
Cytotoxic, Antibacterial, and Antioxidant Activities of the Leaf Extract of Sinningia bullata
Abstract
:1. Introduction
2. Results
2.1. Total Phenolic Content (TPC)
2.2. Total Flavonoid Content (TFC)
2.3. Antioxidant Activity
2.4. Antibacterial Activity
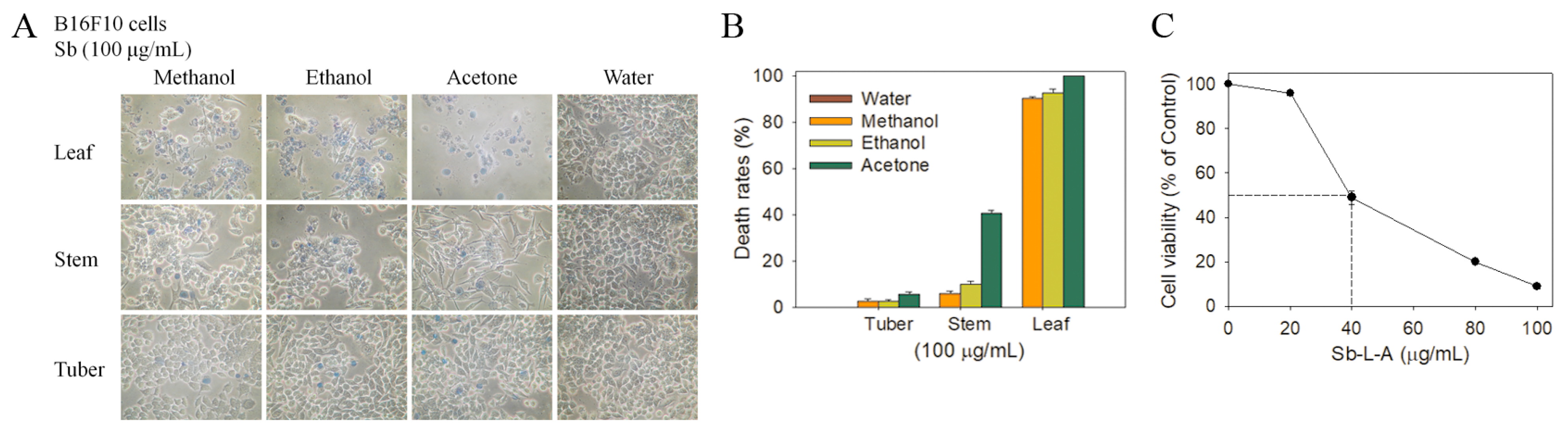
2.5. Anticancer Potential
2.6. Cytotoxic Activities against B16F10 Melanoma Cells
2.7. Cytotoxic Activities against 4T1 Mammary Carcinoma Cells
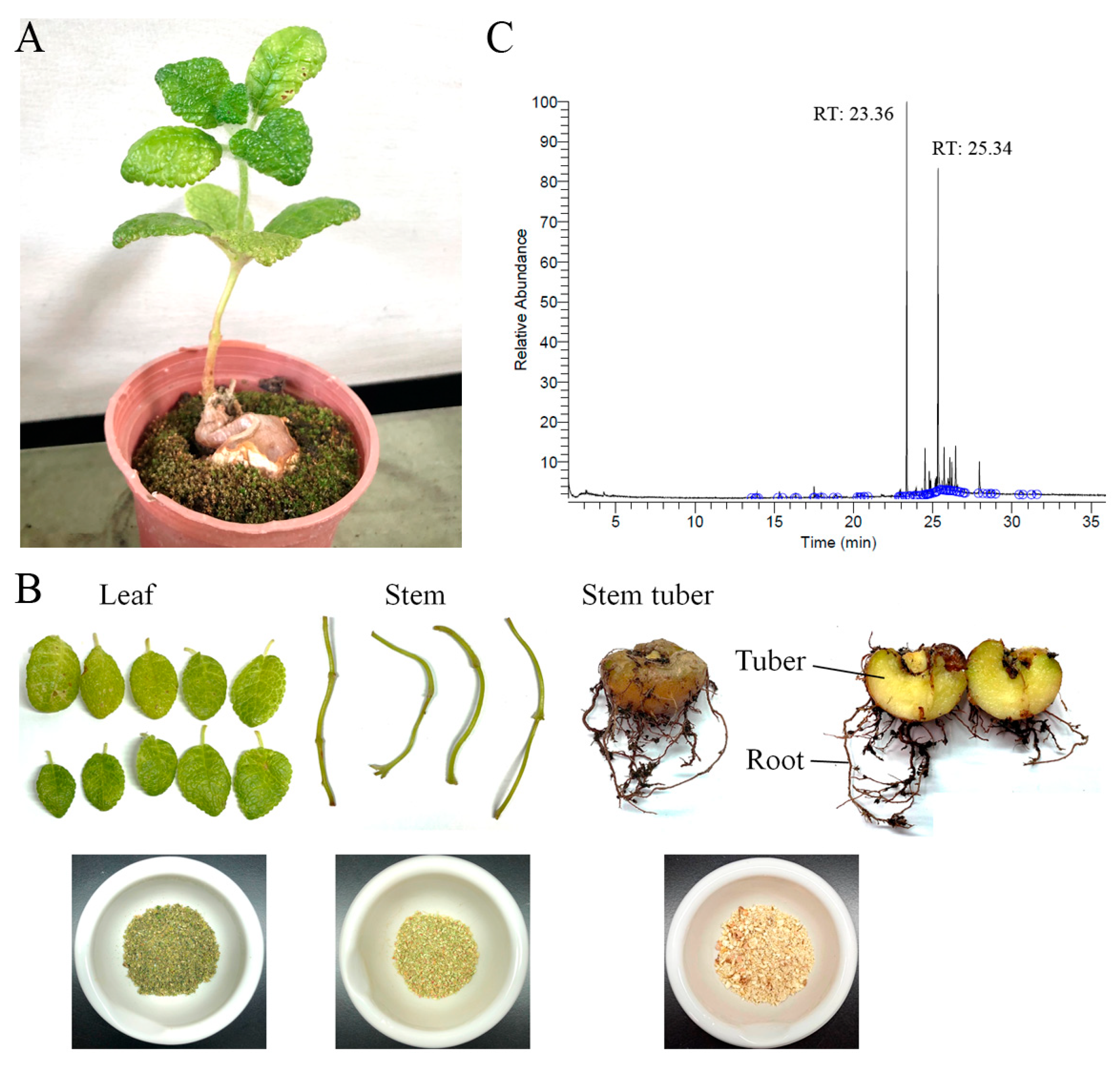
2.8. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis of Sb-L-A
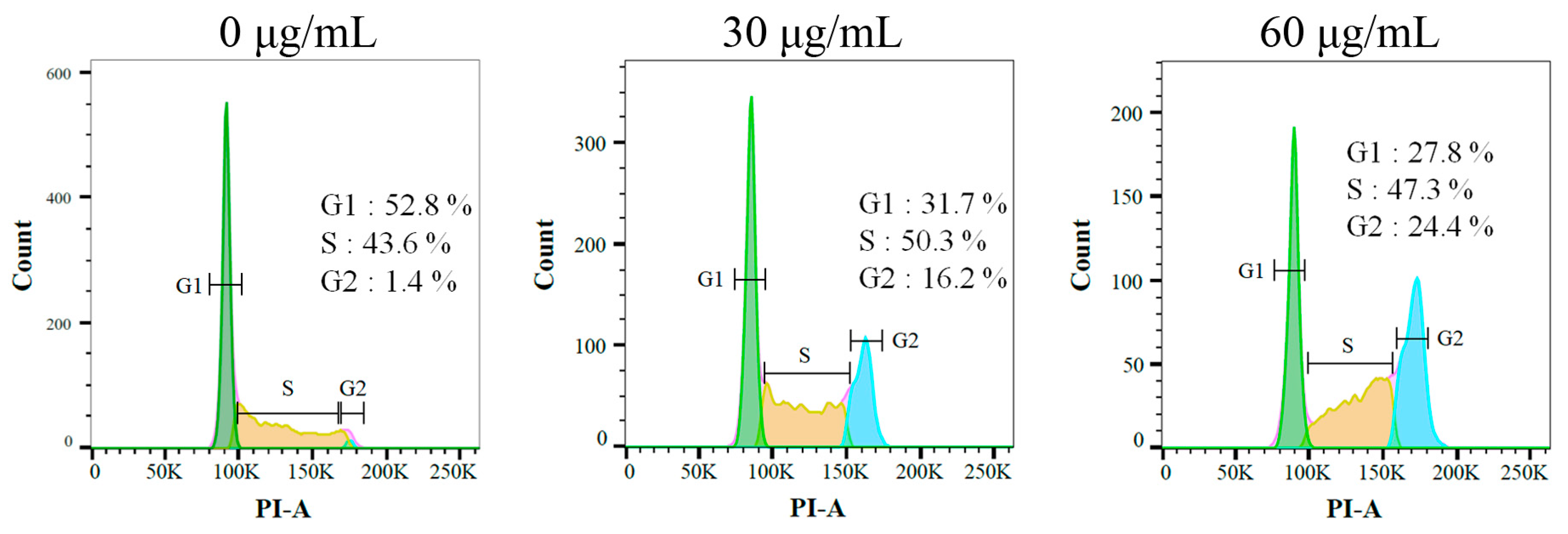
2.9. Sb-L-A Suppressed Melanoma Cell Proliferation by Inducing G2 Cell-Cycle Arrest
2.10. Co-Treatment of Sb-L-A with Epothilone B against B16F10 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Cell Culture
4.2. Plant Materials and Extract Preparations
4.3. Determination of TPC
4.4. Determination of TFC
4.5. Determination of Antioxidant Activity by DPPH Radical Scavenging Assay
4.6. GC-MS Analysis
4.7. Trypan Blue Cytotoxicity Assay
4.8. Chromatin Condensation Assay
4.9. Clonogenic Formation Assay
4.10. Wound-Healing Assay
4.11. Antibacterial Activities
4.12. Flow Analysis
4.13. MTT Cell Viability Assay
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Legardón, A.; García-Plazaola, J.I. Gesneriads, a Source of Resurrection and Double-Tolerant Species: Proposal of New Desiccation-and Freezing-Tolerant Plants and Their Physiological Adaptations. Biology 2023, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Merecz-Sadowska, A.; Sitarek, P.; Stelmach, J.; Zajdel, K.; Kucharska, E.; Zajdel, R. Plants as Modulators of Melanogenesis: Role of Extracts, Pure Compounds and Patented Compositions in Therapy of Pigmentation Disorders. Int. J. Mol. Sci. 2022, 23, 14787. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomas-Barberan, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.W.; Farooq, M.; Haseeb, M.; Choi, S. Role of Plant-Derived Active Constituents in Cancer Treatment and Their Mechanisms of Action. Cells 2022, 11, 1326. [Google Scholar] [CrossRef]
- Garcia-Oliveira, P.; Otero, P.; Pereira, A.G.; Chamorro, F.; Carpena, M.; Echave, J.; Fraga-Corral, M.; Simal-Gandara, J.; Prieto, M.A. Status and Challenges of Plant-Anticancer Compounds in Cancer Treatment. Pharmaceuticals 2021, 14, 157. [Google Scholar] [CrossRef] [PubMed]
- Gligorov, J.; Richard, S. Breast cancer: Weekly paclitaxel—Still preferred first-line taxane for mBC. Nat. Rev. Clin. Oncol. 2015, 12, 508–509. [Google Scholar] [CrossRef] [PubMed]
- Payton, S. Bladder cancer: Second-line nab-paclitaxel for advanced urothelial carcinoma. Nat. Rev. Urol. 2013, 10, 431. [Google Scholar] [CrossRef] [PubMed]
- Baird, R.D.; Tan, D.S.; Kaye, S.B. Weekly paclitaxel in the treatment of recurrent ovarian cancer. Nat. Rev. Clin. Oncol. 2010, 7, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.R.; Kozalak, G.; di Bari, I.; Babar, Q.; Niknam, Z.; Rasmi, Y.; Yong, K.W. In Vitro Human Cancer Models for Biomedical Applications. Cancers 2022, 14, 2284. [Google Scholar] [CrossRef]
- Zhou, Y.; Abel, G.A.; Hamilton, W.; Pritchard-Jones, K.; Gross, C.P.; Walter, F.M.; Renzi, C.; Johnson, S.; McPhail, S.; Elliss-Brookes, L.; et al. Diagnosis of cancer as an emergency: A critical review of current evidence. Nat. Rev. Clin. Oncol. 2017, 14, 45–56. [Google Scholar] [CrossRef]
- Truong, V.L.; Jeong, W.S. Cellular Defensive Mechanisms of Tea Polyphenols: Structure-Activity Relationship. Int. J. Mol. Sci. 2021, 22, 9109. [Google Scholar] [CrossRef] [PubMed]
- Islam, B.U.; Suhail, M.; Khan, M.K.; Zughaibi, T.A.; Alserihi, R.F.; Zaidi, S.K.; Tabrez, S. Polyphenols as anticancer agents: Toxicological concern to healthy cells. Phytother. Res. 2021, 35, 6063–6079. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Structure-activity relationships of flavonoids in the cellular antioxidant activity assay. J. Agric. Food Chem. 2008, 56, 8404–8411. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, Y.; Liu, X.; Chen, X.; Chen, R. Comprehensive Review of Recent Advances in Chiral A-Ring Flavonoid Containing Compounds: Structure, Bioactivities, and Synthesis. Molecules 2023, 28, 365. [Google Scholar] [CrossRef]
- Dini, I.; Grumetto, L. Recent Advances in Natural Polyphenol Research. Molecules 2022, 27, 8777. [Google Scholar] [CrossRef]
- Tommasi, R.; Brown, D.G.; Walkup, G.K.; Manchester, J.I.; Miller, A.A. ESKAPEing the labyrinth of antibacterial discovery. Nat. Rev. Drug Discov. 2015, 14, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, A. The cost of resistance. Nat. Rev. Microbiol. 2022, 20, 317. [Google Scholar] [CrossRef]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular mechanisms of antibiotic resistance revisited. Nat. Rev. Microbiol. 2022, 1–16. [Google Scholar] [CrossRef]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef]
- Kokoska, L.; Kloucek, P.; Leuner, O.; Novy, P. Plant-Derived Products as Antibacterial and Antifungal Agents in Human Health Care. Curr. Med. Chem. 2019, 26, 5501–5541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, G.; Han, K.; Sun, D.; Zhou, N.; Song, Z.; Liu, H.; Li, J.; Li, G. Applications of Molecular Imprinting Technology in the Study of Traditional Chinese Medicine. Molecules 2023, 28, 301. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug. Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Pereira, A.M.; Cidade, H.; Tiritan, M.E. Stereoselective Synthesis of Flavonoids: A Brief Overview. Molecules 2023, 28, 426. [Google Scholar] [CrossRef] [PubMed]
- Golonko, A.; Olichwier, A.J.; Swislocka, R.; Szczerbinski, L.; Lewandowski, W. Why Do Dietary Flavonoids Have a Promising Effect as Enhancers of Anthracyclines? Hydroxyl Substituents, Bioavailability and Biological Activity. Int. J. Mol. Sci. 2023, 24, 391. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Kozyra, P.; Krasowska, D.; Pitucha, M. New Potential Agents for Malignant Melanoma Treatment-Most Recent Studies 2020–2022. Int. J. Mol. Sci. 2022, 23, 6084. [Google Scholar] [CrossRef]
- Abdel-Salam, M.A.L.; Pinto, B.; Cassali, G.; Bueno, L.; Pêgas, G.; Oliveira, F.; Silva, I.; Klein, A.; Souza-Fagundes, E.M.; de Lima, M.E.; et al. LyeTx I-b Peptide Attenuates Tumor Burden and Metastasis in a Mouse 4T1 Breast Cancer Model. Antibiotics 2021, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Pulaski, B.A.; Ostrand-Rosenberg, S. Mouse 4T1 breast tumor model. Curr. Protoc. Immunol. 2001, 39, 20–22. [Google Scholar] [CrossRef]
- Lee, J.J.; Kelly, W.K. Epothilones: Tubulin polymerization as a novel target for prostate cancer therapy. Nat. Clin. Pract. Oncol. 2009, 6, 85–92. [Google Scholar] [CrossRef]
- Larkin, J.M.; Kaye, S.B. Epothilones in the treatment of cancer. Expert Opin. Investig. Drugs 2006, 15, 691–702. [Google Scholar] [CrossRef]
- Peng, B.; Zhang, S.Y.; Chan, K.I.; Zhong, Z.F.; Wang, Y.T. Novel Anti-Cancer Products Targeting AMPK: Natural Herbal Medicine against Breast Cancer. Molecules 2023, 28, 740. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Zhou, L.; Li, L.; Zhou, P.; Shen, Z. Nano-Based Drug Delivery of Polyphenolic Compounds for Cancer Treatment: Progress, Opportunities, and Challenges. Pharmaceuticals 2023, 16, 101. [Google Scholar] [CrossRef] [PubMed]
- Almeida, T.C.; Seibert, J.B.; Amparo, T.R.; de Souza, G.H.B.; da Silva, G.N.; Dos Santos, O.D.H. Modulation of Long Non-coding RNAs by Different Classes of Secondary Metabolites from Plants: A Mini-review on Antitumor Effects. Mini-Rev. Med. Chem. 2022, 22, 1232–1255. [Google Scholar] [CrossRef]
- Bitchagno, G.T.M.; Nchiozem-Ngnitedem, V.A.; Melchert, D.; Fobofou, S.A. Demystifying racemic natural products in the homochiral world. Nat. Rev. Chem. 2022, 6, 806–822. [Google Scholar] [CrossRef]
- Ouyang, L.; Luo, Y.; Tian, M.; Zhang, S.Y.; Lu, R.; Wang, J.H.; Kasimu, R.; Li, X. Plant natural products: From traditional compounds to new emerging drugs in cancer therapy. Cell Prolif. 2014, 47, 506–515. [Google Scholar] [CrossRef]
- Patridge, E.; Gareiss, P.; Kinch, M.S.; Hoyer, D. An analysis of FDA-approved drugs: Natural products and their derivatives. Drug Discov. Today 2016, 21, 204–207. [Google Scholar] [CrossRef]
- Szewczyk, K.; Chojnacka, A.; Górnicka, M. Tocopherols and Tocotrienols-Bioactive Dietary Compounds; What Is Certain, What Is Doubt? Int. J. Mol. Sci. 2021, 22, 6222. [Google Scholar] [CrossRef]
- Asokkumar, S.; Naveenkumar, C.; Raghunandhakumar, S.; Kamaraj, S.; Anandakumar, P.; Jagan, S.; Devaki, T. Antiproliferative and antioxidant potential of beta-ionone against benzo(a)pyrene-induced lung carcinogenesis in Swiss albino mice. Mol. Cell. Biochem. 2012, 363, 335–345. [Google Scholar] [CrossRef]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef]
- Bailly, C. Yuanhuacin and Related Anti-Inflammatory and Anticancer Daphnane Diterpenes from Genkwa Flos-An Overview. Biomolecules 2022, 12, 192. [Google Scholar] [CrossRef]
- Ansari, M.; Emami, S. β-Ionone and its analogs as promising anticancer agents. Eur. J. Med. Chem. 2016, 123, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef] [PubMed]
- Riva, D.; Barison, A.; Stefanello, M.É.A.; Poliquesi, C.B.; Ruiz, A.L.T.G.; Salvador, M.J. Estudo químico de Sinningia allagophylla guiado por testes de atividade antiproliferativa. Quim. Nova 2012, 35, 974–977. [Google Scholar] [CrossRef]
- Winiewski, V.; Serain, A.F.; Sá, E.L.; Salvador, M.J.; Stefanello, M.É.A. Chemical constituents of Sinningia mauroana and screening of its extracts for antimicrobial, antioxidant and cytotoxic activities. Quim. Nova 2020, 43, 181–187. [Google Scholar] [CrossRef]
- Verdan, M.H.; Mera de Souza, L.; Ernesto de Carvalho, J.; Vendramini Costa, D.B.; Salvador, M.J.; Barison, A.; Alves Stefanello, M.É. Two new hydronaphthoquinones from Sinningia aggregata (Gesneriaceae) and cytotoxic activity of aggregatin D. Chem. Biodiv. 2015, 12, 148–152. [Google Scholar] [CrossRef]
- Silva, A.S.; Amorim, M.S.; Fonseca, M.M.; Salvador, M.J.; Sá, E.L.; Stefanello, M.É.A. A new cytotoxic naphthoquinone and other chemical constituents of Sinningia reitzii. J. Braz. Chem. Soc. 2019, 30, 2060–2065. [Google Scholar] [CrossRef]
- Verdan, M.H.; Unemoto, L.K.; Faria, R.T.; Salvador, M.J.; Sá, E.L.; Barison, A.; Stefanello, M.É.A. Leucotrichoic acid, a novel sesquiterpene from Sinningia leucotricha (Gesneriaceae). Tet. Let. 2013, 54, 4735–4737. [Google Scholar] [CrossRef]
- Amorim, M.S.; Serain, A.F.; Salvador, M.J.; Stefanello, M.É.A. Chemical constituents of Sinningia hatschbachii. Nat. Prod. Commun. 2017, 12, 1763–1764. [Google Scholar] [CrossRef]
- Serain, A.F.; Silvério, S.E.B.; De Lourenço, C.C.; Nunes, V.K.; Corrêa, W.R.; Stefanello, M.É.A.; Salvador, M.J. Development of Sinningia magnifica (Otto & A. Dietr.) Wiehler (Gesneriaceae) tissue culture for in vitro production of quinones and bioactive molecules. Ind. Crops Prod. 2021, 159, 113046. [Google Scholar] [CrossRef]
- Verdan, M.H.; Barison, A.; Sá, E.L.; Salvador, M.J.; Poliquesi, C.B.; Eberlin, M.N.; Stefanello, M.É.A. Lactones and quinones from the tubers of Sinningia aggregata. J. Nat. Prod. 2010, 73, 1434–1437. [Google Scholar] [CrossRef]
- Winiewski, V.; Verdan, M.H.; Ribeiro, M.A.; Hernandez-Tasco, A.J.; Salvador, M.J.; Stefanello, M.É.A. Warmingiins A and B, two new dimeric naphthoquinone derivatives from Sinningia warmingii (Gesneriaceae). J. Braz. Chem. Soc. 2017, 28, 598–602. [Google Scholar] [CrossRef]
- Winiewski, V.; Fonseca, M.M.; Salvador, M.J.; Sá, E.L.; Stefanello, M.É.A. Three new naphthoquinones from the tubers of Sinningia mauroana. Nat. Prod. Res. 2023, 37, 263–268. [Google Scholar] [CrossRef]
- Winiewski, V.; Silva, A.S.; Alvarez, K.D.C.; Sá, E.L.; Salvador, M.J.; Stefanello, M.É.A. Antioxidant naphthoquinones of Sinningia reitzii from Santa Catarina State, Brazil. Quim. Nova 2021, 44, 284–287. [Google Scholar] [CrossRef]
- Lin, E.S.; Huang, C.Y. Cytotoxic Activities and the Allantoinase Inhibitory Effect of the Leaf Extract of the Carnivorous Pitcher Plant Nepenthes miranda. Plants 2022, 11, 2265. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.H.; Huang, Y.H.; Lin, E.S.; Chen, C.J.; Huang, C.Y. Plumbagin, a Natural Product with Potent Anticancer Activities, Binds to and Inhibits Dihydroorotase, a Key Enzyme in Pyrimidine Biosynthesis. Int. J. Mol. Sci. 2021, 22, 6861. [Google Scholar] [CrossRef]
- Liu, H.W.; Chiang, W.Y.; Huang, Y.H.; Huang, C.Y. The Inhibitory Effects and Cytotoxic Activities of the Stem Extract of Sarracenia purpurea against Melanoma Cells and the SsbA Protein. Plants 2022, 11, 3164. [Google Scholar] [CrossRef]
- Huang, Y.H.; Chiang, W.Y.; Chen, P.J.; Lin, E.S.; Huang, C.Y. Anticancer and Antioxidant Activities of the Root Extract of the Carnivorous Pitcher Plant Sarracenia purpurea. Plants 2022, 11, 1668. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Lien, Y.; Chen, J.H.; Lin, E.S.; Huang, C.Y. Identification and characterization of dihydropyrimidinase inhibited by plumbagin isolated from Nepenthes miranda extract. Biochimie 2020, 171–172, 124–135. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001, 21, A-3B. [Google Scholar]
- Larsson, R.; Nygren, P. A rapid fluorometric method for semiautomated determination of cytotoxicity and cellular proliferation of human tumor cell lines in microculture. Anticancer Res. 1989, 9, 1111–1119. [Google Scholar]
- Chen, M.H.; Yang, W.L.; Lin, K.T.; Liu, C.H.; Liu, Y.W.; Huang, K.W.; Chang, P.M.; Lai, J.M.; Hsu, C.N.; Chao, K.M.; et al. Gene expression-based chemical genomics identifies potential therapeutic drugs in hepatocellular carcinoma. PLoS ONE 2011, 6, e27186. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf | Stem | Tuber | |
|---|---|---|---|
| Water | 16.4 ± 0.4 | 16.6 ± 0.3 | 6.5 ± 0.2 |
| Methanol | 39.6 ± 1.0 | 46.6 ± 0.8 | 21.7 ± 0.4 |
| Ethanol | 40.3 ± 0.9 | 47.1 ± 1.1 | 23.7 ± 0.3 |
| Acetone | 46.2 ± 1.2 | 49.7 ± 0.8 | 28.4 ± 0.8 |
| Leaf | Stem | Tuber | |
|---|---|---|---|
| Water | 31.4 ± 0.4 | 21.1 ± 0.3 | 16.0 ± 0.3 |
| Methanol | 62.1 ± 1.2 | 50.3 ± 0.8 | 31.9 ± 0.7 |
| Ethanol | 63.0 ± 1.0 | 51.9 ± 0.6 | 33.8 ± 0.6 |
| Acetone | 65.8 ± 1.3 | 52.5 ± 0.6 | 37.1 ± 0.7 |
| IC50 (μg/mL) | |||
|---|---|---|---|
| Solvent | Leaf | Stem | Tuber |
| Methanol | 250.9 ± 4.2 | 298.4 ± 4.6 | 340.1 ± 5.6 |
| Ethanol | 232.2 ± 4.0 | 294.0 ± 5.2 | 321.2 ± 4.2 |
| Acetone | 180.8 ± 2.8 | 266.3 ± 3.8 | 293.2 ± 4.6 |
| Zone of Inhibition (mm) | ||||
|---|---|---|---|---|
| Material | Solvent | E. coli | S. aureus | P. aeruginosa |
| Leaf | Water | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Methanol | 10 ± 1 | 27 ± 1 | 7 ± 0 | |
| Ethanol | 10 ± 0 | 27 ± 1 | 8 ± 0 | |
| Acetone | 11 ± 1 | 30 ± 2 | 8 ± 1 | |
| Stem | Water | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Methanol | 7 ± 0 | 18 ± 1 | 7 ± 0 | |
| Ethanol | 8 ± 1 | 19 ± 1 | 7 ± 0 | |
| Acetone | 8 ± 1 | 21 ± 1 | 7 ± 1 | |
| Tuber | Water | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Methanol | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| Ethanol | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| Acetone | 0 ± 0 | 9 ± 1 | 0 ± 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.-J.; Lin, E.-S.; Su, H.-H.; Huang, C.-Y. Cytotoxic, Antibacterial, and Antioxidant Activities of the Leaf Extract of Sinningia bullata. Plants 2023, 12, 859. https://doi.org/10.3390/plants12040859
Chen P-J, Lin E-S, Su H-H, Huang C-Y. Cytotoxic, Antibacterial, and Antioxidant Activities of the Leaf Extract of Sinningia bullata. Plants. 2023; 12(4):859. https://doi.org/10.3390/plants12040859
Chicago/Turabian StyleChen, Pin-Jui, En-Shyh Lin, Hsin-Hui Su, and Cheng-Yang Huang. 2023. "Cytotoxic, Antibacterial, and Antioxidant Activities of the Leaf Extract of Sinningia bullata" Plants 12, no. 4: 859. https://doi.org/10.3390/plants12040859