, and Cowpea for Sustainable Agriculture and Global Food Security. Front. Plant. Sci. 2018, 9, 1314. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.R.; Samac, D.A. Biological Control of Fungi Causing Alfalfa Seedling Damping-Off with a Disease-Suppressive Strain of Streptomyces. Biol. Control 1996, 7, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Nutter, F.W.; Guan, J.; Gotlieb, A.R.; Rhodes, L.H.; Grau, C.R.; Sulc, R.M. Quantifying Alfalfa Yield Losses Caused by Foliar Diseases in Iowa, Ohio, Wisconsin, and Vermont. Plant Dis. 2002, 86, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Gao, M.; Xu, C.; Gao, J.; Deshpande, S.; Lin, S.; Roe, B.A.; Zhu, H. Alfalfa Benefits from Medicago Truncatula: The RCT1 Gene from M. truncatula Confers Broad-Spectrum Resistance to Anthracnose in Alfalfa. Proc. Natl. Acad. Sci. USA 2008, 105, 12164–12169. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhang, C.; Wang, Z.; Duan, T.; Yu, B.; Jia, X.; Pang, J.; Ma, L.; Wang, Y.; Nan, Z. Co-Infection by Soil-Borne Fungal Pathogens Alters Disease Responses among Diverse Alfalfa Varieties. Front. Microbiol. 2021, 12, 664385. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messéan, A. Toward a Reduced Reliance on Conventional Pesticides in European Agriculture. Plant Dis. 2016, 100, 10–24. [Google Scholar] [CrossRef] [Green Version]
- Thomashow, L.S.; Weller, D.M. Current Concepts in the Use of Introduced Bacteria for Biological Disease Control: Mechanisms and Antifungal Metabolites. In Plant-Microbe Interactions; Stacey, G., Keen, N.T., Eds.; Plant-Microbe Interactions; Springer: New York, NY, USA, 1996; pp. 187–235. ISBN 978-1-4612-8514-4. [Google Scholar]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Enhanced Plant Growth by Siderophores Produced by Plant Growth-Promoting Rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar] [CrossRef]
- Jacobsen, B.J.; Zidack, N.K.; Larson, B.J. The Role of Bacillus-Based Biological Control Agents in Integrated Pest Management Systems: Plant Diseases. Phytopathology 2004, 94, 1272–1275. [Google Scholar] [CrossRef] [Green Version]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus Species in Soil as a Natural Resource for Plant Health and Nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [Green Version]
- Pérez-García, A.; Romero, D.; De Vicente, A. Plant Protection and Growth Stimulation by Microorganisms: Biotechnological Applications of Bacilli in Agriculture. Curr. Opin. Biotechnol. 2011, 22, 187–193. [Google Scholar] [CrossRef]
- Handelsman, J.; Raffel, S.; Mester, E.H.; Wunderlich, L.; Grau, C.R. Biological Control of Damping-Off of Alfalfa Seedlings with Bacillus Cereus UW85. Appl. Environ. Microbiol. 1990, 56, 713–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Wu, H.-J.; Qiao, J.; Gao, X.; Borriss, R. Novel Routes for Improving Biocontrol Activity of Bacillus Based Bioinoculants. Front. Microbiol. 2015, 6, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural Functions of Lipopeptides from Bacillus and Pseudomonas: More than Surfactants and Antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrić, S.; Meyer, T.; Ongena, M. Bacillus Responses to Plant-Associated Fungal and Bacterial Communities. Front. Microbiol. 2020, 11, 1350. [Google Scholar] [CrossRef]
- Geissler, M.; Heravi, K.M.; Henkel, M.; Hausmann, R. Chapter 6—Lipopeptide Biosurfactants from Bacillus Species. In Biobased Surfactants, 2nd ed.; Hayes, D.G., Solaiman, D.K.Y., Ashby, R.D., Eds.; AOCS Press: Urbana, IL, USA, 2019; pp. 205–240. ISBN 978-0-12-812705-6. [Google Scholar]
- Quadri, L.E.N.; Weinreb, P.H.; Lei, M.; Nakano, M.M.; Zuber, P.; Walsh, C.T. Characterization of Sfp, a Bacillus subtilis Phosphopantetheinyl Transferase for Peptidyl Carrier Protein Domains in Peptide Synthetases. Biochemistry 1998, 37, 1585–1595. [Google Scholar] [CrossRef]
- Batool, M.; Khalid, M.H.; Hassan, M.N.; Fauzia Yusuf, H. Homology Modeling of an Antifungal Metabolite Plipastatin Synthase from the Bacillus subtilis 168. Bioinformation 2011, 7, 384–387. [Google Scholar] [CrossRef]
- Hamon, M.A.; Lazazzera, B.A. The Sporulation Transcription Factor Spo0A Is Required for Biofilm Development in Bacillus subtilis. Mol. Microbiol. 2001, 42, 1199–1209. [Google Scholar] [CrossRef]
- Nakano, M.M.; Marahiel, M.A.; Zuber, P. Identification of a Genetic Locus Required for Biosynthesis of the Lipopeptide Antibiotic Surfactin in Bacillus subtilis. J. Bacteriol. 1988, 170, 5662–5668. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.; Chu, F.; Kolter, R.; Losick, R. Bistability and Biofilm Formation in Bacillus subtilis. Mol. Microbiol. 2008, 67, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A Major Protein Component of the Bacillus subtilis Biofilm Matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef]
- Romero, D.; Aguilar, C.; Losick, R.; Kolter, R. Amyloid Fibers Provide Structural Integrity to Bacillus subtilis Biofilms. Proc. Natl. Acad. Sci. USA 2010, 107, 2230–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking Together: Building a Biofilm the Bacillus subtilis Way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against Infection of Arabidopsis Roots by Pseudomonas Syringae Is Facilitated by Biofilm Formation and Surfactin Production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yan, F.; Chai, Y.; Liu, H.; Kolter, R.; Losick, R.; Guo, J. Biocontrol of Tomato Wilt Disease by Bacillus subtilis Isolates from Natural Environments Depends on Conserved Genes Mediating Biofilm Formation. Environ. Microbiol. 2013, 15, 848–864. [Google Scholar] [CrossRef] [Green Version]
- Rahman, F.B.; Sarkar, B.; Moni, R.; Rahman, M.S. Molecular Genetics of Surfactin and Its Effects on Different Sub-Populations of Bacillus subtilis. Biotechnol. Rep. 2021, 32, e00686. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis Antibiotics: Structures, Syntheses and Specific Functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis Biofilm Induction by Plant Polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thérien, M.; Kiesewalter, H.T.; Auria, E.; Charron-Lamoureux, V.; Wibowo, M.; Maróti, G.; Kovács, Á.T.; Beauregard, P.B. Surfactin Production Is Not Essential for Pellicle and Root-Associated Biofilm Development of Bacillus subtilis. Biofilm 2020, 2, 100021. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.; Rudrappa, T. Methods for Promoting Plant Health 2013. Patent no. US12758361, 8 October 2013. Available online: https://patents.google.com/patent/US8551919B2/en (accessed on 27 June 2022).
- Bishnoi, U.; Polson, S.W.; Sherrier, D.J.; Bais, H.P. Draft Genome Sequence of a Natural Root Isolate, Bacillus subtilis UD1022, a Potential Plant Growth-Promoting Biocontrol Agent. Genome Announc. 2015, 3, e00696-15. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, V.; Kitto, S.L.; Caplan, J.L.; Hsueh, Y.-H.; Kearns, D.B.; Wu, Y.-S.; Bais, H.P. Microbe-Associated Molecular Patterns-Triggered Root Responses Mediate Beneficial Rhizobacterial Recruitment in Arabidopsis. Plant Physiol. 2012, 160, 1642–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloepper, J.W.; Schroth, M.N. Relationship of In Vitro Antibiosis of Plant Growth-Promoting Rhizobacteria to Plant Growth and the Displacement of Root Microflora. Phytopathology 1981, 71, 1020–1024. [Google Scholar] [CrossRef]
- Hu, J.; Zheng, M.; Dang, S.; Shi, M.; Zhang, J.; Li, Y. Biocontrol Potential of Bacillus Amyloliquefaciens LYZ69 Against Anthracnose of Alfalfa (Medicago sativa). Phytopathology 2021, 111, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Duan, T.; Christensen, M.J.; Nan, Z. Bacillus subtilis Subsp. Spizizenii MB29 Controls Alfalfa Root Rot Caused by Fusarium Semitectum. Biocontrol Sci. Technol. 2015, 25, 898–910. [Google Scholar] [CrossRef]
- Khan, N.; Maymon, M.; Hirsch, A.M. Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms 2017, 5, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boerema, G.H. Phoma Identification Manual: Differentiation of Specific and Infra-Specific Taxa in Culture; CABI: Wallingford, UK, 2004; ISBN 0-85199-743-0. [Google Scholar]
- Castell-Miller, C.V.; Zeyen, R.J.; Samac, D.A. Infection and Development of Phoma Medicaginis on Moderately Resistant and Susceptible Alfalfa Genotypes. Can. J. Plant Pathol. 2007, 29, 290–298. [Google Scholar] [CrossRef]
- Coles, D.W.; Bithell, S.L.; Mikhael, M.; Cuddy, W.S.; Plett, J.M. Chickpea Roots Undergoing Colonisation by Phytophthora Medicaginis Exhibit Opposing Jasmonic Acid and Salicylic Acid Accumulation and Signalling Profiles to Leaf Hemibiotrophic Models. Microorganisms 2022, 10, 343. [Google Scholar] [CrossRef]
- Vandemark, G.J.; Ariss, J.J.; Hughes, T.J. Real-Time PCR Suggests That Aphanomyces Euteiches Is Associated with Reduced Amounts of Phytophthora Medicaginis in Alfalfa That Is Co-Inoculated with Both Pathogens. J. Phytopathol. 2010, 158, 117–124. [Google Scholar] [CrossRef]
- Xiao, K.; Kinkel, L.L.; Samac, D.A. Biological Control of Phytophthora Root Rots on Alfalfa and Soybean with Streptomyces. Biol. Control 2002, 23, 285–295. [Google Scholar] [CrossRef]
- Kiesewalter, H.T.; Lozano-Andrade, C.N.; Wibowo, M.; Strube, M.L.; Maróti, G.; Snyder, D.; Jørgensen, T.S.; Larsen, T.O.; Cooper, V.S.; Weber, T.; et al. Genomic and Chemical Diversity of Bacillus subtilis Secondary Metabolites against Plant Pathogenic Fungi. mSystems 2021, 6, e00770-20. [Google Scholar] [CrossRef]
- Sun, J.; Liu, Y.; Lin, F.; Lu, Z.; Lu, Y. CodY, ComA, DegU and Spo0A Controlling Lipopeptides Biosynthesis in Bacillus Amyloliquefaciens FmbJ. J. Appl. Microbiol. 2021, 131, 1289–1304. [Google Scholar] [CrossRef]
- Wang, M.; Yu, H.; Li, X.; Shen, Z. Single-Gene Regulated Non-Spore-Forming Bacillus subtilis: Construction, Transcriptome Responses, and Applications for Producing Enzymes and Surfactin. Metab. Eng. 2020, 62, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Liu, Y.; Xie, J.; Štefanič, P.; Lv, Y.; Fan, B.; Mandic-Mulec, I.; Zhang, R.; Shen, Q.; Xu, Z. Annulment of Bacterial Antagonism Improves Plant Beneficial Activity of a Bacillus Velezensis Consortium. Appl. Environ. Microbiol. 2022, 88, e0024022. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, C.; Liang, J.; Wu, L.; Gao, W.; Jiang, J. Iturin A Extracted from Bacillus subtilis WL-2 Affects Phytophthora Infestans via Cell Structure Disruption, Oxidative Stress, and Energy Supply Dysfunction. Front. Microbiol. 2020, 11, 536083. [Google Scholar] [CrossRef]
- Hölscher, T.; Kovács, Á.T. Sliding on the Surface: Bacterial Spreading without an Active Motor. Environ. Microbiol. 2017, 19, 2537–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gestel, J.; Vlamakis, H.; Kolter, R. From Cell Differentiation to Cell Collectives: Bacillus subtilis Uses Division of Labor to Migrate. PLoS Biol. 2015, 13, e1002141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grau, R.R.; De Oña, P.; Kunert, M.; Leñini, C.; Gallegos-Monterrosa, R.; Mhatre, E.; Vileta, D.; Donato, V.; Hölscher, T.; Boland, W.; et al. A Duo of Potassium-Responsive Histidine Kinases Govern the Multicellular Destiny of Bacillus subtilis. mBio 2015, 6, e00581. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chen, Z.; Yu, L.; Hu, D.; Song, B. Investigating the Antifungal Activity and Mechanism of a Microbial Pesticide Shenqinmycin against Phoma Sp. Pestic. Biochem. Physiol. 2018, 147, 46–50. [Google Scholar] [CrossRef]
- Nihorimbere, V.; Cawoy, H.; Seyer, A.; Brunelle, A.; Thonart, P.; Ongena, M. Impact of Rhizosphere Factors on Cyclic Lipopeptide Signature from the Plant Beneficial Strain Bacillus Amyloliquefaciens S499. FEMS Microbiol. Ecol. 2012, 79, 176–191. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Yasbin, R.E.; Young, F.E. Transduction in Bacillus subtilis by Bacteriophage SPP1. J. Virol. 1974, 14, 1343–1348. [Google Scholar] [CrossRef] [Green Version]
- Stragier, P.; Losick, R. Molecular Genetics of Sporulation in Bacillus subtilis. Annu. Rev. Genet. 1996, 30, 297–341. [Google Scholar] [CrossRef] [PubMed]
- Ben Slimene, I.; Tabbene, O.; Djebali, N.; Cosette, P.; Schmitter, J.M.; Jouenne, T.; Urdaci, M.-C.; Limam, F. Putative Use of a Bacillus subtilis L194 Strain for Biocontrol of Phoma Medicaginis in Medicago Truncatula Seedlings. Res. Microbiol. 2012, 163, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tao, J.; Yan, Y.; Li, B.; Li, H.; Li, C. Biocontrol Efficiency of Bacillus subtilis SL-13 and Characterization of an Antifungal Chitinase. Chin. J. Chem. Eng. 2011, 19, 128–134. [Google Scholar] [CrossRef]
- Dimkić, I.; Stanković, S.; Nišavić, M.; Petković, M.; Ristivojević, P.; Fira, D.; Berić, T. The Profile and Antimicrobial Activity of Bacillus Lipopeptide Extracts of Five Potential Biocontrol Strains. Front. Microbiol. 2017, 8, 925. [Google Scholar] [CrossRef] [Green Version]
- López, D.; Kolter, R. Extracellular Signals That Define Distinct and Coexisting Cell Fates in Bacillus subtilis. FEMS Microbiol. Rev. 2010, 34, 134–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spacapan, M.; Danevčič, T.; Mandic-Mulec, I. ComX-Induced Exoproteases Degrade ComX in Bacillus subtilis PS-216. Front. Microbiol. 2018, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Cawoy, H.; Debois, D.; Franzil, L.; De Pauw, E.; Thonart, P.; Ongena, M. Lipopeptides as Main Ingredients for Inhibition of Fungal Phytopathogens by Bacillus subtilis/Amyloliquefaciens. Microb. Biotechnol. 2015, 8, 281–295. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosier, A.; Pomerleau, M.; Beauregard, P.B.; Samac, D.A.; Bais, H.P. Surfactin and Spo0A-Dependent Antagonism by Bacillus subtilis Strain UD1022 against Medicago sativa Phytopathogens. Plants 2023, 12, 1007. https://doi.org/10.3390/plants12051007
Rosier A, Pomerleau M, Beauregard PB, Samac DA, Bais HP. Surfactin and Spo0A-Dependent Antagonism by Bacillus subtilis Strain UD1022 against Medicago sativa Phytopathogens. Plants. 2023; 12(5):1007. https://doi.org/10.3390/plants12051007
Chicago/Turabian StyleRosier, Amanda, Maude Pomerleau, Pascale B. Beauregard, Deborah A. Samac, and Harsh P. Bais. 2023. "Surfactin and Spo0A-Dependent Antagonism by Bacillus subtilis Strain UD1022 against Medicago sativa Phytopathogens" Plants 12, no. 5: 1007. https://doi.org/10.3390/plants12051007