

Chemistry, Occurrence, Properties, Applications, and Encapsulation of Carotenoids—A Review
 ,
,
Abstract
:
1. Introduction
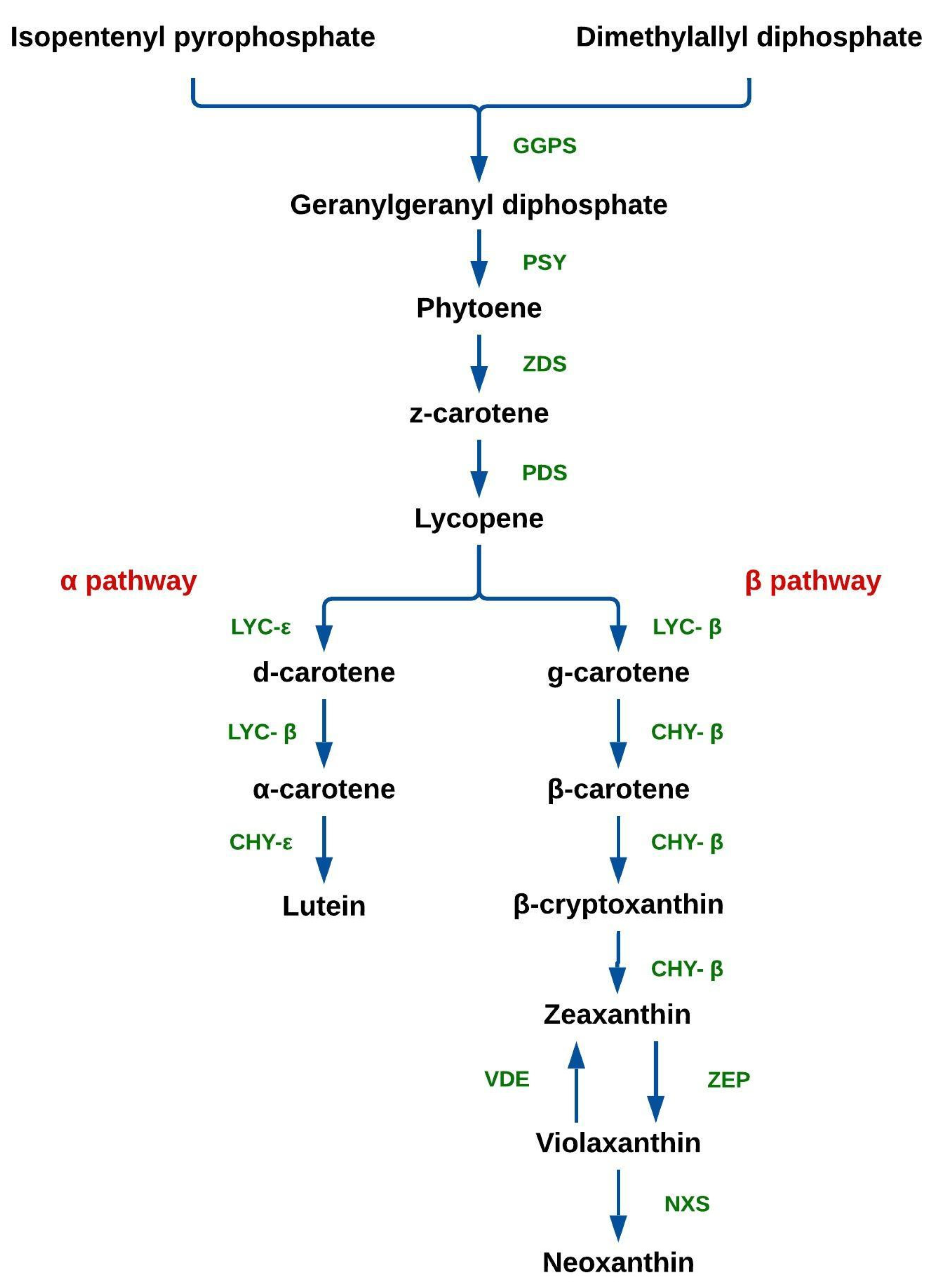
2. Chemistry of Carotenoids
3. Natural Occurrence of Carotenoids
4. Bioavailability of Carotenoids
5. Use of Carotenoids
5.1. Colorants
5.2. Vitamin a Activity
5.3. Antioxidants
5.4. Biological Properties and Other Uses of Carotenoids
6. Stability of Carotenoids
7. Encapsulation Techniques for Carotenoid Protection
7.1. Microencapsulation
7.2. Nanoencapsulation
7.3. Supercritical Fluids
8. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.; Tsao, R.; Deng, Z. Factors Affecting the Antioxidant Potential and Health Benefits of Plant Foods. Can. J. Plant Sci. 2012, 92, 1101–1111. [Google Scholar] [CrossRef]
- Lemmens, L.; Colle, I.; Van Buggenhout, S.; Palmero, P.; Van Loey, A.; Hendrickx, M. Carotenoid Bioaccessibility in Fruit- and Vegetable-Based Food Products as Affected by Product (Micro)Structural Characteristics and the Presence of Lipids: A Review. Trends Food Sci. Technol. 2014, 38, 125–135. [Google Scholar] [CrossRef]
- Saini, R.K.; Nile, S.H.; Park, S.W. Carotenoids from Fruits and Vegetables: Chemistry, Analysis, Occurrence, Bioavailability and Biological Activities. Food Res. Int. 2015, 76, 735–750. [Google Scholar] [CrossRef] [PubMed]
- Yahia, E.M.; Gutiérrez-Orozco, F.; Arvizu-de Leon, C. Phytochemical and Antioxidant Characterization of Mamey (Pouteria sapota Jacq. H.E. Moore & Stearn) Fruit. Food Res. Int. 2011, 44, 2175–2181. [Google Scholar] [CrossRef]
- Butnariu, M. Methods of Analysis (Extraction, Separation, Identification and Quantification) of Carotenoids from Natural Products. J. Ecosyst. Ecography 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Santos, D.T.; Meireles, M.A.A. Carotenoid Pigments Encapsulation: Fundamentals, Techniques and Recent Trends. Open Chem. Eng. J. 2010, 4, 42–50. [Google Scholar] [CrossRef]
- Salter-Venzon, D.; Kazlova, V.; Izzy Ford, S.; Intra, J.; Klosner, A.E.; Gellenbeck, K.W. Evidence for Decreased Interaction and Improved Carotenoid Bioavailability by Sequential Delivery of a Supplement. Food Sci. Nutr. 2017, 5, 424–433. [Google Scholar] [CrossRef]
- Guerrero-Legarreta, I.; López-Hernández, E.; Armenta-López, R.E.; García-Barrientos, R. Pigmentos. In Química de Alimentos; Badui-Dergal, S., Ed.; Pearson: Mexico City, Mexico, 2013; p. 379. [Google Scholar]
- Schawartz, S.J.; Cooperstone, J.L.; Cichon, M.J.; von Elbe, J.H.; Giusti, M.M. Colorants. In Fennema’s Food Chemistry; Damodaran, S., Parkin, K.L., Eds.; CRC PressTaylor & Francis: Boca Raton, FL, USA, 2017; p. 681. [Google Scholar]
- Howitt, C.A.; Pogson, B.J. Carotenoid Accumulation and Function in Seeds and Non-Green Tissues. Plant Cell Environ. 2006, 29, 435–445. [Google Scholar] [CrossRef]
- Perera, C.O.; Yen, G.M. Functional Properties of Carotenoids in Human Health. Int. J. Food Prop. 2007, 10, 201–230. [Google Scholar] [CrossRef]
- Kang, L.; Park, S.-C.; Ji, C.Y.; Kim, H.S.; Lee, H.-S.; Kwak, S.-S. Metabolic Engineering of Carotenoids in Transgenic Sweetpotato. Breed. Sci. 2017, 67, 27–34. [Google Scholar] [CrossRef]
- Olmedilla-Alonso, B. Carotenoids: Content in Foods, in Diet and Bioavailability. COST Action EUROCAROTEN CA15136. Sci. Newsl. 2017, 2, 1–9. [Google Scholar] [CrossRef]
- Food-Info Foundation E-Numbers: E100–E200 Food Colours. Available online: http://www.food-info.net/uk/e/e100-200.htm (accessed on 2 June 2020).
- Liu, X.; Luo, Q.; Cao, Y.; Goulette, T.; Liu, X.; Xiao, H. Mechanism of Different Stereoisomeric Astaxanthin in Resistance to Oxidative Stress in Caenorhabditis elegans. J. Food Sci. 2016, 81, H2280–H2287. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Hosokawa, M.; Miyashita, K. Fucoxanthin: A Marine Carotenoid Exerting Anti-Cancer Effects by Affecting Multiple Mechanisms. Mar. Drugs 2013, 11, 5130–5147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gul, K.; Tak, A.; Singh, A.K.; Singh, P.; Yousuf, B.; Wani, A.A. Chemistry, Encapsulation, and Health Benefits of β-Carotene—A Review. Cogent Food Agric. 2015, 1, 1018696. [Google Scholar] [CrossRef]
- Yoon, G.-A.; Yeum, K.-J.; Cho, Y.-S.; Chen, C.-Y.O.; Tang, G.; Blumberg, J.B.; Russell, R.M.; Yoon, S.; Lee-Kim, Y.C. Carotenoids and Total Phenolic Contents in Plant Foods Commonly Consumed in Korea. Nutr. Res. Pract. 2012, 6, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagetti, M.; Facco, E.M.P.; Piccolo, J.; Hirsch, G.E.; Rodriguez-Amaya, D.; Kobori, C.N.; Vizzotto, M.; Emanuelli, T. Physicochemical Characterization and Antioxidant Capacity of Pitanga Fruits (Eugenia uniflora L.). Food Sci. Technol. 2011, 31, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Chandrika, U.G.; Fernando, K.S.S.P.; Ranaweera, K.K.D.S. Carotenoid Content and in Vitro Bioaccessibility of Lycopene from Guava (Psidium guajava) and Watermelon (Citrullus lanatus) by High-Performance Liquid Chromatography Diode Array Detection. Int. J. Food Sci. Nutr. 2009, 60, 558–566. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Victoria-Campos, C.I.; de Ornelas-Paz, J.J. Absorption of Carotenoids and Mechanisms Involved in Their Health-Related Properties. In Carotenoids in Nature; Stange, C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 415–454. ISBN 978-3-319-39124-3. [Google Scholar]
- Bohn, T.; Desmarchelier, C.; Dragsted, L.O.; Nielsen, C.S.; Stahl, W.; Rühl, R.; Keijer, J.; Borel, P. Host-Related Factors Explaining Interindividual Variability of Carotenoid Bioavailability and Tissue Concentrations in Humans. Mol. Nutr. Food Res. 2017, 61, 1600685. [Google Scholar] [CrossRef] [Green Version]
- Borel, P.; Desmarchelier, C.; Dumont, U.; Halimi, C.; Lairon, C.; Paige, C.; Sébédio, L.; Bulsson, C.; Buffiére, C.; Rémond, D. Dietary Calcium Impairs Tomato Lycopene Bioavailability in Healthy Humans. Br. J. Nutr. 2016, 116, 2091–2096. [Google Scholar] [CrossRef]
- Corte-Real, J.; Guignard, C.; Gantenbein, M.; Weber, B.; Burgard, K.; Hoffmann, L.; Richling, E.; Bohn, T. No Influence of Supplemental Dietary Calcium Intake on the Bioavailability of Spinach Carotenoids in Humans. Br. J. Nutr. 2017, 117, 1560–1569. [Google Scholar] [CrossRef]
- Marriage, B.J.; Williams, J.A.; Choe, Y.S.; Maki, K.C.; Vurma, M.; DeMichele, S.J. Mono- and Diglycerides Improve Lutein Absorption in Healthy Adults: A Randomised, Double-Blind, Cross-over, Single-Dose Study. Br. J. Nutr. 2017, 118, 813–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, W.S.; Zhou, Y.; Crane, A.; Dixon, P.; Quadt, F.; Flendring, L.M. Modeling the Dose Effects of Soybean Oil in Salad Dressing on Carotenoid and Fat-Soluble Vitamin Bioavailability in Salad Vegetables. Am. J. Clin. Nutr. 2017, 106, 1041–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aschoff, J.K.; Rolke, C.L.; Breusing, N.; Bosy-Westphal, A.; Högel, J.; Carle, R.; Schweiggert, R.M. Bioavailability of β-Cryptoxanthin Is Greater from Pasteurized Orange Juice than from Fresh Oranges—A Randomized Cross-over Study. Mol. Nutr. Food Res. 2015, 59, 1896–1904. [Google Scholar] [CrossRef] [PubMed]
- Vimala, B.; Nambisan, B.; Hariprakash, B. Retention of Carotenoids in Orange-Fleshed Sweet Potato during Processing. J. Food Sci. Technol. 2011, 48, 520–524. [Google Scholar] [CrossRef] [Green Version]
- Odriozola-Serrano, I.; Soliva-Fortuny, R.; Hernández-Jover, T.; Martín-Belloso, O. Carotenoid and Phenolic Profile of Tomato Juices Processed by High Intensity Pulsed Electric Fields Compared with Conventional Thermal Treatments. Food Chem. 2009, 112, 258–266. [Google Scholar] [CrossRef]
- Sanches-Silva, A.; Albuquerque, T.G.; Finglas, P.; Ribeiro, T.; Valente, A.; Vasilopoulou, E.; Trichopoulou, A.; Alexieva, I.; Boyko, N.; Costea, C.-E.; et al. Carotenoids, Vitamins (A, B2, C and E) and Total Folate of Traditional Foods from Black Sea Area Countries: Health-Promoting Compounds in Traditional Foods from Black Sea Area. J. Sci. Food Agric. 2013, 93, 3545–3557. [Google Scholar] [CrossRef] [PubMed]
- Lehto, S.; Buchweitz, M.; Klimm, A.; Straßburger, R.; Bechtold, C.; Ulberth, F. Comparison of Food Colour Regulations in the EU and the US: A Review of Current Provisions. Food Addit. Contam. Part A 2017, 34, 335–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Drug Administration Color Additive Status List. Available online: https://www.fda.gov/industry/color-additive-inventories/color-additive-status-list (accessed on 2 June 2020).
- Beltrán-de-Miguel, B.; Estévez-Santiago, R.; Olmedilla-Alonso, B. Assessment of Dietary Vitamin A Intake (Retinol, α-Carotene, β-Carotene, β-Cryptoxanthin) and Its Sources in the National Survey of Dietary Intake in Spain (2009–2010). Int. J. Food Sci. Nutr. 2015, 66, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Chacón-Ordóñez, T.; Esquivel, P.; Jiménez, V.M.; Carle, R.; Schweiggert, R.M. Deposition Form and Bioaccessibility of Keto-Carotenoids from Mamey Sapote (Pouteria sapota), Red Bell Pepper (Capsicum annuum), and Sockeye Salmon (Oncorhynchus nerka) Filet. J. Agric. Food Chem. 2016, 64, 1989–1998. [Google Scholar] [CrossRef]
- Cruz-Bojórquez, R.M.; González-Gallego, J.; Sánchez-Collado, P. Propiedades Funcionales y Beneficios Para La Salud Del Licopeno. Nutr. Hosp. 2013, 28, 6–15. [Google Scholar] [CrossRef]
- Cañete, A.; Cano, E.; Muñoz-Chápuli, R.; Carmona, R. Role of Vitamin A/Retinoic Acid in Regulation of Embryonic and Adult Hematopoiesis. Nutrients 2017, 9, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, R.; Meléndez-Martínez, A.J.; Vicario, I.M.; Alcalde, M.J. Carotenoid and Vitamin A Contents in Biological Fluids and Tissues of Animals as an Effect of the Diet: A Review. Food Rev. Int. 2015, 31, 319–340. [Google Scholar] [CrossRef]
- National Institutes of Health Vitamin A. Available online: https://ods.od.nih.gov/factsheets/VitaminA-HealthProfessional/ (accessed on 15 June 2020).
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic Effects of Inflammation on Vitamin A and Carotenoids in Humans and Animal Models. Adv. Nutr. Int. Rev. J. 2017, 8, 197–212. [Google Scholar] [CrossRef] [Green Version]
- Cser, M.A.; Majchrzak, D.; Rust, P.; Sziklai-László, I.; Kovács, I.; Bocskai, E.; Elmadfa, I. Serum Carotenoid and Retinol Levels during Childhood Infections. Ann. Nutr. Metab. 2004, 48, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Kitts, D.D. Antioxidant Property of Coffee Components: Assessment of Methods That Define Mechanisms of Action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [Green Version]
- Nagayama, J.; Noda, K.; Uchikawa, T.; Maruyama, I.; Shimomura, H.; Miyahara, M. Effect of Maternal Chlorella Supplementation on Carotenoid Concentration in Breast Milk at Early Lactation. Int. J. Food Sci. Nutr. 2014, 65, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and Pro-Oxidant Activities of Carotenoids and Their Oxidation Products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Xavier, A.A.O.; Pérez-Gálvez, A. Carotenoids as a Source of Antioxidants in the Diet. In Carotenoids in Nature; Stange, C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 359–375. ISBN 978-3-319-39124-3. [Google Scholar]
- Pons, E.; Alquézar, B.; Rodríguez, A.; Martorell, P.; Genovés, S.; Ramón, D.; Rodrigo, M.J.; Zacarías, L.; Peña, L. Metabolic Engineering of β-Carotene in Orange Fruit Increases Its in Vivo Antioxidant Properties. Plant Biotechnol. J. 2014, 12, 17–27. [Google Scholar] [CrossRef] [PubMed]
- You, J.S.; Jeon, S.; Byun, Y.J.; Koo, S.; Choi, S.S. Enhanced Biological Activity of Carotenoids Stabilized by Phenyl Groups. Food Chem. 2015, 177, 339–345. [Google Scholar] [CrossRef]
- Yazaki, K.; Yoshikoshi, C.; Oshiro, S.; Yanase, S. Supplemental Cellular Protection by a Carotenoid Extends Lifespan via Ins/IGF-1 Signaling in Caenorhabditis elegans. Oxid. Med. Cell. Longev. 2011, 2011, 1–9. [Google Scholar] [CrossRef]
- Lashmanova, E.; Proshkina, E.; Zhikrivetskaya, S.; Shevchenko, O.; Marusich, E.; Leonov, S.; Melerzanov, A.; Zhavoronkov, A.; Moskalev, A. Fucoxanthin Increases Lifespan of Drosophila melanogaster and Caenorhabditis elegans. Pharmacol. Res. 2015, 100, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Van Rooyen, J.; Esterhuyse, A.J.; Engelbrecht, A.-M.; Du Toit, E.F. Health Benefits of a Natural Carotenoid Rich Oil: A Proposed Mechanism of Protection against Ischaemia/Reperfusion Injury. Asia Pac. J. Clin. Nutr. 2008, 17, 316–319. [Google Scholar] [PubMed]
- Milde, J.; Elstner, E.F.; Graßmann, J. Synergistic Effects of Phenolics and Carotenoids on Human Low-Density Lipoprotein Oxidation. Mol. Nutr. Food Res. 2007, 51, 956–961. [Google Scholar] [CrossRef] [PubMed]
- Varakumar, S.; Kumar, Y.S.; Reddy, O.V.S. Carotenoid Composition of Mango (Mangifera indica L.) Wine and Its Antioxidant Activity. J. Food Biochem. 2011, 35, 1538–1547. [Google Scholar] [CrossRef]
- Sangha, J.S.; Fan, D.; Banskota, A.H.; Stefanova, R.; Khan, W.; Hafting, J.; Craigie, J.; Critchley, A.T.; Prithiviraj, B. Bioactive Components of the Edible Strain of Red Alga, Chondrus crispus, Enhance Oxidative Stress Tolerance in Caenorhabditis elegans. J. Funct. Foods 2013, 5, 1180–1190. [Google Scholar] [CrossRef]
- Brown, M.A.; Storlien, L.H.; Huang, X.-F.; Tapsell, L.C.; Else, P.L.; Higgins, J.A.; Brown, I.L. Dietary Fat and Carbohydrate Composition: Metabolic Disease. In Fat Detection: Taste, Texture, and Post Ingestive Effects; Frontiers in Neuroscience; Montmayeur, J.-P., le Coutre, J., Eds.; CRC Press: Boca Raton, FL, USA, 2010; ISBN 978-1-4200-6775-0. [Google Scholar]
- Muskiet, F.A.J. Pathophysiology and Evolutionary Aspects of Dietary Fats and Long-Chain Polyunsaturated Fatty Acids across the Life Cycle. In Fat Detection: Taste, Texture, and Post Ingestive Effects; Frontiers in Neuroscience; Montmayeur, J.-P., le Coutre, J., Eds.; CRC Press: Boca Raton, FL, USA, 2010; ISBN 978-1-4200-6775-0. [Google Scholar]
- Shegokar, R.; Mitri, K. Carotenoid Lutein: A Promising Candidate for Pharmaceutical and Nutraceutical Applications. J. Diet. Suppl. 2012, 9, 183–210. [Google Scholar] [CrossRef]
- Dai, Z.; Wang, R.; Ang, L.-W.; Low, Y.-L.; Yuan, J.-M.; Koh, W.-P. Protective Effects of Dietary Carotenoids on Risk of Hip Fracture in Men: The Singapore Chinese Health Study: DIETARY CAROTENOIDS in RELATION to HIP FRACTURE RISK in MEN. J. Bone Miner. Res. 2014, 29, 408–417. [Google Scholar] [CrossRef]
- Chen, W.; Rezaizadehnajafi, L.; Wink, M. Influence of Resveratrol on Oxidative Stress Resistance and Life Span in Caenorhabditis elegans. J. Pharm. Pharmacol. 2013, 65, 682–688. [Google Scholar] [CrossRef]
- Desjardins, D.; Cacho-Valadez, B.; Liu, J.-L.; Wang, Y.; Yee, C.; Bernard, K.; Khaki, A.; Breton, L.; Hekimi, S. Antioxidants Reveal an Inverted U-Shaped Dose-Response Relationship between Reactive Oxygen Species Levels and the Rate of Aging in Caenorhabditis elegans. Aging Cell 2017, 16, 104–112. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; González-Manzano, S.; Escudero-Gilete, M.L.; Hernanz, D.; Dueñas, M.; González-Paramás, A.M.; Heredia, F.J.; Santos-Buelga, C. Study of Zalema Grape Pomace: Phenolic Composition and Biological Effects in Caenorhabditis elegans. J. Agric. Food Chem. 2013, 61, 5114–5121. [Google Scholar] [CrossRef]
- Khalil, A.; Tazeddinova, D.; Aljoumaa, K.; Kazhmukhanbetkyzy, Z.A.; Orazov, A.; Toshev, A.D. Carotenoids: Therapeutic Strategy in the Battle against Viral Emerging Diseases, COVID-19: An Overview. Prev. Nutr. Food Sci. 2021, 26, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids Functionality, Sources, and Processing by Supercritical Technology: A Review. J. Chem. 2016, 2016, e3164312. [Google Scholar] [CrossRef] [Green Version]
- Ambrósio, C.L.B.; de Campos, F.A.C.; de Faro, Z.P. Carotenóides como alternativa contra a hipovitaminose A. Rev. Nutr. 2006, 19, 233–243. [Google Scholar] [CrossRef]
- Roohbakhsh, A.; Karimi, G.; Iranshahi, M. Carotenoids in the Treatment of Diabetes Mellitus and Its Complications: A Mechanistic Review. Biomed. Pharmacother. 2017, 91, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, Pharmacology and Treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [Green Version]
- Hussain, Y.; Alsharif, K.F.; Aschner, M.; Theyab, A.; Khan, F.; Saso, L.; Khan, H. Therapeutic Role of Carotenoids in Blood Cancer: Mechanistic Insights and Therapeutic Potential. Nutrients 2022, 14, 1949. [Google Scholar] [CrossRef]
- Koklesova, L.; Liskova, A.; Samec, M.; Zhai, K.; Abotaleb, M.; Ashrafizadeh, M.; Brockmueller, A.; Shakibaei, M.; Biringer, K.; Bugos, O.; et al. Carotenoids in Cancer Metastasis—Status Quo and Outlook. Biomolecules 2020, 10, 1653. [Google Scholar] [CrossRef]
- Van Hoang, D.; Pham, N.M.; Lee, A.H.; Tran, D.N.; Binns, C.W. Dietary Carotenoid Intakes and Prostate Cancer Risk: A Case-Control Study from Vietnam. Nutrients 2018, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Ono, A.; Koshiyama, M.; Nakagawa, M.; Watanabe, Y.; Ikuta, E.; Seki, K.; Oowaki, M. The Preventive Effect of Dietary Antioxidants on Cervical Cancer Development. Medicina 2020, 56, 604. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Jang, J.-H.; Lee, S.-Y. An Updated Comprehensive Review on Vitamin A and Carotenoids in Breast Cancer: Mechanisms, Genetics, Assessment, Current Evidence, and Future Clinical Implications. Nutrients 2021, 13, 3162. [Google Scholar] [CrossRef]
- Koklesova, L.; Liskova, A.; Samec, M.; Buhrmann, C.; Samuel, S.M.; Varghese, E.; Ashrafizadeh, M.; Najafi, M.; Shakibaei, M.; Büsselberg, D.; et al. Carotenoids in Cancer Apoptosis—The Road from Bench to Bedside and Back. Cancers 2020, 12, 2425. [Google Scholar] [CrossRef] [PubMed]
- Black, H.S.; Boehm, F.; Edge, R.; Truscott, T.G. The Benefits and Risks of Certain Dietary Carotenoids That Exhibit Both Anti- and Pro-Oxidative Mechanisms—A Comprehensive Review. Antioxidants 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addis, G.; Baskaran, R.; Raju, M.; Ushadevi, A.; Asfaw, Z.; Woldu, Z.; Baskaran, V. Effect of Blanching and Drying Process on Carotenoids Composition of Underutilized Ethiopian (Coccinia grandis L. Voigt) and Indian (Trigonella foenum-graecum L.) Green Leafy Vegetables. J. Food Process. Preserv. 2009, 33, 744–762. [Google Scholar] [CrossRef]
- Xu, C.; Wang, S.; Ti, W.; Yang, J.; Yasen, Y.; Memetsidiq, M.; Shi, S. Role of Dietary Patterns and Factors in Determining the Risk of Knee Osteoarthritis: A Meta-Analysis. Mod. Rheumatol. 2022, 32, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Nakamura, M.; Ogawa, K.; Ikoma, Y.; Ando, F.; Shimokata, H.; Yano, M. Dietary Patterns of Antioxidant Vitamin and Carotenoid Intake Associated with Bone Mineral Density: Findings from Post-Menopausal Japanese Female Subjects. Osteoporos. Int. 2011, 22, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Estévez-Santiago, R.; Olmedilla-Alonso, B.; Fernández-Jalao, I. Bioaccessibility of Provitamin A Carotenoids from Fruits: Application of a Standardised Static in Vitro Digestion Method. Food Funct. 2016, 7, 1354–1366. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Uchiyama, S. Beta-Cryptoxanthin Stimulates Bone Formation and Inhibits Bone Resorption in Tissue Culture in Vitro. Mol. Cell. Biochem. 2004, 258, 137–144. [Google Scholar] [CrossRef]
- Jiang, H.; Yin, Y.; Wu, C.-R.; Liu, Y.; Guo, F.; Li, M.; Ma, L. Dietary Vitamin and Carotenoid Intake and Risk of Age-Related Cataract. Am. J. Clin. Nutr. 2019, 109, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.D.; Karki, S.; Thakur, N.S.; Attri, S. Chemical Composition, Functional Properties and Processing of Carrot—A Review. J. Food Sci. Technol. 2012, 49, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Lem, D.W.; Davey, P.G.; Gierhart, D.L.; Rosen, R.B. A Systematic Review of Carotenoids in the Management of Age-Related Macular Degeneration. Antioxidants 2021, 10, 1255. [Google Scholar] [CrossRef]
- Yao, Y.; Goh, H.M.; Kim, J.E. The Roles of Carotenoid Consumption and Bioavailability in Cardiovascular Health. Antioxidants 2021, 10, 1978. [Google Scholar] [CrossRef]
- Matsumoto, M.; Waki, N.; Suganuma, H.; Takahashi, I.; Kurauchi, S.; Sawada, K.; Tokuda, I.; Misawa, M.; Ando, M.; Itoh, K.; et al. Association between Biomarkers of Cardiovascular Diseases and the Blood Concentration of Carotenoids among the General Population without Apparent Illness. Nutrients 2020, 12, 2310. [Google Scholar] [CrossRef]
- Shivakoti, R.; Gupte, N.; Tripathy, S.; Poongulali, S.; Kanyama, C.; Berendes, S.; Cardoso, S.W.; Santos, B.R.; La Rosa, A.; Mwelase, N.; et al. Inflammation and Micronutrient Biomarkers Predict Clinical HIV Treatment Failure and Incident Active TB in HIV-Infected Adults: A Case-Control Study. BMC Med. 2018, 16, 161. [Google Scholar] [CrossRef]
- Pechinskii, S.V.; Kuregyan, A.G. The Impact of Carotenoids on Immunity (Review) | SpringerLink. Pharm. Chem. J. 2014, 47, 509–513. [Google Scholar] [CrossRef]
- Rowles, J.L.; Erdman, J.W. Carotenoids and Their Role in Cancer Prevention. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158613. [Google Scholar] [CrossRef]
- Thakur, D.; Jain, A.; Ghoshal, G.; Shivhare, U.S.; Katare, O.P. Microencapsulation of β-Carotene Based on Casein/Guar Gum Blend Using Zeta Potential-Yield Stress Phenomenon: An Approach to Enhance Photo-Stability and Retention of Functionality. AAPS PharmSciTech 2017, 18, 1447–1459. [Google Scholar] [CrossRef]
- Delgado-Vargas, F.; Jiménez, A.R.; Paredes-López, O. Natural Pigments: Carotenoids, Anthocyanins, and Betalains—Characteristics, Biosynthesis, Processing, and Stability. Crit. Rev. Food Sci. Nutr. 2000, 40, 173–289. [Google Scholar] [CrossRef]
- Burton, G.W.; Daroszewski, J.; Mogg, T.J.; Nikiforov, G.B.; Nickerson, J.G. Discovery and Characterization of Carotenoid-Oxygen Copolymers in Fruits and Vegetables with Potential Health Benefits. J. Agric. Food Chem. 2016, 64, 3767–3777. [Google Scholar] [CrossRef] [Green Version]
- Carail, M.; Caris-Veyrat, C. Carotenoid Oxidation Products: From Villain to Saviour? Pure Appl. Chem. 2006, 78, 1493–1503. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.B.; Nickerson, J.G.; Daroszewski, J.; Mogg, T.J.; Burton, G.W. Biologically Active Polymers from Spontaneous Carotenoid Oxidation: A New Frontier in Carotenoid Activity. PLoS ONE 2014, 9, e111346. [Google Scholar] [CrossRef] [PubMed]
- Soukoulis, C.; Bohn, T. A Comprehensive Overview on the Micro- and Nano-Technological Encapsulation Advances for Enhancing the Chemical Stability and Bioavailability of Carotenoids. Crit. Rev. Food Sci. Nutr. 2018, 58, 1–36. [Google Scholar] [CrossRef]
- McClements, D.J. Food Emulsions: Principles, Practices, and Techniques, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 978-0-429-15403-4. [Google Scholar]
- Walstra, P.; van Vliet, T. Dispersed Systems: Basic Considerations. In Fennema’s Food Chemistry; Damodaran, S., Parkin, K.L., Fennema, O.R., Eds.; Marcel Dekker: New York, NY, USA, 2007; pp. 783–847. ISBN 978-0-8493-9272-6. [Google Scholar]
- Focsan, A.L.; Polyakov, N.E.; Kispert, L.D. Supramolecular Carotenoid Complexes of Enhanced Solubility and Stability—The Way of Bioavailability Improvement. Molecules 2019, 24, 3947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janiszewska-Turak, E. Carotenoids Microencapsulation by Spray Drying Method and Supercritical Micronization. Food Res. Int. 2017, 99, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Capelezzo, A.P.; Mohr, L.C.; Dalcanton, F.; de Mello, J.M.M.; Fiori, M.A. β-Cyclodextrins as Encapsulating Agents of Essential Oils. In Cyclodextrin. A versatile Ingdredient; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Donhowe, E.G.; Kong, F. Beta-Carotene: Digestion, Microencapsulation, and in Vitro Bioavailability. Food Bioprocess Technol. 2014, 7, 338–354. [Google Scholar] [CrossRef]
- Arellano Corral, C.A. Extracción de Luteína a Partir de Flores de Tagete (Tagete erecta) y Estabilización por Microencapsulación. Ph.D. Thesis, Facultad de Ciencias Químicas y Farmacéuticas, Universidad de Chile, Santiago, Chile, 2011. [Google Scholar]
- Mestry, A.P.; Mujumdar, A.S.; Thorat, B.N. Optimization of Spray Drying of an Innovative Functional Food: Fermented Mixed Juice of Carrot and Watermelon. Dry. Technol. 2011, 29, 1121–1131. [Google Scholar] [CrossRef]
- Apanasenko, I.E.; Selyutina, O.Y.; Polyakov, N.E.; Suntsova, L.P.; Meteleva, E.S.; Dushkin, A.V.; Vachali, P.; Bernstein, P.S. Solubilization and Stabilization of Macular Carotenoids by Water Soluble Oligosaccharides and Polysaccharides. Arch. Biochem. Biophys. 2015, 572, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, I.L.; Mercadante, A.Z. Encapsulation of Lycopene Using Spray-Drying and Molecular Inclusion Processes. Braz. Arch. Biol. Technol. 2007, 50, 893–900. [Google Scholar] [CrossRef]
- Juscamaita Fabián, L.; Pérez Salvatierra, T.; Espinoza Silva, C.; Quispe Solano, M.; Hinostroza Quiñónez, G.; Flores Ramos, O.; Manyari Cervantes, G. Evaluación de la Estabilidad de Carotenoides y Actividad Antioxidante de la Flor de Mastuerzo (Tropaeolum majus L.) en la Microencapsulación por Spray-Drying. Rev. Soc. Quím. Perú 2017, 83, 282–293. [Google Scholar] [CrossRef]
- Lim, A.S.L.; Roos, Y.H. Spray Drying of High Hydrophilic Solids Emulsions with Layered Interface and Trehalose-Maltodextrin as Glass Formers for Carotenoids Stabilization. J. Food Eng. 2016, 171, 174–184. [Google Scholar] [CrossRef]
- De Marco, R.; Vieria, A.M.S.; Ugri, M.A.; Monteiro, A.; Bergamasco, R. Microencapsulation of Annatto Seed Extract: Stability and Application. Chem. Eng. Trans. 2013, 32, 1777–1782. [Google Scholar] [CrossRef]
- Przybysz, M.; Szterk, A.; Symoniuk, E.; Gąszczyk, M.; Dłużewska, E. α- and β-Carotene Stability during Storage of Microspheres Obtained from Spray-Dried Microencapsulation Technology. Pol. J. Food Nutr. Sci. 2018, 68, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Hojjati, M.; Razavi, S.H.; Rezaei, K.; Gilani, K. Stabilization of Canthaxanthin Produced by Dietzia natronolimnaea HS-1 with Spray Drying Microencapsulation. J. Food Sci. Technol. 2014, 51, 2134–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, E.; Mariutti, L.R.B.; Chisté, R.C.; Mercadante, A.Z. Development of a Novel Micro-Assay for Evaluation of Peroxyl Radical Scavenger Capacity: Application to Carotenoids and Structure—Activity Relationship. Food Chem. 2012, 135, 2103–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aissa, A.F.; Bianchi, M.L.P.; Ribeiro, J.C.; Hernandes, L.C.; de Faria, A.F.; Mercadante, A.Z.; Antunes, L.M.G. Comparative Study of β-Carotene and Microencapsulated β-Carotene: Evaluation of Their Genotoxic and Antigenotoxic Effects. Food Chem. Toxicol. 2012, 50, 1418–1424. [Google Scholar] [CrossRef] [PubMed]
- Corrêa-Filho, L.C.; Lourenço, S.C.; Duarte, D.F.; Moldão-Martins, M.; Alves, V.D. Microencapsulation of Tomato (Solanum lycopersicum L.) Pomace Ethanolic Extract by Spray Drying: Optimization of Process Conditions. Appl. Sci. 2019, 9, 612. [Google Scholar] [CrossRef] [Green Version]
- Corrêa-Filho, L.C.; Lourenço, M.M.; Moldão-Martins, M.; Alves, V.D. Microencapsulation of β-Carotene by Spray Drying: Effect of Wall Material Concentration and Drying Inlet Temperature. Int. J. Food Sci. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Faria, A.F.; Mignone, R.A.; Montenegro, M.A.; Mercadante, A.Z.; Borsarelli, C.D. Characterization and Singlet Oxygen Quenching Capacity of Spray-Dried Microcapsules of Edible Biopolymers Containing Antioxidant Molecules. J. Agric. Food Chem. 2010, 58, 8004–8011. [Google Scholar] [CrossRef] [Green Version]
- Boiero, M.L.; Mandrioli, M.; Braber, N.V.; Rodriguez-Estrada, M.T.; García, N.A.; Borsarelli, C.D.; Montenegro, M.A. Gum Arabic Microcapsules as Protectors of the Photoinduced Degradation of Riboflavin in Whole Milk. J. Dairy Sci. 2014, 97, 5328–5336. [Google Scholar] [CrossRef] [Green Version]
- Weiss, J.; Decker, E.A.; McClements, D.J.; Kristbergsson, K.; Helgason, T.; Awad, T. Solid Lipid Nanoparticles as Delivery Systems for Bioactive Food Components. Food Biophys. 2008, 3, 146–154. [Google Scholar] [CrossRef]
- Sessa, M. Nanoencapsulation of Bioactive Compounds for Food Applications; Universitá Degli Studi di Salerno: Fisciano, Italy, 2012. [Google Scholar]
- Rehman, A.; Tong, Q.; Jafari, S.M.; Assadpour, E.; Shehzad, Q.; Aadil, R.M.; Iqbal, M.W.; Rashed, M.M.A.; Mushtaq, B.S.; Ashraf, W. Carotenoid-Loaded Nanocarriers: A Comprehensive Review. Adv. Colloid Interface Sci. 2020, 275, 102048. [Google Scholar] [CrossRef]
- Dos Santos, P.P.; de Andrade, L.A.; Flôres, S.H.; de Rios, A.O. Nanoencapsulation of Carotenoids: A Focus on Different Delivery Systems and Evaluation Parameters. J. Food Sci. Technol. 2018, 55, 3851–3860. [Google Scholar] [CrossRef]
- Tasch Holkem, A.; Franco Codevilla, C.; De Bona da Silva, C.; Ragagnin de Menezes, C. Técnicas de preparação de sistemas nanotecnológicos aplicados a alimentos. Ciênc. Nat. 2015, 37, 87. [Google Scholar] [CrossRef]
- Tamjidi, F.; Shahedi, M.; Varshosaz, J.; Nasirpour, A. Stability of Astaxanthin-Loaded Nanostructured Lipid Carriers as Affected by PH, Ionic Strength, Heat Treatment, Simulated Gastric Juice and Freeze–Thawing. J. Food Sci. Technol. 2017, 54, 3132–3141. [Google Scholar] [CrossRef] [PubMed]
- Soh, S.H.; Lee, L.Y. Microencapsulation and Nanoencapsulation Using Supercritical Fluid (SCF) Techniques. Pharmaceutics 2019, 11, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocero, M.J.; Martín, Á.; Mattea, F.; Varona, S. Encapsulation and Co-Precipitation Processes with Supercritical Fluids: Fundamentals and Applications. J. Supercrit. Fluids 2009, 47, 546–555. [Google Scholar] [CrossRef]
- Silva, E.K.; Meireles, A.A. Encapsulation of Food Compounds Using Supercritical Technologies: Applications of Supercritical Carbon Dioxide as an Antisolvent. Food Public Health 2014, 4, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Xia, Z.; McClements, D.J.; Xiao, H. Influence of Lipid Content in a Corn Oil Preparation on the Bioaccessibility of β-Carotene: A Comparison of Low-Fat and High-Fat Samples: Influence of Lipid Conten. J. Food Sci. 2017, 82, 373–379. [Google Scholar] [CrossRef]
- Zhao, L.; Temelli, F.; Curtis, J.M.; Chen, L. Encapsulation of Lutein in Liposomes Using Supercritical Carbon Dioxide. Food Res. Int. 2017, 100, 168–179. [Google Scholar] [CrossRef]
- Machado, F.R.S., Jr.; Reis, D.F.; Boschetto, D.L.; Burkert, J.F.M.; Ferreira, S.R.S.; Oliveira, J.V.; Burkert, C.A.V. Encapsulation of Astaxanthin from Haematococcus Pluvialis in PHBV by Means of SEDS Technique Using Supercritical CO2. Ind. Crops Prod. 2014, 54, 17–21. [Google Scholar] [CrossRef]
- Zhong, J.; Dai, L.C. Liposomal Preparation by Supercritical Fluids Technology. Afr. J. Biotechnol. 2011, 10, 16406–16413. [Google Scholar] [CrossRef]
- Quan, C.; Johan, C.; Charlotta, T. Carotenoids Particle Formation by Supercritical Fluid Technologies. Chin. J. Chem. Eng. 2009, 17, 344–349. [Google Scholar] [CrossRef]
- de Paz, E.; Martín, Á.; Duarte, C.M.M.; Cocero, M.J. Formulation of β-Carotene with Poly-(ε-Caprolactones) by PGSS Process. Powder Technol. 2012, 217, 77–83. [Google Scholar] [CrossRef]
- de Paz, E.; Martín, Á.; Cocero, M.J. Formulation of β-Carotene with Soybean Lecithin by PGSS (Particles from Gas Saturated Solutions)-Drying. J. Supercrit. Fluids 2012, 72, 125–133. [Google Scholar] [CrossRef]
- Santos, D.T.; Martín, Á.; Meireles, M.A.A.; Cocero, M.J. Production of Stabilized Sub-Micrometric Particles of Carotenoids Using Supercritical Fluid Extraction of Emulsions. J. Supercrit. Fluids 2012, 61, 167–174. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Carotenoids | Food Additive | Color | Approved Use In | Food Application [14,31] | |
|---|---|---|---|---|---|
| EU [31] | USA [32] | ||||
| Carotenes or β-carotene | E160a | Yellow to Orange | Yes | Yes | Soft drinks, juice, butter, preserves of red fruits, vegetables in vinegar or brine, jam, chesses, candies, breakfast cereals, fats, sausages, pates, bakery products, precooked and smoked fish. |
| Carrot oil | Yellow to Orange | NR | Yes | Food generally. | |
| Bixin and norbixin from/or Annatto extract | E160b | Orange to Brown Red | Yes | Yes | Dairy products and fermented milk, butter, fats, breakfast cereals, ice cream, desserts, custards, candies, snacks, cheeses, smoked fish, alcoholic beverages and sausages. |
| Capsanthin and capsorubin from/or Paprika oleoresin | E160c | Red | Yes | Yes | Breakfast cereals, cheeses, creams, sausages, surimi, preserves of red fruits, instant soups, snacks, smoked fish, pates, jams, jellies and marmalades. |
| Lycopene or tomato extract or tomato concentrate | E160d | Bright to Deep Red | Yes | Yes | Sauces, fermented milk products, edible ices, seafood, snacks, desserts, condiments, dietary supplements, meat substitutes, coating, fillings and decorations of bakery products, soups, chesses and flavored drinks. |
| β-Apo-8′-carotenal | E160e | Orange Red to Yellow | Yes | Yes | Orange and lemon soft drinks, juice, nectars, shakes, cheese, jams, jellies, marmalades and precooked food. |
| Ethyl ester of β-Apo-8′-carotenic acid | E160f | Orange Red to Yellow | Yes | No | Orange and lemon soft drinks, juice, nectars, shakes, margarines, butter, pies, cereals, and precooked food. |
| Flavoxanthin | E161a | Yellow | No | No | Candies, yoghurts and dairy products. |
| Lutein | E161b | Orange Red to Yellow | Yes | Yes a | Jams, instant soups, creams, yoghurts, cheese, seafood, soft drinks, alcoholic beverages and poultry feed. |
| Cryptoxanthin | E161c | Orange to Red | No | No | Confectionary. |
| Rubixanthin | E161d | Orange to Red | No | No | Confectionary. |
| Violaxanthin | E161e | Orange | No | No | Confectionary. |
| Rhodoxanthin | E161f | Yellow | No | No | Confectionary and ice cream. |
| Canthaxanthin | E161g | Orange | No | Yes a | Fish and poultry feed. |
| Zeaxanthin | E161h | Orange to Red | No | No | Confectionary. |
| Citranaxanthin | E161i | Yellow | No | No | |
| Astaxanthin | E161j | Red | Yes a | Yes a | Fish and poultry feed. |
| Saffron (Crocin) | E164 | Gold Yellow to Orange | NR | Yes | Rice dishes, sausages, margarine, butter, cheese, ice cream, alcoholic and non-alcoholic beverages. |
| Age | Recommended Dietary Allowances (µg RAE/Day) | |
|---|---|---|
| Male | Female | |
| 0 to 6 months | 400 | 400 |
| 7 to 12 months | 500 | 500 |
| 1 to 3 years | 300 | 300 |
| 4 to 6 years | 400 | 400 |
| 9 to 13 years | 600 | 600 |
| 14 to 18 years | 900 | 700 |
| 750 a | ||
| 1200 b | ||
| 19 to 50 years | 900 | 700 |
| 770 a | ||
| 51+ years | 900 | 700 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Peña, M.A.; Ortega-Regules, A.E.; Anaya de Parrodi, C.; Lozada-Ramírez, J.D. Chemistry, Occurrence, Properties, Applications, and Encapsulation of Carotenoids—A Review. Plants 2023, 12, 313. https://doi.org/10.3390/plants12020313
González-Peña MA, Ortega-Regules AE, Anaya de Parrodi C, Lozada-Ramírez JD. Chemistry, Occurrence, Properties, Applications, and Encapsulation of Carotenoids—A Review. Plants. 2023; 12(2):313. https://doi.org/10.3390/plants12020313
Chicago/Turabian StyleGonzález-Peña, Marco Antonio, Ana Eugenia Ortega-Regules, Cecilia Anaya de Parrodi, and José Daniel Lozada-Ramírez. 2023. "Chemistry, Occurrence, Properties, Applications, and Encapsulation of Carotenoids—A Review" Plants 12, no. 2: 313. https://doi.org/10.3390/plants12020313