
Genomic C-Value Variation Analysis in Jujube (Ziziphus jujuba Mill.) in the Middle Yellow River Basin
Abstract
:1. Introduction
2. Results
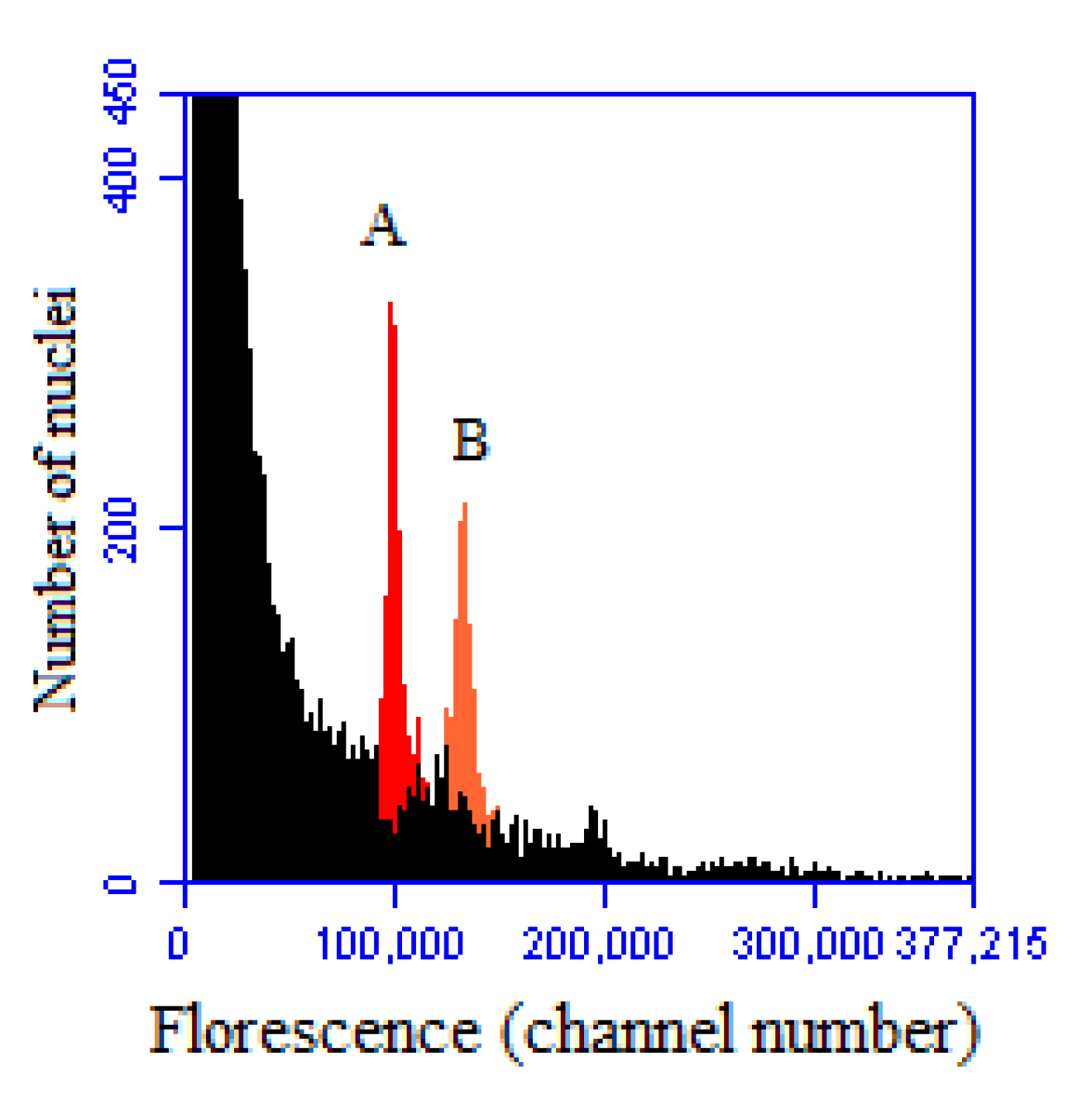
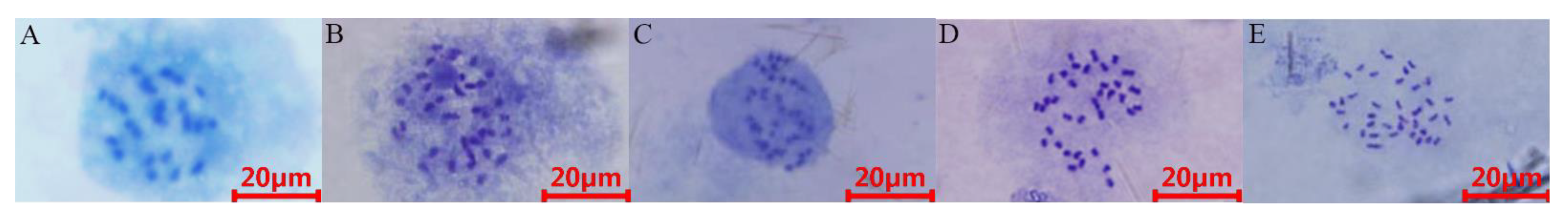
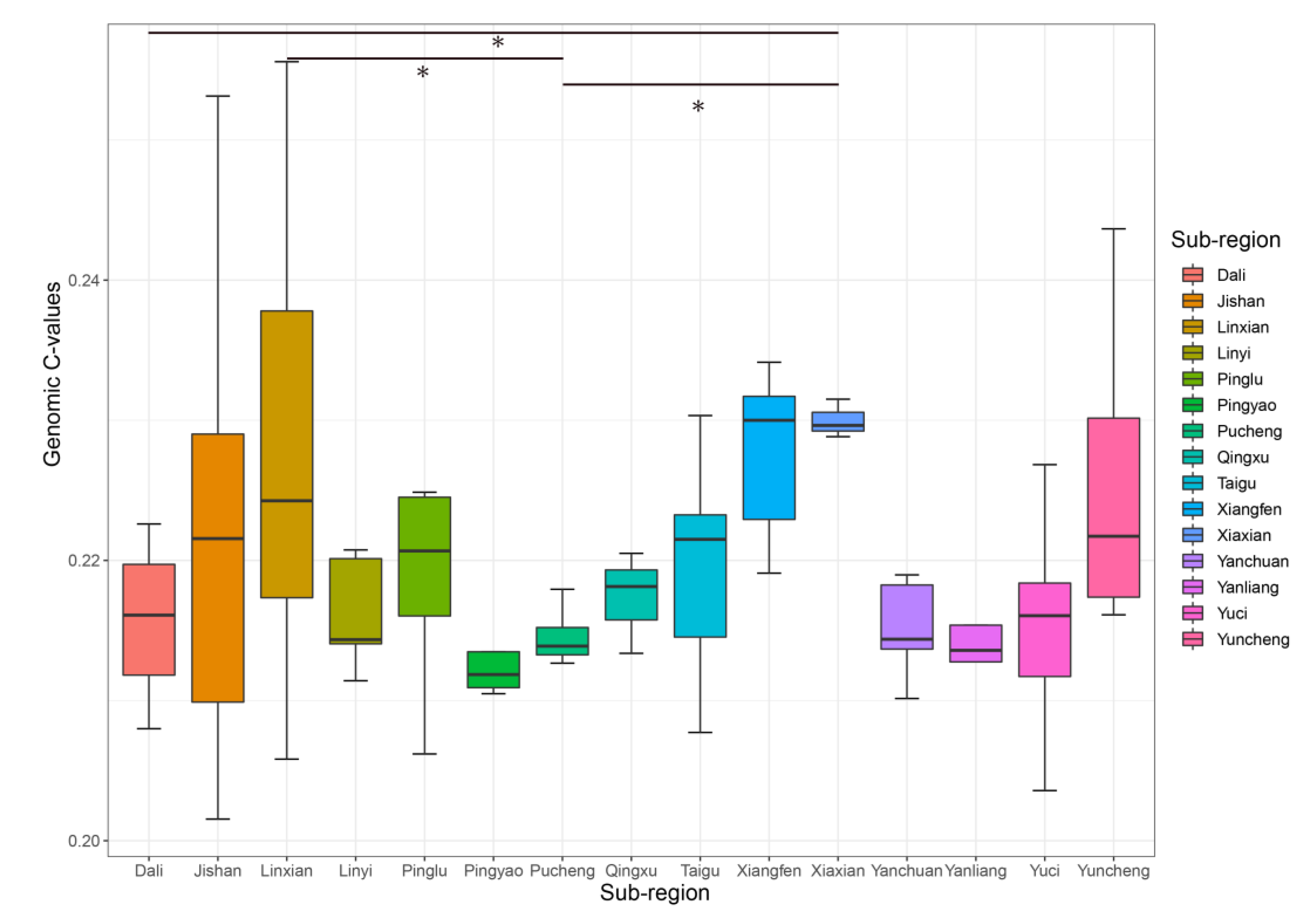
2.1. Variability of Genomic C-Value and Ploidy Analysis
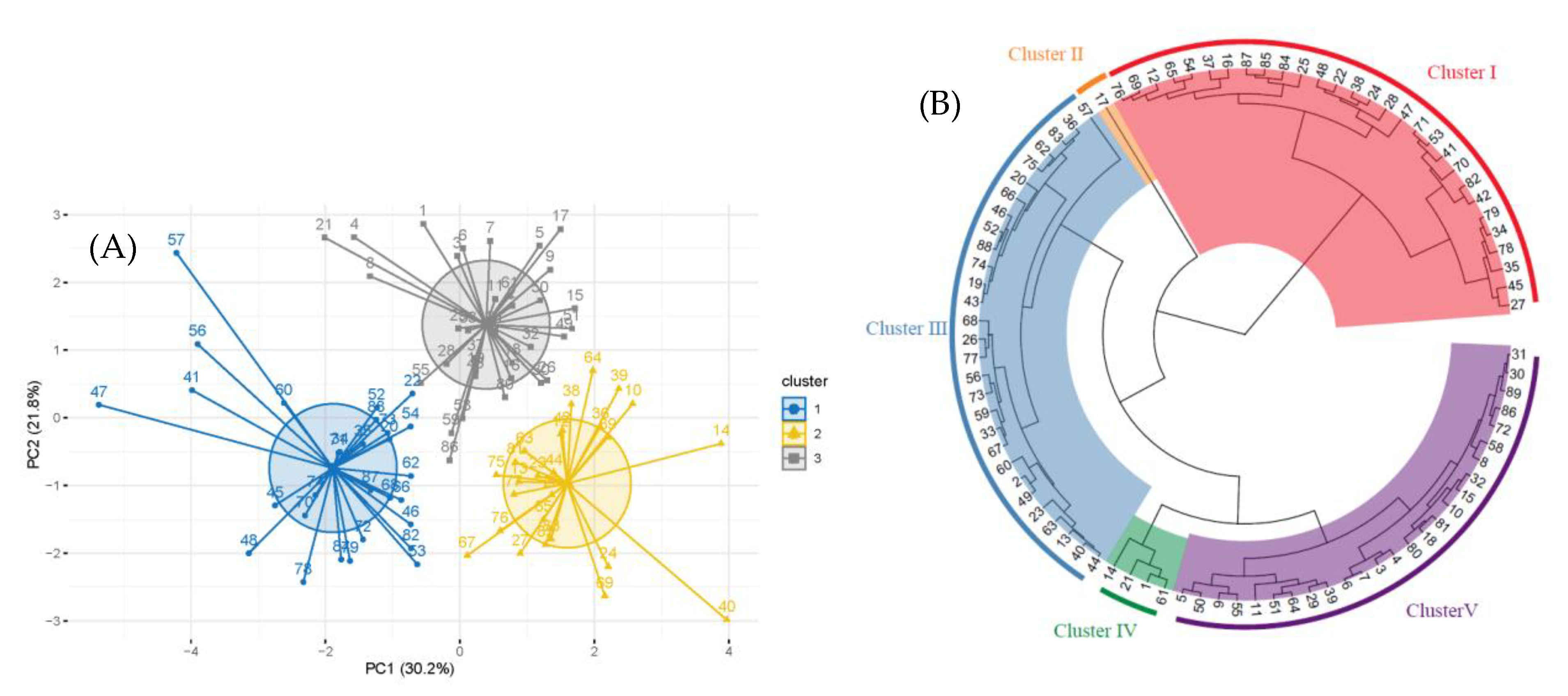
2.2. Principal Component Analysis and Cluster Analysis of Genome Size and Phenotypic Traits in Jujube Germplasm
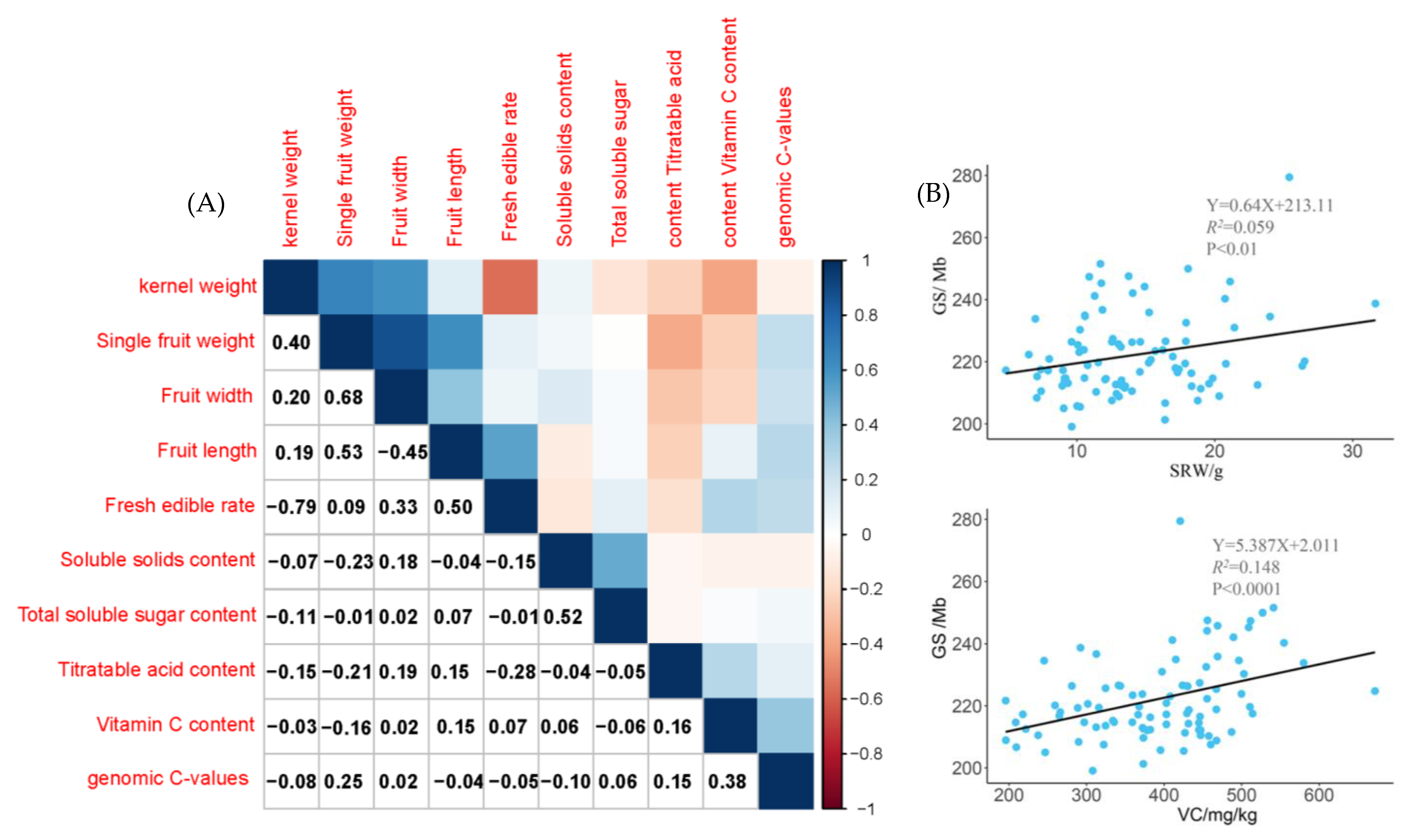
2.3. Correlation Analysis between Genomic C-values and Agronomic Traits of Jujube Germplasm
3. Discussion
3.1. Genomic C-Values and Variation of Jujube Germplasm
3.2. Ploidy of Jujube Germplasm
3.3. Correlation between Genomic C-Values and Fruit Traits of Jujube
4. Materials and Methods
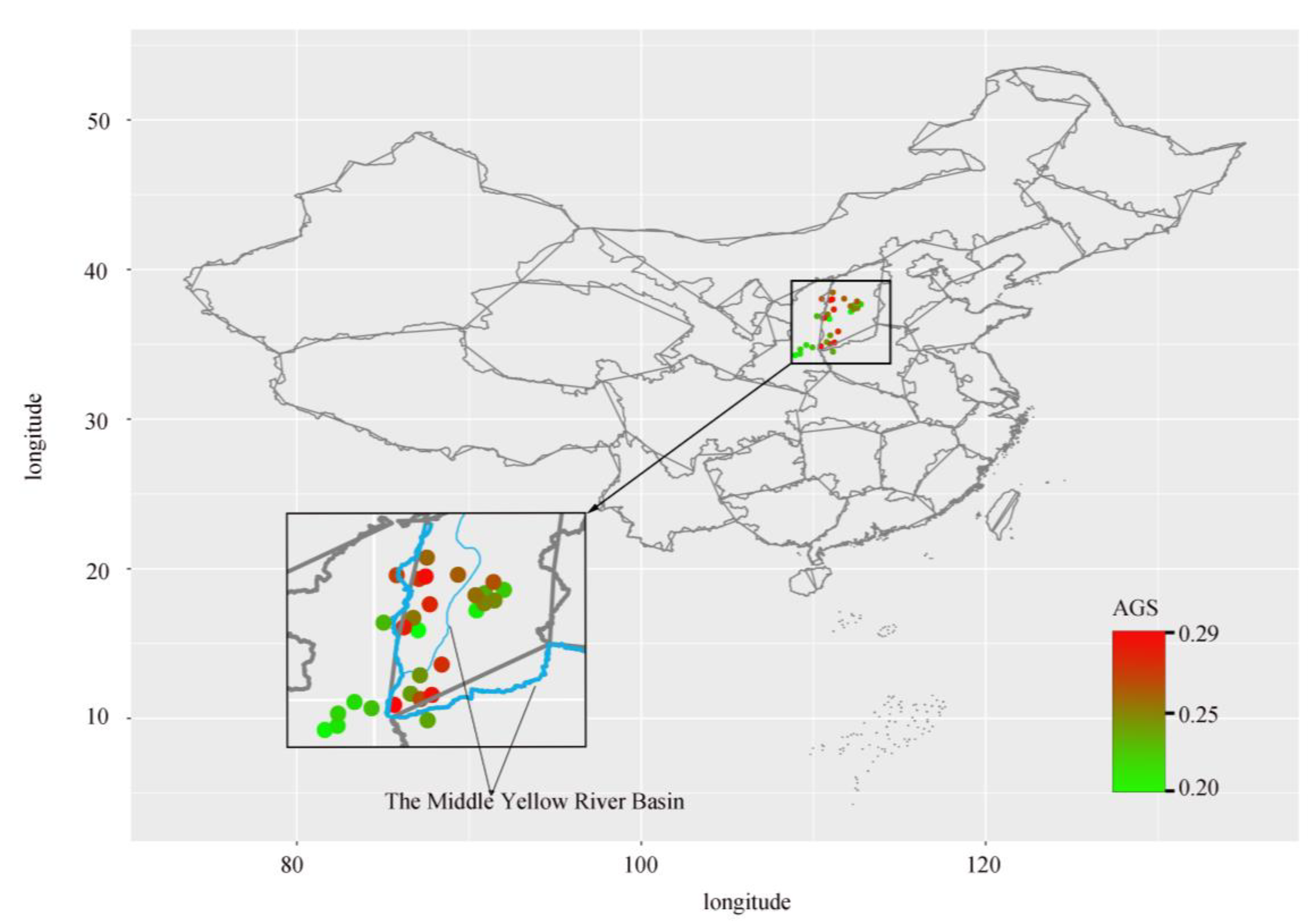
4.1. Plant Material
4.2. Identification and Evaluation of Important Fruit Traits
4.3. Genome Size Assessment
4.4. Chromosome Count
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, M.J.; Wang, J.R.; Wang, L.L.; Wang, P.; Liu, J.; Zhao, Z.H.; Yao, S.R.; Florin, S.; Wang, L.X.; Ao, C.W.; et al. The historical and current research progress on jujube-a superfruit for the future. Hortic. Res. 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y. Genetic Diversity and Population Structure of Chinese Jujube Analysed by SSR Markers. Ph.D. Thesis, Northwest A & F University, Yang Ling, China, 2013. [Google Scholar]
- Qu, Z.; Wang, Y. Chinese Fruit Trees Record-Chinese Jujube; China Forestry Publishing House: Beijing, China, 1993; pp. 10–60. [Google Scholar]
- Liu, M.J.; Wang, M. Germplasm Resources of Chinese Jujube; China Forestry Publishing House: Beijing, China, 2009; pp. 24–65. [Google Scholar]
- Greilhuber, J.; Doležel, J.; Lysák, M.A.; Bennett, M.D. The Origin, Evolution and Proposed Stabilization of the Terms ‘Genome Size’ and ‘C-Value’ to Describe Nuclear DNA Contents. Ann. Bot. 2005, 95, 255–260. [Google Scholar] [CrossRef]
- Xu, P.F.; Yang, Y.H.; Zhang, Y.T.; Chen, Y.; Tang, D.Q. Induction and Preliminary Identification of Tetraploid in Phyllostachys edulis. Sci. Silvae Sin. 2020, 56, 55–62. [Google Scholar]
- Cakovic, D.; Cresti, L.; Steševic, D.; Schönswetter, P.; Frajman, B. High Genetic and Morphological Diversification of the Euphorbia verrucosa Alliance (Euphorbiaceae) in the Balkan and Iberian Peninsulas. Taxon 2021, 70, 286–307. [Google Scholar] [CrossRef]
- Suda, J.; Krahulcová, A.; Trávnícek, P.; Rosenbaumová, R.; Peckert, T.; Krahulec, F. Genome Size Variation and Species Relationships in Hieracium Sub-Genus Pilosella (Asteraceae) as Inferred by Flow Cytometry. Ann. Bot. 2007, 100, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Šmarda, P.; Bureš, P.; Horová, L.; Rotreklová, O. Intrapopulation Genome Size Dynamics in Festuca pallens. Ann. Bot. 2008, 102, 599–607. [Google Scholar] [CrossRef]
- Slovák, M.; Vít, P.; Urfus, T.; Suda, J. Complex Pattern of Genome Size Variation in a Polymorphic Member of the Asteraceae. J Biogeogr 2009, 36, 372–384. [Google Scholar] [CrossRef]
- Ladner, J.; Mayfield, M.H.; Prather, L.A.; Ferguson, C.J. Polyploidy and Genome Size Variation in Phlox nana (Polemoniaceae) from the Pecos Plains of New Mexico and the Davis Mountains of West Texas, USA. J. Bot. Res. Inst. Texas 2017, 11, 351–362. [Google Scholar] [CrossRef]
- Pellicer, J.; Hidalgo, O.; Dodsworth, S.; Leitch, I. Genome Size Diversity and Its Impact on the Evolution of Land Plants. Genes 2018, 9, 88. [Google Scholar] [CrossRef]
- Hodálová, I.; Mártonfiová, L.; Skokanová, K.; Majerová, M.; Somlyay, L.; Mered’a, P. The Utility of Genome Size in Plant Identification: A Case Study on Sesleria (Poaceae) from Croatia and Slovenia. Plant Syst. Evol. 2020, 306, 87. [Google Scholar] [CrossRef]
- Fay, M.F.; Conwan, R.S.; Leitch, L.J. The effects of nuclear DNA content (C-value) on the quality and utility of AFLP fingerprints. Ann. Bot. 2005, 95, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rewers, M.; Jedrzejczyk, I.; Rewicz, A.; Busse, A.J. Genome Size Diversity in Rare, Endangered, and Protected Orchids in Poland. Genes 2021, 12, 563. [Google Scholar] [CrossRef]
- Aliyu, O.M. Analysis of absolute nuclear DNA content reveals a small genome and intra-specific variation in Cashew (Anacardium occidentale L.), Anacardiaceae. Silvae Genet. 2014, 63, 285–293. [Google Scholar] [CrossRef]
- Comertpay, G. Assessment of Nuclear DNA Contents Variation and Their Relationship with Flowering in Corn Genotypes. Turk J. Field Crops 2019, 24, 39–45. [Google Scholar] [CrossRef]
- Meagher, T.R.; Christine, V. Phenotypic impacts of repetitive DNA in flowering plants. New Phytol. 2005, 168, 71–80. [Google Scholar] [CrossRef]
- Carta, A.; Mattana, E.; Dickie, J.; Vandelook, F. Correlated evolution of seed mass and genome size varies among life forms in flowering plants. Seed Sci. Res. 2022, 32, 46–52. [Google Scholar] [CrossRef]
- Korban, S.S.; Wannarat, W.; Rayburn, C.M.; Tatum, T.C.; Rayburn, A.L. Genome size and nucleotypic variation in Malus germplasm. Genome 2009, 52, 148–155. [Google Scholar] [CrossRef]
- Liu, M.J.; Zhao, J.; Cai, Q.L.; Liu, G.C.; Wang, J.R.; Zhao, Z.H.; Liu, P.; Dai, L.; Yan, G.J.; Wang, W.J.; et al. The complex jujube genome provides insights into fruit tree biology. Nat. Commun. 2014, 5, 5315. [Google Scholar] [CrossRef]
- Dong, X.G.; Wang, Z.; Tian, L.M.; Zhang, Y.; Qi, D.; Huo, H.L.; Xu, J.Y.; Li, Z.; Liao, R.; Shi, M.; et al. De novo assembly of a wild pear (Pyrus betuleafolia) genome. Plant Biotechnol. J. 2020, 18, 581–595. [Google Scholar] [CrossRef]
- Trávnícek, P.; Certner, M.; Ponert, J.; Chumová, Z.; Jersáková, J.; Suda, J. Diversity in Genome Size and GC Content Shows Adaptive Potential in Orchids and is Closely Linked to Partial Endoreplication, Plant Life-History Traits and Climatic Conditions. New Phytol. 2019, 224, 1642–1656. [Google Scholar] [CrossRef]
- Kraaijeveld, K. Genome Size and Species Diversification. Evol. Biol. 2010, 37, 227–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.K.; Yan, X.; Feng, Y.Y.; Ran, J.H.; Qian, Z.J.; Yan, X.Y.; Zhou, S.S.; Fang, T.Z.; Ma, X.F. Genome size variations and species differentiation of Reaumuria soongarica. Biodiv. Sci. 2021, 29, 1308–1320. [Google Scholar] [CrossRef]
- Piegu, B.; Guyot, R.; Picault, N.; Roulin, A.; Saniyal, A.; Kim, H.; Collura, K.; Brar, D.S.; Jackson, S.; Wing, R.A.; et al. Doubling Genome Size without Polyploidization: Dynamics of Retrotransposition-Driven Genomic Expansions in Oryza australiensis, a Wild Relative of Rice. Genome Res. 2006, 16, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Šmarda, P.; Bureš, P. Intraspecific DNA Content Variability in Festuca pallens on Different Geographical Scales and Ploidy Levels. Ann. Bot. 2006, 98, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, K.X.; Kong, G.H.; Yi, S.B. Genome Size and Variation Analysis of Mango (Mangifera indica L.) Germplasms in Yunnan by Flow Cytometry. J. Trop. Subtrop. Bot. 2015, 23, 386–390. [Google Scholar]
- Quan, L.; Rabanal, F.A.; Meng, D.Z.; Huber, C.D.; Farlow, A.; Pltzer, A.; Zhang, Q.R.; Vilhjmsson, T.; Lysak, M.A.; Seren, U.; et al. Massive genomic variation and strong selection in Arabidopsis thaliana lines from Sweden. Nat. Genet. 2013, 45, 884–890. [Google Scholar]
- Noirot, M.; Poncet, V.; Barre, P.; Hamon, S.; DE, K.A. Genome Size Variations in Diploid African Coffea Species. Ann. Bot. 2003, 92, 709–714. [Google Scholar] [CrossRef]
- Nowicka, A.; Sliwinsk, E.; Grzebelus, D.; Baranski, R.; Simon, P.W.; Nothnagel, T.; Grzebelus, E. Nuclear DNA content variation within the genus Daucus (Apiaceae) determined by flow cytometry. Sci. Hortic. 2016, 209, 132–138. [Google Scholar] [CrossRef]
- Redpath, L.E.; Aryal, R.; Lynch, N.; Spencer, J.A.; Hulse-kemp, A.M.; Ballington, J.R.; Green, J.; Bassil, N.; Hummer, K.; Ranney, T.; et al. Nuclear DNA contents and ploidy levels of North American Vaccinium species and interspecific hybrids. Sci. Hortic. 2022, 297, 110955. [Google Scholar] [CrossRef]
- Bureš, P.; Wang, Y.-F.; Horova, L.; Suda, J. Genome Size Variation in Central European Species of Cirsium (Compositae) and Their Natural Hybrids. Ann. Bot. 2004, 94, 353–363. [Google Scholar] [CrossRef]
- Bennett, M.D.; Leitch, I.J. Genome Size Evolution in Plants. In The Evolution of the Genome; Gregory, T.R., Ed.; Academic Press: Burlington, MA, USA, 2005; pp. 89–162. [Google Scholar]
- Vidic, T.; Greilhuber, J.; Vilhar, B.; Dermastia, M. Selective Significance of Genome Size in a Plant Community with Heavy Metal Pollution. Ecol. Appl. 2009, 19, 1515–1521. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Wang, J.; Huang, H. Nitrogen Limitation as a Driver of Genome Size Evolution in a Group of Karst Plants. Sci. Rep. 2015, 5, 11636. [Google Scholar] [CrossRef]
- Bilinski, P.; Albert, P.S.; Berg, J.J.; Birchler, J.A.; Grote, M.N.; Lorant, A.; Quezada, J.; Swarts, K.; Yang, J.; Ross-Ibarra, J. Parallel Altitudinal Clines Reveal Trends in Adaptive Evolution of Genome Size in Zea mays. PLoS Genet. 2018, 14, e1007162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kron, P.; Suda, J.; Husband, B.C. Applications of Flow Cytometry to Evolutionary and Population Biology. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 847–876. [Google Scholar] [CrossRef]
- Slynko, N.M.; Burmakina, N.V.; Potseluyev, O.M.; Kapustyanchik, S.Y.; Galitsin, G.Y.; Goryachkovskaya, T.N.; Kuybida, L.V.; Shekhovtsov, S.V.; Peltek, S.E.; Shumny, V.K. Erratum to: “Gas chromatography-mass spectrometry in the taxonomy of Miscanthus”. Vavilovskii Zhurnal Genet. Sel. 2019, 23, 1076–1081. [Google Scholar] [CrossRef] [PubMed]
- Bennett, S.J.; Hayward, M.; Marshall, D. Morphological differentiation in four species of the genus Lolium. Genet. Resour. Crop Evol. 2000, 47, 247–255. [Google Scholar] [CrossRef]
- Landis, J.B.; Soltis, D.E.; Li, Z.; Marx, H.E.; Barker, M.S.; Tank, D.C.; Soltis, P.S. Impact of whole-genome duplication events on diversification rates in angiosperms. Am. J. Bot. 2018, 105, 348–363. [Google Scholar] [CrossRef]
- Hawkins, J.S.; Hu, G.J.; Rapp, R.A.; Grafenberg, J.L.; Wendel, J.F. Phylogenetic determination of the pace of transposable element proliferation in plants: Copia and LINE-like elements in Gossypium. Genome 2008, 51, 11–18. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, Z.; Li, S.; Lian, Q.; Fu, P.; He, Y.; Qiao, J.; Xu, K.; Liu, L.; Wu, M.; et al. Genomic analyses of diverse wild and cultivated accessions provide insights into the evolutionary history of jujube. Plant Biotechnol. J. 2020, 19, 517–531. [Google Scholar] [CrossRef]
- Liu, M.J. Chinese jujube: Botany and horticulture. Hortic. Rev. 2006, 32, 229–298. [Google Scholar]
- Wang, L.H.; Luo, Z.; Liu, Z.G.; Zhao, J.; Deng, W.P.; Wei, H.R.; Liu, P.; Liu, M.J. Genome Size Variation within Species of Chinese Jujube (Ziziphus jujuba Mill.) and Its Wild Ancestor Sour Jujube (Z. acidojujuba Cheng et Liu). Forests 2019, 10, 460. [Google Scholar] [CrossRef]
- Kadkhodaei, S.; Arzani, K.; Yadollahi, A.; Karimzadeh, G.; Abdollahi, H. Genetic Diversity and Similarity of Asian and European Pears (Pyrus Spp.) Revealed by Genome Size and Morphological Traits Prediction. Int. J. Fruit Sci. 2021, 21, 619–633. [Google Scholar] [CrossRef]
- Sarikhani, S.; Arzani, K.; Karimzadeh, G.; Shojaeiyan, A.; Ligterink, W. Genome Size: A Novel Predictor of Nut Weight and Nut Size of Walnut Trees. HortScience 2018, 53, 275–282. [Google Scholar]
- Tiley, G.P.; Burleigh, J.G. The relationship of recombination rate, genome structure, and patterns of molecular evolution across angiosperms. BMC Evol. Biol. 2015, 15, 194. [Google Scholar] [CrossRef]
- Domínguez, M.; Dugas, E.; Benchouaia, M.; Leduque, B.; Jiménez-Gómez, J.M.; Colot, V.; Quadrana, L. The impact of transposable elements on tomato diversity. Nat. Commun. 2020, 11, 4058. [Google Scholar] [CrossRef]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons control fruit-specific, cold-dependent accumulation of anthocyanins in blood oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef]
- Chen, J.; Lu, L.; Benjamin, J.; Diaz, S.; Hancock, C.N.; Stajich, J.E.; Wessler, S.R. Tracking the origin of two genetic components associated with transposable element bursts in domesticated rice. Nat. Commun. 2019, 10, 641. [Google Scholar] [CrossRef]
- Wang, L.H.; Lv, Y.; Luo, Z.; Liu, P.; Liu, M.J. Establishment and Application of a Method for Chromosome Ploidy Identification and Genome Size Estimation Using Flow Cytometry in Ziziphus jujuba. Chin. J. Agric. Biotechnol. 2018, 26, 511–520. [Google Scholar]
- Loureiro, J.; Rodriguez, E.; Dolezel, J.; Santos, C. Two New Nuclear Isolation Buffers for Plant DNA Flow Cytometry: A Test with 37 Species. Ann. Bot. 2007, 100, 875–888. [Google Scholar] [CrossRef]
- Kuo, L.Y.; Tang, S.K.; Kao, T.T.; Ebihara, A.; Fawcett, S.; Hsiao, M.C.; Shinohara, W.; Dauphin, B. A dormant resource for genome size estimation in ferns: C-value inference of the Ophioglossaceae using herbarium specimen spores. Appl. Plant Sci. 2021, 9, e11452. [Google Scholar]
- Chen, R.Y.; Song, W.Q.; Li, X.L. Wall Degradation Hypotonic Method of Preparing Chromosome Samples in Plant and Its Significance in the Cytogenetics. Acta Genet. Sin. 1982, 9, 151–159. [Google Scholar]
- Dolezel, J.; Bartos, J.; Voglmayr, H.; Greilhuber, J. Nuclear DNA content and genome size of trout and human. Cytometry A 2003, 51, 127–128. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Germplasm | Mean(pg) ± SD | LSD Group | No. | Germplasm | Mean ± SD (pg) | LSD Group |
|---|---|---|---|---|---|---|---|
| 1 | LXMJW-1 | 0.25559 ± 0.00455 | a | 27 | LXWWZ | 0.21163 ± 0.01031 | nop |
| 2 | LXMJW-2 | 0.24662 ± 0.00335 | abcde | 28 | LXYLLZ | 0.20582 ± 0.00153 | p |
| 3 | LXMJW-3 | 0.24119 ± 0.0097 | abcdefg | 29 | LXLJPAAZ | 0.24571 ± 0.00771 | abcdef |
| 4 | LXMJW-4 | 0.25135 ± 0.00137 | ab | 30 | LXHGZ | 0.21552 ± 0.00298 | klmnop |
| 5 | LXXZZ-2 | 0.24856 ± 0.0085 | abcd | 31 | LXGJJZ | 0.23797 ± 0.00104 | abcdefgh |
| 6 | LXXZZ-1 | 0.25082 ± 0.00826 | ab | 32 | LXXCZ | 0.23099 ± 0.00449 | defghijkl |
| 7 | LXWJZDZ | 0.24969 ± 0.01293 | abc | 33 | LXQJSTZ | 0.21403 ± 0.00069 | lmnop |
| 8 | LXYJZ-1 | 0.25313 ± 0.00008 | ab | 34 | LXYJMYZ | 0.23151 ± 0.00345 | cdefghijkl |
| 9 | LXYJZ-2 | 0.2378 ± 0.00069 | abcdefghi | 35 | LXLJZ | 0.21962 ± 0.00232 | ijklmnop |
| 10 | LXYJZ-3 | 0.23711 ± 0.00918 | bcdefghij | 36 | LXQJSLJZ | 0.21703 ± 0.00684 | klmnop |
| 11 | LXTHDMZ-1 | 0.24027 ± 0.00215 | abcdefgh | 37 | LXQJSLJZ | 0.21935 ± 0.00071 | jklmnop |
| 12 | LXWJZMYZ | 0.2273 ± 0.00185 | ghijklmno | 38 | LXQJSDMZ | 0.22379 ± 0.00643 | ghijklmnop |
| 13 | LXSJLNNZ | 0.22777 ± 0.00141 | fghijklmno | 39 | LXQJSFZ-2 | 0.21436 ± 0.00656 | lmnop |
| 14 | LXQYNNZ | 0.21633 ± 0.00311 | klmnop | 40 | LMX1 | 0.22396 ± 0.00239 | ghijklmnop |
| 15 | LXCNNZ | 0.22209 ± 0.0121 | hijklmnop | 41 | LXSEZ | 0.22238 ± 0.00664 | hijklmnop |
| 16 | LXXZZ-2 | 0.21841 ± 0.0074 | klmnop | 42 | LXRHZ | 0.22202 ± 0.00831 | hijklmnop |
| 17 | LXYZCZ | 0.21886 ± 0.00524 | jklmnop | 43 | LXYZ | 0.21734 ± 0.00234 | klmnop |
| 18 | LXKYDSZ | 0.23908 ± 0.0003 | abcdefgh | 44 | LXMZ | 0.21845 ± 0.00663 | klmnop |
| 19 | LXEBCSZ | 0.2338 ± 0.00265 | bcdefghijk | 45 | LXDSZ | 0.22215 ± 0.0004 | hijklmnop |
| 20 | LXEBCCMZ | 0.23048 ± 0.00431 | defghijklm | 46 | LXSTZ | 0.21212 ± 0.00365 | nop |
| 21 | LXLJHPBB | 0.24204 ± 0.00591 | abcdefg | 47 | LXHTZ | 0.23116 ± 0.00672 | defghijkl |
| 22 | SXLXZ22 | 0.22982 ± 0.0068 | efghijklmn | 48 | LXHZ | 0.22509 ± 0.0064 | ghijklmno |
| 23 | SXLXZ23 | 0.2122 ± 0.00666 | mnop | 49 | LXDLLZ | 0.22425 ± 0.00325 | ghijklmno |
| 24 | SXLXZ24 | 0.2144 ± 0.0007 | klmnop | 50 | LXHHZ | 0.21893 ± 0.00281 | jklmnop |
| 25 | SXLXZ25 | 0.21333 ± 0.00527 | lmnop | 51 | LYHDZ | 0.20962 ± 0.00484 | op |
| 26 | SXLXZ26 | 0.21418 ± 0.00299 | lmnop | 52 | LXMZLMZ | 0.22536 ± 0.00646 | ghijklmno |
| 53 | LXMALSZ | 0.23322 ± 0.00405 | bcdefghijk | ||||
| Cluster | SRW/g | FLD/cm | FTP/cm | SW/g | SF/% |
|---|---|---|---|---|---|
| cluster1 | 9.21 ± 2.01 C | 3.17 ± 0.41 B | 2.48 ± 0.23 C | 0.49 ± 0.16 B | 29.72 ± 2.66 A |
| cluster2 | 13.4 ± 2.93 B | 3.99 ± 0.39 A | 2.71 ± 0.27 B | 0.4 ± 0.11 C | 30.17 ± 2.81 A |
| cluster3 | 19.25 ± 4.26 A | 3.88 ± 0.54 A | 3.2 ± 0.28 A | 0.81 ± 0.19 A | 30.66 ± 3.93 A |
| cluster | TS/% | TA/% | VC/mg/g | FER/% | GS/pg |
| cluster1 | 24.96 ± 2.53 b | 0.75 ± 0.34 A | 381.92 ± 83.48 B | 94.53 ± 1.71 B | 0.221 ± 0.007 B |
| cluster2 | 26.82 ± 3.23 a | 0.62 ± 0.19 B | 456.48 ± 72.41 A | 96.92 ± 0.96 A | 0.23 ± 0.01 A |
| cluster3 | 25.52 ± 3.11 ab | 0.51 ± 0.15 B | 322.05 ± 79.85 C | 95.1 ± 1.44 B | 0.225 ± 0.01 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Su, W.; Shi, M.; Xue, X.; Ren, H.; Wang, Y.; Zhao, A.; Li, D.; Liu, M. Genomic C-Value Variation Analysis in Jujube (Ziziphus jujuba Mill.) in the Middle Yellow River Basin. Plants 2023, 12, 858. https://doi.org/10.3390/plants12040858
Wu H, Su W, Shi M, Xue X, Ren H, Wang Y, Zhao A, Li D, Liu M. Genomic C-Value Variation Analysis in Jujube (Ziziphus jujuba Mill.) in the Middle Yellow River Basin. Plants. 2023; 12(4):858. https://doi.org/10.3390/plants12040858
Chicago/Turabian StyleWu, Hao, Wanlong Su, Meijuan Shi, Xiaofang Xue, Haiyan Ren, Yongkang Wang, Ailing Zhao, Dengke Li, and Mengjun Liu. 2023. "Genomic C-Value Variation Analysis in Jujube (Ziziphus jujuba Mill.) in the Middle Yellow River Basin" Plants 12, no. 4: 858. https://doi.org/10.3390/plants12040858