
Transcriptome Analysis Reveals Changes in Whole Gene Expression, Biological Process, and Molecular Functions Induced by Nickel in Jack Pine (Pinus banksiana)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Treatment
2.2. RNA Extraction
2.3. RNA Sequencing and De Novo Transcriptome Assembly
2.4. BLAT Matching and Annotation of Pinus banksiana Genes
2.5. Quantification of Gene Expression and Quality Control (QC) Analysis
2.6. Differential Gene Expression (DGE) Analysis of Pairwise Comparisons
2.7. Analysis of Top Differentially Regulated Genes
3. Results
3.1. Transcript Assembly and Sequence Data QC
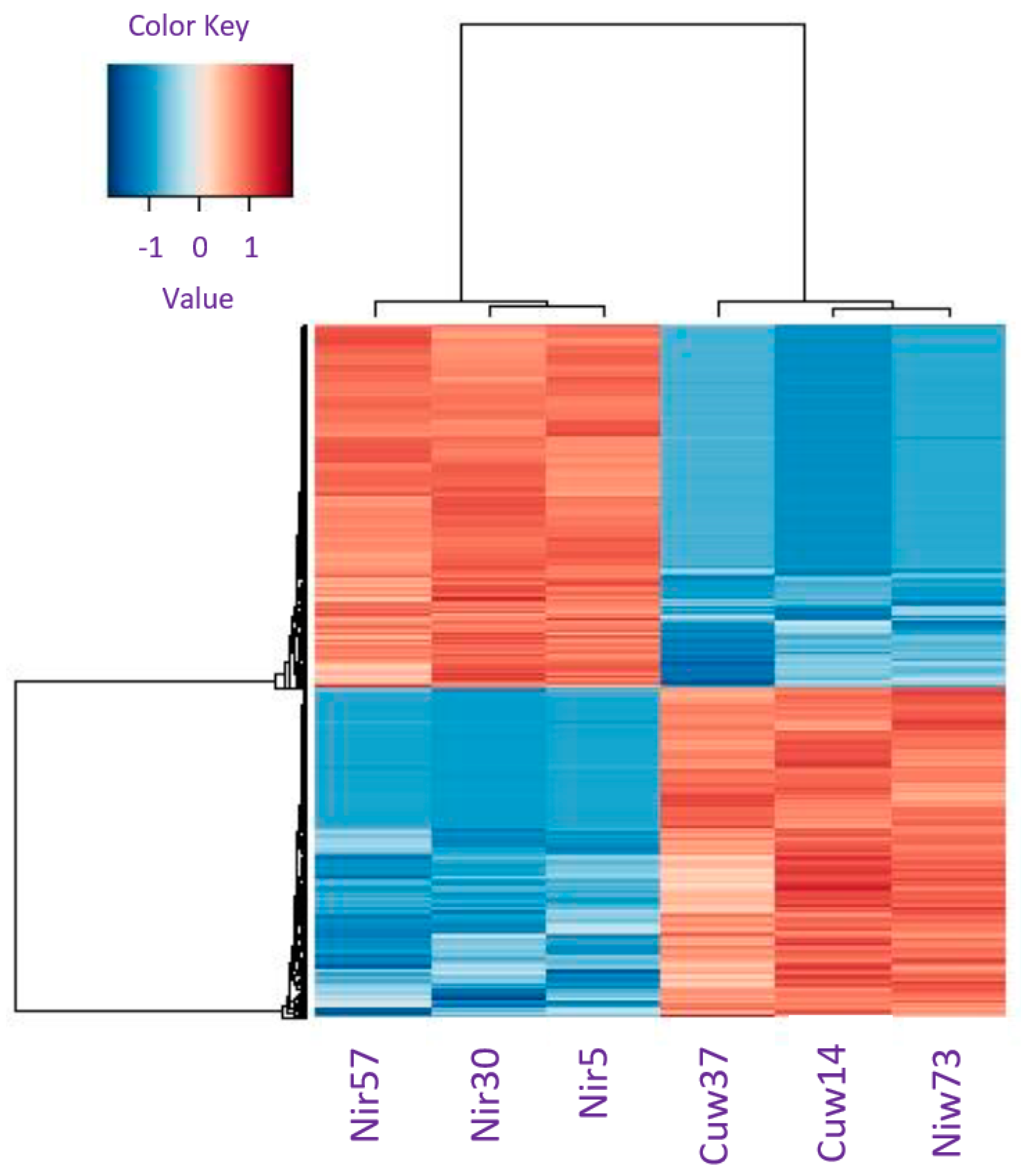
3.2. Differential Gene Expression (DGE) Analysis
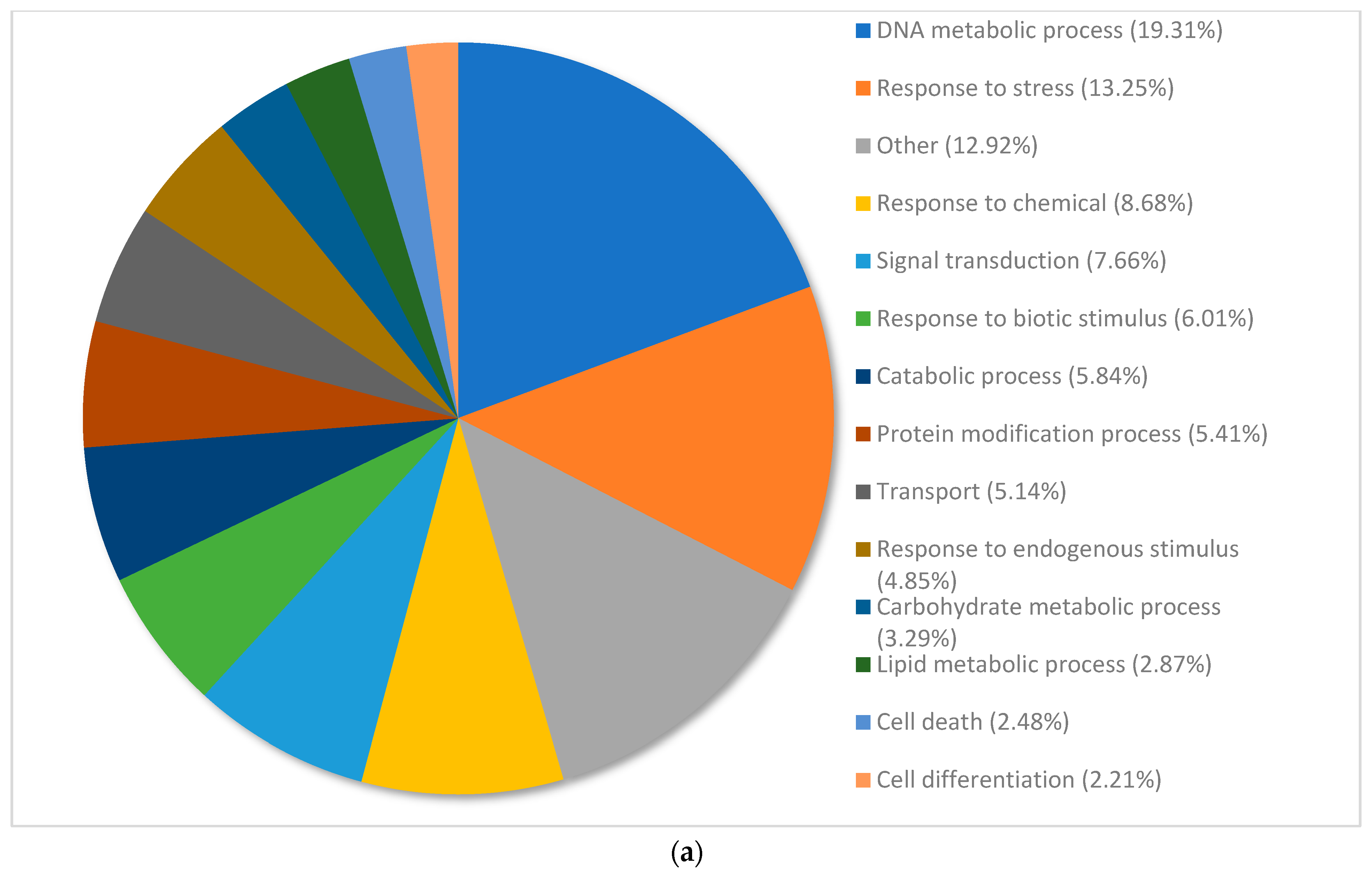
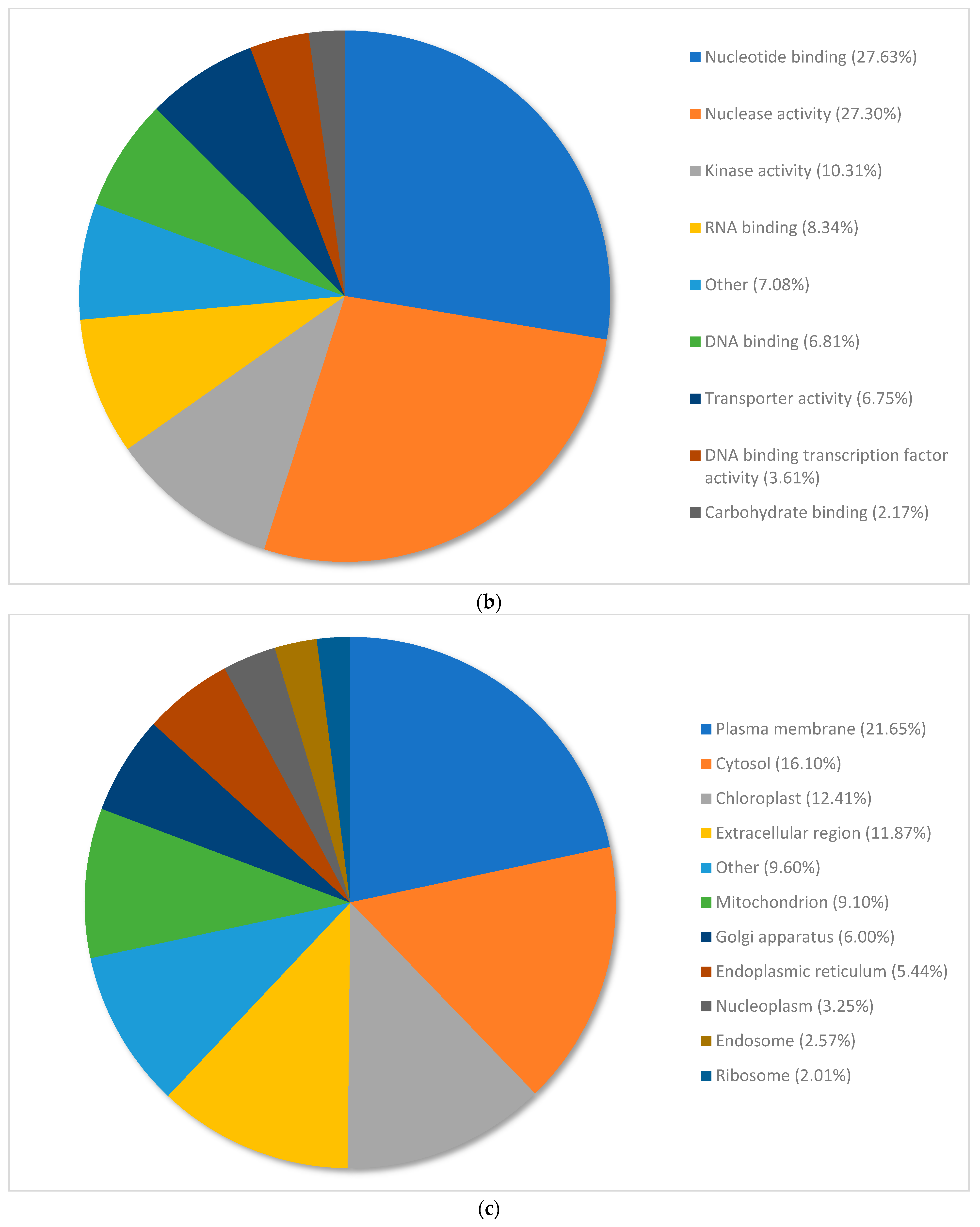
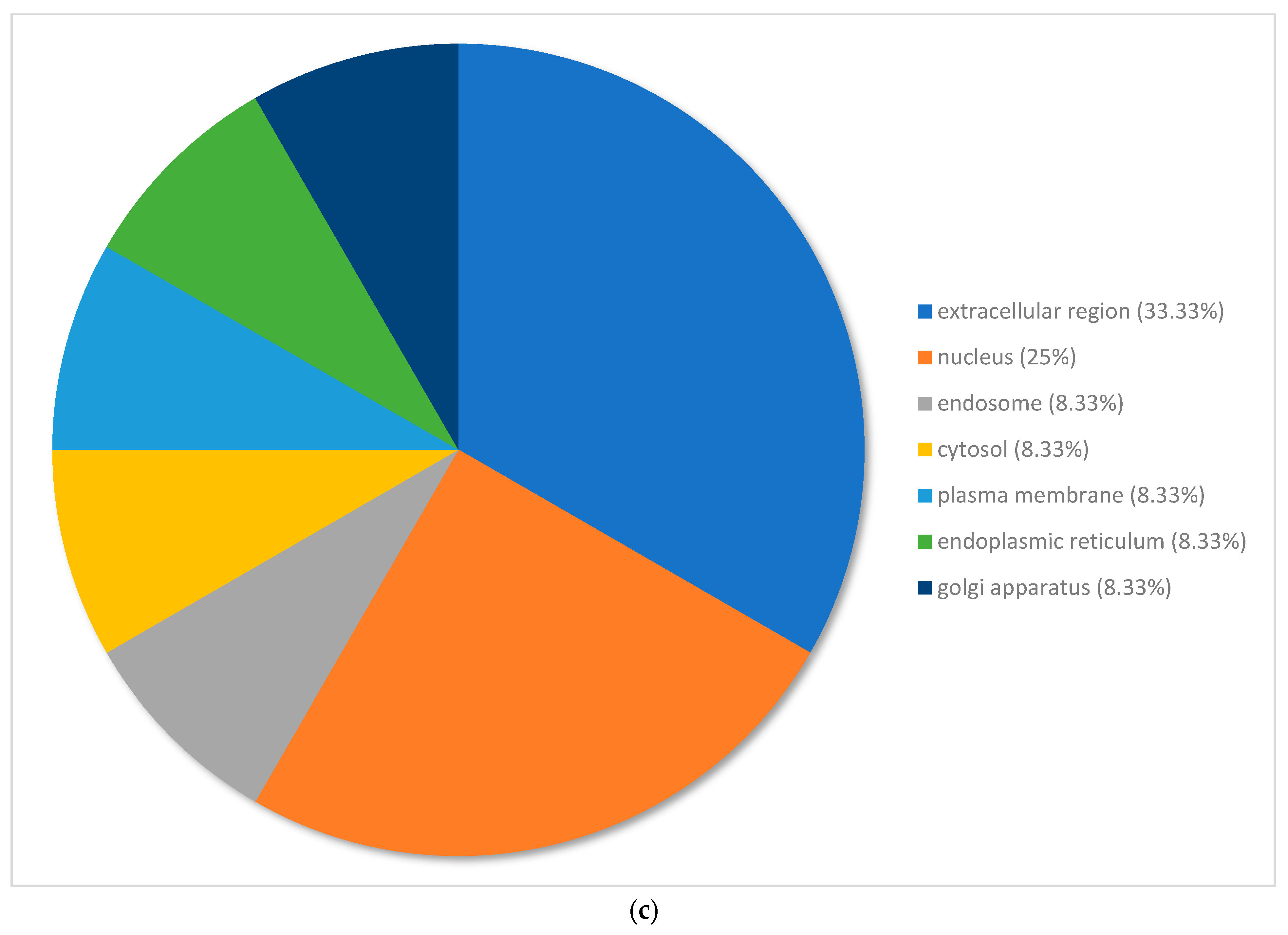
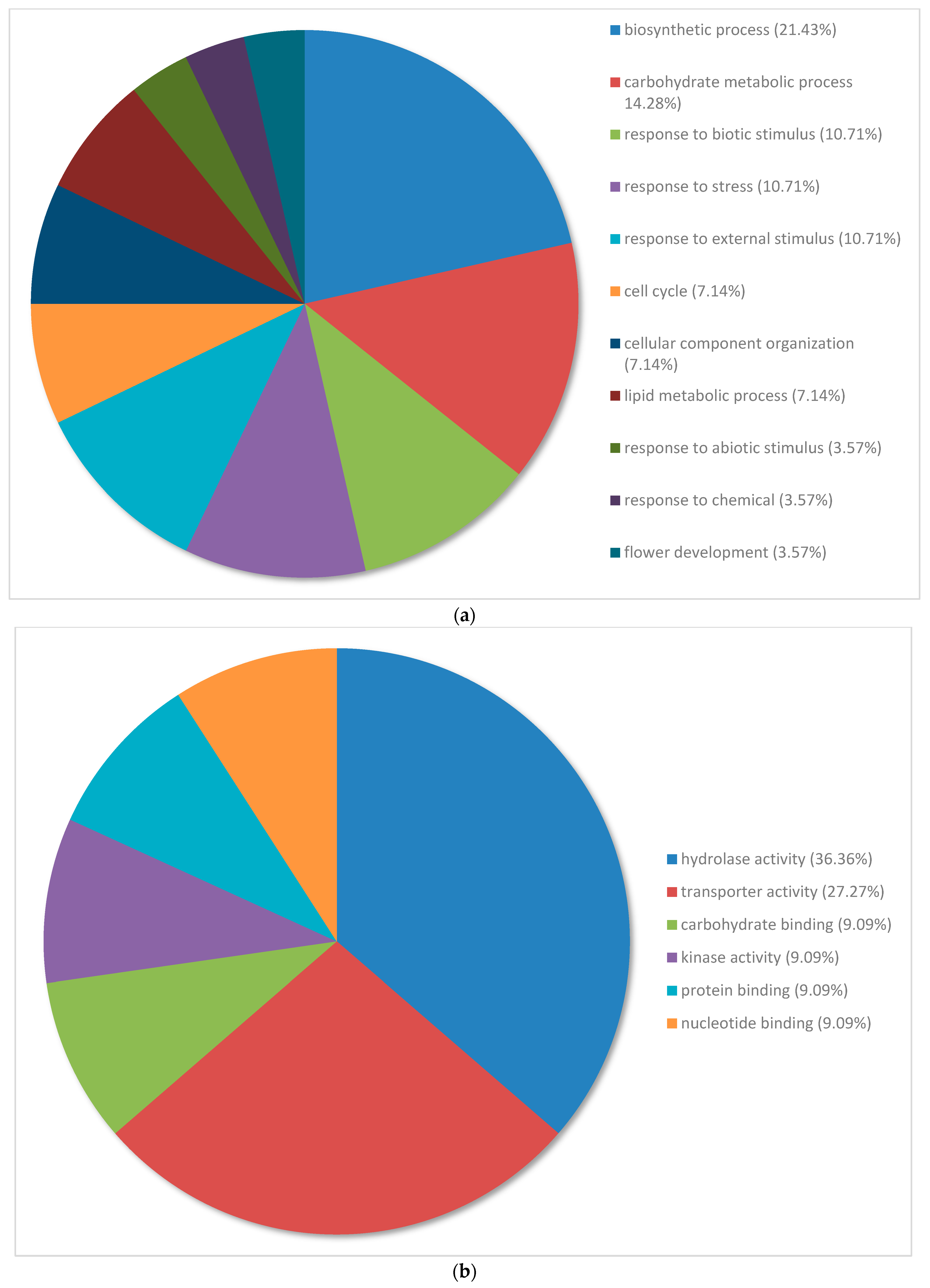
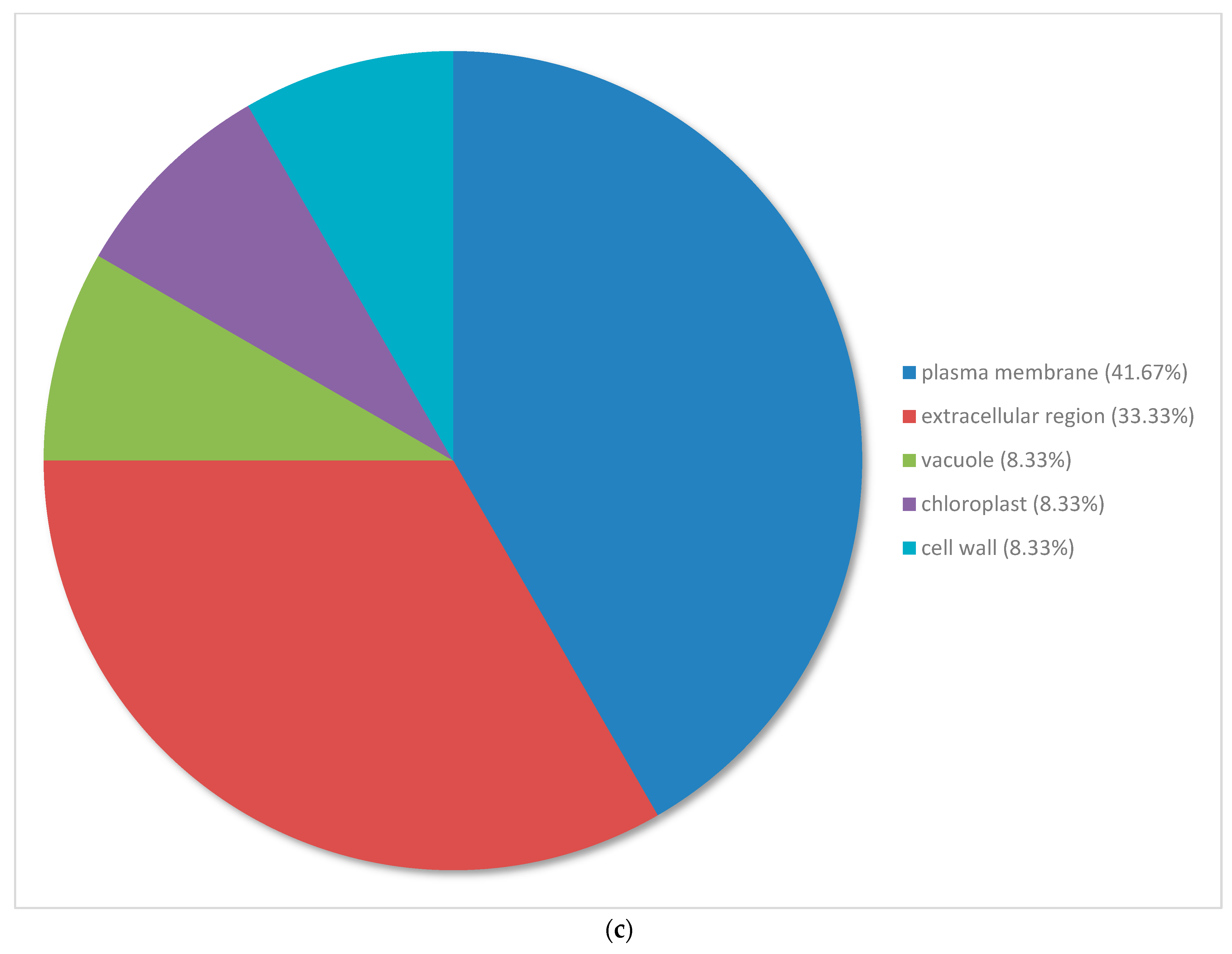
3.3. Gene Ontology Classification of Differentially Expressed Genes in Pinus banksiana
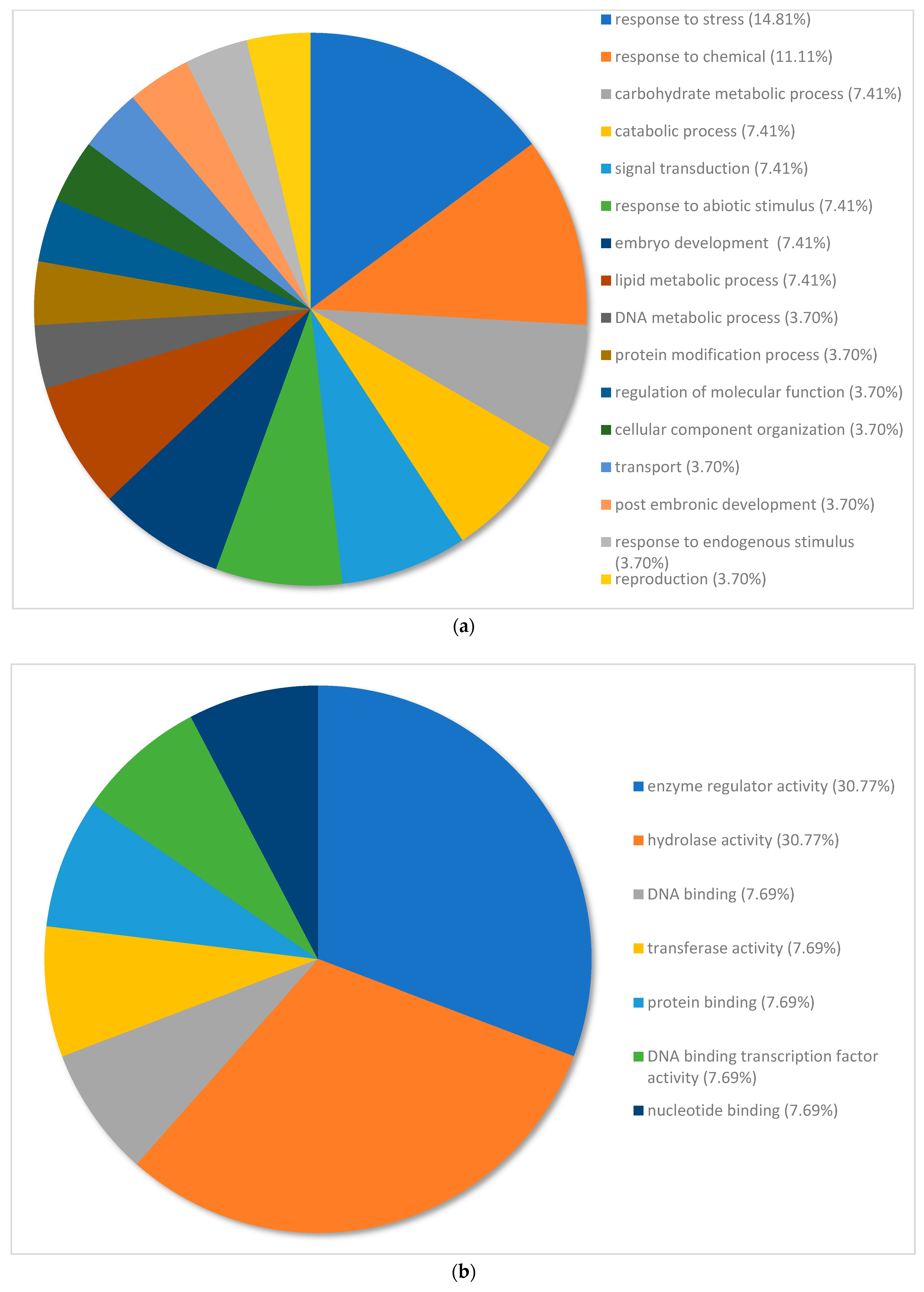
3.4. Gene Ontology of the Top 100 Differentially Expressed Genes in Pinus banksiana
3.5. Top Differentially Expressed Genes for Pairwise Comparisons
4. Discussion
4.1. Differential Gene Expression (DEG) Analysis
4.2. Gene Ontology of the Top 100 DEGs in Response to Excess Nickel
4.3. Annotation of the Top Upregulated Genes between Nickel-Treated Plants and the Control
4.4. Annotation of the Top Downregulated Genes between Nickel-Treated Plants and the Control
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schindler, M. A mineralogical and geochemical study of slag from the historical O’donnell roast yards, Sudbury, Ontario, Canada. Can. Mineral. 2014, 52, 433–452. [Google Scholar] [CrossRef]
- Jewiss, T. The Mining History of the Sudbury Area|Earth Sciences Museum. Available online: https://uwaterloo.ca/earth-sciences-museum/resources/mining-canada/mining-history-sudbury-area (accessed on 1 January 2022).
- Keller, W.; Yan, N.D.; Gunn, J.M.; Heneberry, J. Recovery of Acidified Lakes: Lessons From Sudbury, Ontario, Canada. In Acid Rain-Deposition to Recovery; Brimblecombe, P., Hara, H., Houle, D., Novak, M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 317–322. ISBN 978-1-4020-5884-4. [Google Scholar]
- Xu, J.; Yang, L.; Wang, Z.; Dong, G.; Huang, J.; Wang, Y. Toxicity of Copper on Rice Growth and Accumulation of Copper in Rice Grain in Copper Contaminated Soil. Chemosphere 2006, 62, 602–607. [Google Scholar] [CrossRef]
- Baccouch, S.; Chaoui, A.; El Ferjani, E. Nickel toxicity induces oxidative damage in Zea mays roots. J. Plant Nutr. 2001, 24, 1085–1097. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Hayat, S.; Ahmad, A. Nickel: An Overview of Uptake, Essentiality and Toxicity in Plants. Bull. Environ. Contam. Toxicol. 2011, 86, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, S.; Joly, D.; Leclerc, S.; Govindachary, S.; Harnois, J.; Carpentier, R. Inhibition of the Oxygen-Evolving Complex of Photosystem II and Depletion of Extrinsic Polypeptides by Nickel. BioMetals 2007, 20, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Küpper, H.; Küpper, F.; Spiller, M. Environmental Relevance of Heavy Metal-Substituted Chlorophylls Using the Example of Water Plants. J. Exp. Bot. 1996, 47, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Batool, S. Effect of Nickel Toxicity on Growth, Photosynthetic Pigments and Dry Matter Yield of Cicer arietinum L. Varieties. Asian J. Agric. Biol. 2018, 6, 143–148. [Google Scholar]
- Baran, U.; Ekmekçi, Y. Correction to: Physiological, Photochemical, and Antioxidant Responses of Wild and Cultivated Carthamus Species Exposed to Nickel Toxicity and Evaluation of Their Usage Potential in Phytoremediation. Environ. Sci. Pollut. Res. 2021, 28, 61869–61870. [Google Scholar] [CrossRef]
- Ghasemi, R.; Ghaderian, S.M.; Krämer, U. Interference of Nickel with Copper and Iron Homeostasis Contributes to Metal Toxicity Symptoms in the Nickel Hyperaccumulator Plant Alyssum inflatum. New Phytol. 2009, 184, 566–580. [Google Scholar] [CrossRef]
- Rubio, M.I.; Escrig, I.; Martínez-Cortina, C.; López-Benet, F.J.; Sanz, A. Cadmium and Nickel Accumulation in Rice Plants. Effects on Mineral Nutrition and Possible Interactions of Abscisic and Gibberellic Acids. Plant Growth Regul. 1994, 14, 151–157. [Google Scholar] [CrossRef]
- Yang, X.; Baligar, V.C.; Martens, D.C.; Clark, R.B. Plant Tolerance to Nickel Toxicity: II Nickel Effects on Influx and Transport of Mineral Nutrients in Four Plant Species. J. Plant Nutr. 1996, 19, 265–279. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M. Effect of Nickel on ROS Content and Antioxidative Enzyme Activities in Wheat Leaves. BioMetals 2006, 20, 27–36. [Google Scholar] [CrossRef]
- Pavlova, D. Nickel Effect on Root-Meristem Cell Division in Plantago lanceolata (Plantaginaceae) Seedlings. Aust. J. Bot. 2017, 65, 446. [Google Scholar] [CrossRef]
- Yadav, S.S.; Shukla, R.; Sharma, Y.K. Yadav Nickel Toxicity on Seed Germination and Growth in Radish (Raphanus Sativus) and Its Recovery Using Copper and Boron. J. Environ. Biol. 2022, 30, 461–466. [Google Scholar]
- Ranger, M.; Nkongolo, K.K.; Michael, P.; Beckett, P. Genetic Differentiation of Jack Pine (Pinus banksiana) and Red Pine (P. resinosa) Populations From Metal Contaminated Areas in Northern Ontario (Canada) Using ISSR Markers. Silvae Genet. 2008, 57, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Mari, S.; Gendre, D.; Pianelli, K.; Ouerdane, L.; Lobinski, R.; Briat, J.-F.; Lebrun, M.; Czernic, P. Root-to-Shoot Long-Distance Circulation of Nicotianamine and Nicotianamine-Nickel Chelates in the Metal Hyperaccumulator Thlaspi caerulescens. J. Exp. Bot. 2006, 57, 4111–4122. [Google Scholar] [CrossRef] [Green Version]
- Ingle, R.A.; Mugford, S.T.; Rees, J.D.; Campbell, M.M.; Smith, J.A.C. Constitutively High Expression of the Histidine Biosynthetic Pathway Contributes to Nickel Tolerance in Hyperaccumulator Plants. Plant Cell 2005, 17, 2089–2106. [Google Scholar] [CrossRef] [Green Version]
- Merlot, S.; Hannibal, L.; Martins, S.; Martinelli, L.; Amir, H.; Lebrun, M.; Thomine, S. The Metal Transporter PgIREG1 from the Hyperaccumulator Psychotria Gabriellae Is a Candidate Gene for Nickel Tolerance and Accumulation. J. Exp. Bot. 2014, 65, 1551–1564. [Google Scholar] [CrossRef] [Green Version]
- Nishida, S.; Tanikawa, R.; Ishida, S.; Yoshida, J.; Mizuno, T.; Nakanishi, H.; Furuta, N. Elevated Expression of Vacuolar Nickel Transporter Gene IREG2 Is Associated With Reduced Root-to-Shoot Nickel Translocation in Noccaea japonica. Front. Plant Sci. 2020, 11, 610. [Google Scholar] [CrossRef]
- Moarefi, N.; Nkongolo, K.K. Contrasting Tolerance and Gene Expression between White Pine (Pinus strobus) and Jack Pine (P. banksiana) Exposed to an Increasing Nickel Concentration. Ecol. Genet. Genom. 2022, 24, 100124. [Google Scholar] [CrossRef]
- Boyd, M.; Nkongolo, K. Expression of Genes Associated with Nickel Resistance in White Spruce (Picea glauca) under Nickel Stress: Analysis of AT2G16800 and NRAMP Genes. Am. J. Plant Sci. 2020, 11, 1163–1174. [Google Scholar] [CrossRef]
- Wei, W.; Chai, T.; Zhang, Y.; Han, L.; Xu, J.; Guan, Z. The Thlaspi Caerulescens NRAMP Homologue TcNRAMP3 Is Capable of Divalent Cation Transport. Mol. Biotechnol. 2009, 41, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Bastow, E.L.; Garcia de la Torre, V.S.; Maclean, A.E.; Green, R.T.; Merlot, S.; Thomine, S.; Balk, J. Vacuolar Iron Stores Gated by NRAMP3 and NRAMP4 Are the Primary Source of Iron in Germinating Seeds. Plant Physiol. 2018, 177, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkongolo, K.K.; Spiers, G.; Beckett, P.; Narendrula, R.; Theriault, G.; Tran, A.; Kalubi, K.N. Long-Term Effects of Liming on Soil Chemistry in Stable and Eroded Upland Areas in a Mining Region. Water. Air. Soil Pollut. 2013, 224, 1618. [Google Scholar] [CrossRef]
- Vandeligt, K.K.; Nkongolo, K.K.; Mehes, M.; Beckett, P. Genetic Analysis of Pinus Banksiana and Pinus Resinosa Populations from Stressed Sites Contaminated with Metals in Northern Ontario (Canada). Chem. Ecol. 2011, 27, 369–380. [Google Scholar] [CrossRef]
- Gratton, W.S.; Nkongolo, K.K.; Spiers, G.A. Heavy Metal Accumulation in Soil and Jack Pine (Pinus banksiana) Needles in Sudbury, Ontario, Canada. Bull. Environ. Contam. Toxicol. 2000, 64, 550–557. [Google Scholar] [CrossRef]
- Theriault, G.; Michael, P.; Nkongolo, K. Comprehensive Transcriptome Analysis of Response to Nickel Stress in White Birch (Betula papyrifera). PLoS ONE 2016, 11, e0153762. [Google Scholar] [CrossRef] [Green Version]
- Czajka, K.M.; Nkongolo, K. Transcriptome Analysis of Trembling Aspen (Populus tremuloides) under Nickel Stress. PLoS ONE 2022, 17, e0274740. [Google Scholar] [CrossRef]
- Nkongolo, K.; Theriault, G.; Michael, P. Differential Levels of Gene Expression and Molecular Mechanisms between Red Maple (Acer Rubrum) Genotypes Resistant and Susceptible to Nickel Toxicity Revealed by Transcriptome Analysis. Ecol. Evol. 2018, 8, 4876–4890. [Google Scholar] [CrossRef]
- Rizwan, M.; Imtiaz, M.; Dai, Z.; Mehmood, S.; Adeel, M.; Liu, J.; Tu, S. Nickel Stressed Responses of Rice in Ni Subcellular Distribution, Antioxidant Production, and Osmolyte Accumulation. Environ. Sci. Pollut. Res. 2017, 24, 20587–20598. [Google Scholar] [CrossRef]
- Llamas, A.; Ullrich, C.I.; Sanz, A. Ni2+ Toxicity in Rice: Effect on Membrane Functionality and Plant Water Content. Plant Physiol. Biochem. 2008, 46, 905–910. [Google Scholar] [CrossRef]
- Dos Reis, A.R.; de Queiroz Barcelos, J.P.; de Souza Osório, C.R.W.; Santos, E.F.; Lisboa, L.A.M.; Santini, J.M.K.; dos Santos, M.J.D.; Furlani Junior, E.; Campos, M.; de Figueiredo, P.A.M.; et al. A Glimpse into the Physiological, Biochemical and Nutritional Status of Soybean Plants under Ni-Stress Conditions. Environ. Exp. Bot. 2017, 144, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Çelik, Ö.; Akdaş, E.Y. Tissue-Specific Transcriptional Regulation of Seven Heavy Metal Stress-Responsive MiRNAs and Their Putative Targets in Nickel Indicator Castor Bean (R. communis L.) Plants. Ecotoxicol. Environ. Saf. 2019, 170, 682–690. [Google Scholar] [CrossRef]
- Wang, R.; Wang, J.; Zhao, L.; Yang, S.; Song, Y. Impact of Heavy Metal Stresses on the Growth and Auxin Homeostasis of Arabidopsis Seedlings. BioMetals 2015, 28, 123–132. [Google Scholar] [CrossRef]
- Gantayat, S.; Mania, S.; Pradhan, C.; Das, A.B. Ionic Stress Induced Cytotoxic Effect of Cadmium and Nickel Ions on Roots of Allium Cepa L. Cytologia 2017, 83, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, A.; Khan, M.S. Heavy Metal-Mediated Toxicity to Maize: Oxidative Damage, Antioxidant Defence Response and Metal Distribution in Plant Organs. Int. J. Environ. Sci. Technol. 2019, 16, 4873–4886. [Google Scholar] [CrossRef]
- Demirezen Yilmaz, D.; Uruç Parlak, K. Antioxidative Parameters in the Opposite-Leaved Pondweed (Gronlendia Densa) in Response to Nickel Stress. Chem. Speciat. Bioavailab. 2011, 23, 71–79. [Google Scholar] [CrossRef]
- Wang, G.; Xiao, Q.; Tariq, M.; Peng, C.; Wu, J.; Zhang, W. Unveiling the Potential Tolerance and Physiological Response Mechanisms of Wheat after Exposure to Nickel in a Soil–Plant System. ACS Agric. Sci. Technol. 2022, 2, 941–949. [Google Scholar] [CrossRef]
- Fourati, E.; Vogel-Mikuš, K.; Bettaieb, T.; Kavčič, A.; Kelemen, M.; Vavpetič, P.; Pelicon, P.; Abdelly, C.; Ghnaya, T. Physiological Response and Mineral Elements Accumulation Pattern in Sesuvium Portulacastrum L. Subjected In Vitro to Nickel. Chemosphere 2019, 219, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- The Gene Ontology Consortium. The Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Luo, Z.-B.; He, J.; Polle, A.; Rennenberg, H. Heavy Metal Accumulation and Signal Transduction in Herbaceous and Woody Plants: Paving the Way for Enhancing Phytoremediation Efficiency. Biotechnol. Adv. 2016, 34, 1131–1148. [Google Scholar] [CrossRef]
- Sirhindi, G.; Mir, M.A.; Abd-Allah, E.F.; Ahmad, P.; Gucel, S. Jasmonic Acid Modulates the Physio-Biochemical Attributes, Antioxidant Enzyme Activity, and Gene Expression in Glycine Max under Nickel Toxicity. Front. Plant Sci. 2016, 7, 591. [Google Scholar] [CrossRef] [Green Version]
- Valivand, M.; Amooaghaie, R.; Ahadi, A. Interplay between Hydrogen Sulfide and Calcium/Calmodulin Enhances Systemic Acquired Acclimation and Antioxidative Defense against Nickel Toxicity in Zucchini. Environ. Exp. Bot. 2019, 158, 40–50. [Google Scholar] [CrossRef]
- Mahawar, L.; Kumar, R.; Shekhawat, G.S. Evaluation of Heme Oxygenase 1 (HO 1) in Cd and Ni Induced Cytotoxicity and Crosstalk with ROS Quenching Enzymes in Two to Four Leaf Stage Seedlings of Vigna radiata. Protoplasma 2018, 255, 527–545. [Google Scholar] [CrossRef]
- Kazemi, N.; Khavari-Nejad, R.A.; Fahimi, H.; Saadatmand, S.; Nejad-Sattari, T. Effects of Exogenous Salicylic Acid and Nitric Oxide on Lipid Peroxidation and Antioxidant Enzyme Activities in Leaves of Brassica napus L. under Nickel Stress. Sci. Hortic. 2010, 126, 402–407. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Muszyńska, E.; Hanus-Fajerska, E.; Dziurka, K.; Dziurka, M. Evaluation of the Protective Role of Exogenous Growth Regulators against Ni Toxicity in Woody Shrub Daphne Jasminea. Planta 2018, 248, 1365–1381. [Google Scholar] [CrossRef] [Green Version]
- Nazir, F.; Hussain, A.; Fariduddin, Q. Interactive Role of Epibrassinolide and Hydrogen Peroxide in Regulating Stomatal Physiology, Root Morphology, Photosynthetic and Growth Traits in Solanum lycopersicum L. under Nickel Stress. Environ. Exp. Bot. 2019, 162, 479–495. [Google Scholar] [CrossRef]
- Kolbert, Z.; Oláh, D.; Molnár, Á.; Szőllősi, R.; Erdei, L.; Ördög, A. Distinct Redox Signalling and Nickel Tolerance in Brassica Juncea and Arabidopsis Thaliana. Ecotoxicol. Environ. Saf. 2020, 189, 109989. [Google Scholar] [CrossRef]
- Stearns, J.C.; Shah, S.; Greenberg, B.M.; Dixon, D.G.; Glick, B.R. Tolerance of Transgenic Canola Expressing 1-Aminocyclopropane-1-Carboxylic Acid Deaminase to Growth Inhibition by Nickel. Plant Physiol. Biochem. 2005, 43, 701–708. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Baloch, A.R.; Sun, J.; Shu, S.; Wang, Y.; Ahammed, G.J.; Kabir, K.; Roy, R. Melatonin Alleviates Nickel Phytotoxicity by Improving Photosynthesis, Secondary Metabolism and Oxidative Stress Tolerance in Tomato Seedlings. Ecotoxicol. Environ. Saf. 2020, 197, 110593. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, X.; Wei, T.; Zhou, R.; Muhammad, H.; Hua, L.; Ren, X.; Guo, J.; Ding, Y. Accumulation and Fixation of Cd by Tomato Cell Wall Pectin under Cd Stress. Environ. Exp. Bot. 2019, 167, 103829. [Google Scholar] [CrossRef]
- Fan, J.-L.; Wei, X.-Z.; Wan, L.-C.; Zhang, L.-Y.; Zhao, X.-Q.; Liu, W.-Z.; Hao, H.-Q.; Zhang, H.-Y. Disarrangement of Actin Filaments and Ca2+ Gradient by CdCl2 Alters Cell Wall Construction in Arabidopsis Thaliana Root Hairs by Inhibiting Vesicular Trafficking. J. Plant Physiol. 2011, 168, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, Y.; Tian, J.; Zhu, Y.; Fan, J. Changes in Sucrose Metabolism in Maize Varieties with Different Cadmium Sensitivities under Cadmium Stress. PLoS ONE 2020, 15, e0243835. [Google Scholar] [CrossRef]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Effect of Nickel and Grafting Combination on Yield, Fruit Quality, Antioxidative Enzyme Activities, Lipid Peroxidation, and Mineral Composition of Tomato. J. Plant Nutr. Soil Sci. 2015, 178, 848–860. [Google Scholar] [CrossRef]
- Qin, T.; Tian, Q.; Wang, G.; Xiong, L. LOWER TEMPERATURE 1 Enhances ABA Responses and Plant Drought Tolerance by Modulating the Stability and Localization of C2-Domain ABA-Related Proteins in Arabidopsis. Mol. Plant 2019, 12, 1243–1258. [Google Scholar] [CrossRef]
- Yu, M.; Li, R.; Cui, Y.; Chen, W.; Li, B.; Zhang, X.; Bu, Y.; Cao, Y.; Xing, J.; Jewaria, P.K.; et al. The RALF1-FERONIA Interaction Modulates Endocytosis to Mediate Control of Root Growth in Arabidopsis. Development 2020, 147, dev189902. [Google Scholar] [CrossRef]
- Shimomura, S. Identification of a Glycosylphosphatidylinositol-Anchored Plasma Membrane Protein Interacting with the C-Terminus of Auxin-Binding Protein 1: A Photoaffinity Crosslinking Study. Plant Mol. Biol. 2006, 60, 663–677. [Google Scholar] [CrossRef]
- Altaf, M.A.; Hao, Y.; He, C.; Mumtaz, M.A.; Shu, H.; Fu, H.; Wang, Z. Physiological and Biochemical Responses of Pepper (Capsicum annuum L.) Seedlings to Nickel Toxicity. Front. Plant Sci. 2022, 13, 950392. [Google Scholar] [CrossRef]
- Hou, W.-C.; Lin, Y.-H. Sweet Potato (Ipomoea batatas (L.) Lam) Trypsin Inhibitors, the Major Root Storage Proteins, Inhibit One Endogenous Serine Protease Activity. Plant Sci. 2002, 163, 733–739. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.; Takano, T. Two Cysteine Proteinase Inhibitors from Arabidopsis Thaliana, AtCYSa and AtCYSb, Increasing the Salt, Drought, Oxidation and Cold Tolerance. Plant Mol. Biol. 2008, 68, 131–143. [Google Scholar] [CrossRef]
- Zhao, Y.; Gao, C.; Shi, F.; Yun, L.; Jia, Y.; Wen, J. Transcriptomic and Proteomic Analyses of Drought Responsive Genes and Proteins in Agropyron Mongolicum Keng. Curr. Plant Biol. 2018, 14, 19–29. [Google Scholar] [CrossRef]
- Guerra, F.P.; Reyes, L.; Vergara-Jaque, A.; Campos-Hernández, C.; Gutiérrez, A.; Pérez-Díaz, J.; Pérez-Díaz, R.; Blaudez, D.; Ruíz-Lara, S. Populus Deltoides Kunitz Trypsin Inhibitor 3 Confers Metal Tolerance and Binds Copper, Revealing a New Defensive Role against Heavy Metal Stress. Environ. Exp. Bot. 2015, 115, 28–37. [Google Scholar] [CrossRef]
- Major, I.T.; Constabel, C.P. Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against Herbivores. Plant Physiol. 2008, 146, 888–903. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Khan, N.A.; Bano, B. In-Sights into the Effect of Heavy Metal Stress on the Endogenous Mustard Cystatin. Int. J. Biol. Macromol. 2017, 105, 1138–1147. [Google Scholar] [CrossRef]
- Schützendübel, A.; Polle, A. Plant Responses to Abiotic Stresses: Heavy Metal-induced Oxidative Stress and Protection by Mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef]
- Jacques, S.; Ghesquière, B.; De Bock, P.-J.; Demol, H.; Wahni, K.; Willems, P.; Messens, J.; Van Breusegem, F.; Gevaert, K. Protein Methionine Sulfoxide Dynamics in Arabidopsis Thaliana under Oxidative Stress[S]. Mol. Cell. Proteom. 2015, 14, 1217–1229. [Google Scholar] [CrossRef] [Green Version]
- Arefifard, M.; Mahdieh, M.; Amirjani, M. Study of the Effect of Nickel Heavy Metals on Some Physiological Parameters of Catharanthus Roseus. Nat. Prod. Res. 2014, 28, 1499–1502. [Google Scholar] [CrossRef]
- Li, J.; Brader, G.; Palva, E.T. Kunitz Trypsin Inhibitor: An Antagonist of Cell Death Triggered by Phytopathogens and Fumonisin B1 in Arabidopsis. Mol. Plant 2008, 1, 482–495. [Google Scholar] [CrossRef] [Green Version]
- Radisky, E.S.; Lee, J.M.; Lu, C.-J.K.; Koshland, D.E. Insights into the Serine Protease Mechanism from Atomic Resolution Structures of Trypsin Reaction Intermediates. Proc. Natl. Acad. Sci. USA 2006, 103, 6835–6840. [Google Scholar] [CrossRef]
- Qi, Z.-Y.; Ahammed, G.J.; Jiang, C.-Y.; Li, C.-X.; Zhou, J. The E3 Ubiquitin Ligase Gene SlRING1 Is Essential for Plant Tolerance to Cadmium Stress in Solanum Lycopersicum. J. Biotechnol. 2020, 324, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Van Hoewyk, D.; Taskin, M.B.; Yaprak, A.E.; Turgay, O.C.; Ergul, A. Profiling of Proteasome Activity in Alyssum Species on Serpentine Soils in Turkey Reveals Possible Insight into Nickel Tolerance and Accumulation. Plant Physiol. Biochem. 2018, 124, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.C.; Moon, J.-C.; Chapagain, S.; Oh, D.G.; Kim, J.J.; Jang, C.S. Role of Salt-Induced RING Finger Protein 3 (OsSIRP3), a Negative Regulator of Salinity Stress Response by Modulating the Level of Its Target Proteins. Environ. Exp. Bot. 2018, 155, 21–30. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Liu, H.; Chong, K.; Xu, Y. Roles of Ubiquitination-Mediated Protein Degradation in Plant Responses to Abiotic Stresses. Environ. Exp. Bot. 2015, 114, 92–103. [Google Scholar] [CrossRef]
- Liu, C.-X.; Yang, T.; Zhou, H.; Ahammed, G.J.; Qi, Z.-Y.; Zhou, J. The E3 Ubiquitin Ligase Gene Sl1 Is Critical for Cadmium Tolerance in Solanum lycopersicum L. Antioxidants 2022, 11, 456. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, C.-X.; Li, X.; Liu, A.; Chen, S.; Zhou, J. Overexpression of Tomato RING E3 Ubiquitin Ligase Gene SlRING1 Confers Cadmium Tolerance by Attenuating Cadmium Accumulation and Oxidative Stress. Physiol. Plant. 2021, 173, 449–459. [Google Scholar] [CrossRef]
- Ko, J.-H.; Yang, S.H.; Han, K.-H. Upregulation of an Arabidopsis RING-H2 Gene, XERICO, Confers Drought Tolerance through Increased Abscisic Acid Biosynthesis. Plant J. 2006, 47, 343–355. [Google Scholar] [CrossRef]
- Zeng, D.-E.; Hou, P.; Xiao, F.; Liu, Y. Overexpressing a Novel RING-H2 Finger Protein Gene, OsRHP1, Enhances Drought and Salt Tolerance in Rice (Oryza sativa L.). J. Plant Biol. 2014, 57, 357–365. [Google Scholar] [CrossRef]
- Chung, H.S.; Howe, G.A. A Critical Role for the TIFY Motif in Repression of Jasmonate Signaling by a Stabilized Splice Variant of the JASMONATE ZIM-Domain Protein JAZ10 in Arabidopsis. Plant Cell 2009, 21, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Hakata, M.; Muramatsu, M.; Nakamura, H.; Hara, N.; Kishimoto, M.; Iida-Okada, K.; Kajikawa, M.; Imai-Toki, N.; Toki, S.; Nagamura, Y.; et al. Overexpression of TIFY Genes Promotes Plant Growth in Rice through Jasmonate Signaling. Biosci. Biotechnol. Biochem. 2017, 81, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, E.; Marquis, V.; Poirier, L.; Aubert, Y.; Zumsteg, J.; Ménard, R.; Miesch, L.; Heitz, T. Jasmonic Acid Oxidase 2 Hydroxylates Jasmonic Acid and Represses Basal Defense and Resistance Responses against Botrytis Cinerea Infection. Mol. Plant 2017, 10, 1159–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noir, S.; Bömer, M.; Takahashi, N.; Ishida, T.; Tsui, T.-L.; Balbi, V.; Shanahan, H.; Sugimoto, K.; Devoto, A. Jasmonate Controls Leaf Growth by Repressing Cell Proliferation and the Onset of Endoreduplication While Maintaining a Potential Stand-By Mode. Plant Physiol. 2013, 161, 1930–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azeem, U. Ameliorating Nickel Stress by Jasmonic Acid Treatment in Zea mays L. Russ. Agric. Sci. 2018, 44, 209–215. [Google Scholar] [CrossRef]
- Verma, G.; Srivastava, D.; Narayan, S.; Shirke, P.A.; Chakrabarty, D. Exogenous Application of Methyl Jasmonate Alleviates Arsenic Toxicity by Modulating Its Uptake and Translocation in Rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2020, 201, 110735. [Google Scholar] [CrossRef] [PubMed]
- Rakwal, R.; Komatsu, S. Jasmonic Acid-Induced Necrosis and Drastic Decreases in Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase in Rice Seedlings under Light Involves Reactive Oxygen Species. J. Plant Physiol. 2001, 158, 679–688. [Google Scholar] [CrossRef]
- Maksymiec, W.; Krupa, Z. The Effects of Short-Term Exposition to Cd, Excess Cu Ions and Jasmonate on Oxidative Stress Appearing in Arabidopsis Thaliana. Environ. Exp. Bot. 2006, 57, 187–194. [Google Scholar] [CrossRef]
- Gupta, V.; Willits, M.G.; Glazebrook, J. Arabidopsis Thaliana EDS4 Contributes to Salicylic Acid (SA)-Dependent Expression of Defense Responses: Evidence for Inhibition of Jasmonic Acid Signaling by SA. Mol. Plant-Microbe Interact. 2000, 13, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Golldack, D.; Vera, P.; Dietz, K.-J. Expression of Subtilisin-like Serine Proteases in Arabidopsis Thaliana Is Cell-Specific and Responds to Jasmonic Acid and Heavy Metals with Developmental Differences. Physiol. Plant. 2003, 118, 64–73. [Google Scholar] [CrossRef]
- Xiao, L.; Shang, X.-H.; Cao, S.; Xie, X.-Y.; Zeng, W.-D.; Lu, L.-Y.; Chen, S.-B.; Yan, H.-B. Comparative Physiology and Transcriptome Analysis Allows for Identification of LncRNAs Imparting Tolerance to Drought Stress in Autotetraploid Cassava. BMC Genom. 2019, 20, 514. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Li, L.; Zhang, W.; Cheng, H.; Sun, N.; Cheng, S.; Wang, Y. Isolation, Characterization, and Function Analysis of a Flavonol Synthase Gene from Ginkgo Biloba. Mol. Biol. Rep. 2012, 39, 2285–2296. [Google Scholar] [CrossRef]
- Burbulis, I.E.; Iacobucci, M.; Shirley, B.W. A Null Mutation in the First Enzyme of Flavonoid Biosynthesis Does Not Affect Male Fertility in Arabidopsis. Plant Cell 1996, 8, 1013–1025. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.-Y.; Sharma, S.B.; Paiva, N.L.; Ferreira, D.; Dixon, R.A. Role of Anthocyanidin Reductase, Encoded by BANYULS in Plant Flavonoid Biosynthesis. Science 2003, 299, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Takos, A.M.; Ubi, B.E.; Robinson, S.P.; Walker, A.R. Condensed Tannin Biosynthesis Genes Are Regulated Separately from Other Flavonoid Biosynthesis Genes in Apple Fruit Skin. Plant Sci. 2006, 170, 487–499. [Google Scholar] [CrossRef]
- Mahajan, M.; Ahuja, P.S.; Yadav, S.K. Post-Transcriptional Silencing of Flavonol Synthase MRNA in Tobacco Leads to Fruits with Arrested Seed Set. PLoS ONE 2011, 6, e28315. [Google Scholar] [CrossRef]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid Accumulation in Arabidopsis Repressed in Lignin Synthesis Affects Auxin Transport and Plant Growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, Y.; Zhu, C.; Yao, X.; Zheng, Z.; Tian, Z.; Cai, X. EkFLS Overexpression Promotes Flavonoid Accumulation and Abiotic Stress Tolerance in Plant. Physiol. Plant. 2021, 172, 1966–1982. [Google Scholar] [CrossRef]
- Baozhu, L.; Ruonan, F.; Yanting, F.; Runan, L.; Hui, Z.; Tingting, C.; Jiong, L.; Han, L.; Xiang, Z.; Chun-peng, S. The Flavonoid Biosynthesis Regulator PFG3 Confers Drought Stress Tolerance in Plants by Promoting Flavonoid Accumulation. Environ. Exp. Bot. 2022, 196, 104792. [Google Scholar] [CrossRef]
- Verdan, A.M.; Wang, H.C.; García, C.R.; Henry, W.P.; Brumaghim, J.L. Iron Binding of 3-Hydroxychromone, 5-Hydroxychromone, and Sulfonated Morin: Implications for the Antioxidant Activity of Flavonols with Competing Metal Binding Sites. J. Inorg. Biochem. 2011, 105, 1314–1322. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Younts, T.L.B.; Muday, G.K. Flavonols Control Pollen Tube Growth and Integrity by Regulating ROS Homeostasis during High-Temperature Stress. Proc. Natl. Acad. Sci. USA 2018, 115, E11188–E11197. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Man, C.; Xie, Y.; Yan, J.; Chu, J.; Huang, J. A Crucial Role of GA-Regulated Flavonol Biosynthesis in Root Growth of Arabidopsis. Mol. Plant 2019, 12, 521–537. [Google Scholar] [CrossRef] [Green Version]
- Rahman, H.; Sabreen, S.; Alam, S.; Kawai, S. Effects of Nickel on Growth and Composition of Metal Micronutrients in Barley Plants Grown in Nutrient Solution. J. Plant Nutr. 2005, 28, 393–404. [Google Scholar] [CrossRef]
- El-Sheekh, M.M. Inhibition of Photosystem II in the Green Alga Scenedesmus Obliquus by Nickel. Biochem. Physiol. Pflanz. 1993, 188, 363–372. [Google Scholar] [CrossRef]
- Mohanty, N.; Vass, I.; Demeter, S. Impairment of Photosystem 2 Activity at the Level of Secondary Quinone Electron Acceptor in Chloroplasts Treated with Cobalt, Nickel and Zinc Ions. Physiol. Plant. 1989, 76, 386–390. [Google Scholar] [CrossRef]
- Guo, M.; Perez, C.; Wei, Y.; Rapoza, E.; Su, G.; Bou-Abdallah, F.; Chasteen, N.D. Iron-Binding Properties of Plant Phenolics and Cranberry’s Bio-Effects. Dalton Trans. Camb. Engl. 2007, 43, 4951–4961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chobot, V.; Huber, C.; Trettenhahn, G.; Hadacek, F. (±)-Catechin: Chemical Weapon, Antioxidant, or Stress Regulator? J. Chem. Ecol. 2009, 35, 980–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubicova, L.; Bachmann, G.; Weckwerth, W.; Chobot, V. (±)-Catechin—A Mass-Spectrometry-Based Exploration Coordination Complex Formation with FeII and FeIII. Cells 2022, 11, 958. [Google Scholar] [CrossRef]
- Melidou, M.; Riganakos, K.; Galaris, D. Protection against Nuclear DNA Damage Offered by Flavonoids in Cells Exposed to Hydrogen Peroxide: The Role of Iron Chelation. Free Radic. Biol. Med. 2005, 39, 1591–1600. [Google Scholar] [CrossRef]
- Lesjak, M.; Hoque, R.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.S.; Sharp, P.A. Quercetin Inhibits Intestinal Iron Absorption and Ferroportin Transporter Expression In Vivo and In Vitro. PLoS ONE 2014, 9, e102900. [Google Scholar] [CrossRef] [Green Version]
- Lesjak, M.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.S. Quercetin Inhibits Intestinal Non-Haem Iron Absorption by Regulating Iron Metabolism Genes in the Tissues. Eur. J. Nutr. 2019, 58, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Wang, X.; Yao, X.; Chen, X.; Lu, K.; Hu, Y.; Wang, Z.; Mu, Y.; Zhang, L.; Dong, H. Rice Aquaporin OsPIP2;2 Is a Water-Transporting Facilitator in Relevance to Drought-Tolerant Responses. Plant Direct 2021, 5, e338. [Google Scholar] [CrossRef]
- Barozzi, F.; Papadia, P.; Stefano, G.; Renna, L.; Brandizzi, F.; Migoni, D.; Fanizzi, F.P.; Piro, G.; Di Sansebastiano, G.-P. Variation in Membrane Trafficking Linked to SNARE AtSYP51 Interaction With Aquaporin NIP1;1. Front. Plant Sci. 2019, 9, 1949. [Google Scholar] [CrossRef] [Green Version]
- Kholodova, V.; Volkov, K.; Abdeyeva, A.; Kuznetsov, V. Water Status in Mesembryanthemum Crystallinum under Heavy Metal Stress. Environ. Exp. Bot. 2011, 71, 382–389. [Google Scholar] [CrossRef]
- Ranocha, P.; Dima, O.; Nagy, R.; Felten, J.; Corratgé-Faillie, C.; Novák, O.; Morreel, K.; Lacombe, B.; Martinez, Y.; Pfrunder, S.; et al. Arabidopsis WAT1 Is a Vacuolar Auxin Transport Facilitator Required for Auxin Homoeostasis. Nat. Commun. 2013, 4, 2625. [Google Scholar] [CrossRef] [Green Version]
- Lešková, A.; Zvarík, M.; Araya, T.; Giehl, R.F.H. Nickel Toxicity Targets Cell Wall-Related Processes and PIN2-Mediated Auxin Transport to Inhibit Root Elongation and Gravitropic Responses in Arabidopsis. Plant Cell Physiol. 2020, 61, 519–535. [Google Scholar] [CrossRef] [Green Version]
- Hanika, K.; Schipper, D.; Chinnappa, S.; Oortwijn, M.; Schouten, H.J.; Thomma, B.P.H.J.; Bai, Y. Impairment of Tomato WAT1 Enhances Resistance to Vascular Wilt Fungi Despite Severe Growth Defects. Front. Plant Sci. 2021, 12, 721674. [Google Scholar] [CrossRef]
- Denancé, N.; Ranocha, P.; Oria, N.; Barlet, X.; Rivière, M.-P.; Yadeta, K.A.; Hoffmann, L.; Perreau, F.; Clément, G.; Maia-Grondard, A.; et al. Arabidopsis Wat1 (Walls Are Thin1)-Mediated Resistance to the Bacterial Vascular Pathogen, Ralstonia Solanacearum, Is Accompanied by Cross-Regulation of Salicylic Acid and Tryptophan Metabolism. Plant J. 2013, 73, 225–239. [Google Scholar] [CrossRef]
- Wang, D.; Pajerowska-Mukhtar, K.; Culler, A.H.; Dong, X. Salicylic Acid Inhibits Pathogen Growth in Plants through Repression of the Auxin Signaling Pathway. Curr. Biol. 2007, 17, 1784–1790. [Google Scholar] [CrossRef] [Green Version]
- Sinha, P.; Shukla, A.K.; Sharma, Y.K. Amelioration of Heavy-Metal Toxicity in Cauliflower by Application of Salicylic Acid. Commun. Soil Sci. Plant Anal. 2015, 46, 1309–1319. [Google Scholar] [CrossRef]
- Wang, H.; Feng, T.; Peng, X.; Yan, M.; Tang, X. Up-Regulation of Chloroplastic Antioxidant Capacity Is Involved in Alleviation of Nickel Toxicity of Zea Mays L. by Exogenous Salicylic Acid. Ecotoxicol. Environ. Saf. 2009, 72, 1354–1362. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Varshney, P.; Ahmad, A. Salicylic Acid Minimizes Nickel and/or Salinity-Induced Toxicity in Indian Mustard (Brassica Juncea) through an Improved Antioxidant System. Environ. Sci. Pollut. Res. 2012, 19, 8–18. [Google Scholar] [CrossRef]
- Taylor, N.G.; Gardiner, J.C.; Whiteman, R.; Turner, S.R. Cellulose Synthesis in the Arabidopsis Secondary Cell Wall. Cellulose 2004, 11, 329–338. [Google Scholar] [CrossRef]
- Song, X.-Q.; Liu, L.-F.; Jiang, Y.-J.; Zhang, B.-C.; Gao, Y.-P.; Liu, X.-L.; Lin, Q.-S.; Ling, H.-Q.; Zhou, Y.-H. Disruption of Secondary Wall Cellulose Biosynthesis Alters Cadmium Translocation and Tolerance in Rice Plants. Mol. Plant 2013, 6, 768–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cutoff | Standard (Two-Fold and FDR 0.05) | Low Stringency (Two-Fold and p Value 0.01) |
|---|---|---|
| Upregulated genes | 4128 | 11,903 |
| Downregulated genes | 3754 | 6332 |
| Total genes | 7882 | 18,235 |
| Rank | Gene ID | Res 1 | Res 2 | Res 3 | Water 1 | Water 2 | Water 3 | logFC | Adj. p Value | UniProt Description |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | TRINITY_DN2786_c0_g1 | 767.81 | 197.57 | 545.86 | 0 | 0 | 0 | 13.96 | 0.00116 | Predicted Protein |
| 1 | TRINITY_DN5716_c0_g1 | 2328.59 | 913.58 | 3881.87 | 0 | 7.03 | 0.41 | 13.34 | 0.00029 | Predicted Protein |
| 2 | TRINITY_DN57079_c0_g1 | 339.53 | 238.75 | 261.65 | 0 | 0 | 0 | 13.30 | 0.00002 | Predicted Protein |
| 3 | TRINITY_DN5965_c1_g1 | 1173.34 | 760.7 | 1106.06 | 0.33 | 0 | 0 | 13.28 | 0.00009 | Predicted Protein |
| 4 | TRINITY_DN258556_c0_g1 | 280.75 | 98.46 | 494.55 | 0 | 0 | 0 | 13.09 | 0.00181 | Predicted Protein |
| 5 | TRINITY_DN1368_c0_g1 | 1156.77 | 736.4 | 2060.57 | 0 | 1.3 | 0.07 | 12.99 | 0.00047 | Predicted Protein |
| 6 | TRINITY_DN2832_c0_g1 | 334.2 | 111.71 | 258.08 | 0 | 0 | 0 | 12.93 | 0.00056 | Predicted Protein |
| 7 | TRINITY_DN1628_c0_g1 | 646.38 | 288.02 | 710.02 | 0 | 0.32 | 0 | 12.82 | 0.00065 | Trypsin inhibitor [Cleaved into: Trypsin inhibitor chain A; Trypsin inhibitor chain B] |
| 8 | TRINITY_DN7061_c1_g1 | 158.35 | 218.82 | 172.81 | 0 | 0 | 0 | 12.69 | 0.00000 | Predicted Protein |
| 9 | TRINITY_DN690_c0_g1 | 494.67 | 136.83 | 407.74 | 0.05 | 0 | 0 | 12.50 | 0.00181 | Predicted Protein |
| 10 | TRINITY_DN5795_c0_g1 | 753.52 | 420.03 | 412.9 | 0 | 0.84 | 0 | 12.43 | 0.00032 | Predicted Protein |
| 11 | TRINITY_DN1520_c0_g1 | 398.05 | 358.51 | 936.72 | 0.02 | 0.65 | 0 | 11.81 | 0.00043 | Trypsin inhibitor [Cleaved into: Trypsin inhibitor chain A; Trypsin inhibitor chain B] |
| 12 | TRINITY_DN3861_c0_g1 | 179.52 | 38.1 | 108.51 | 0 | 0 | 0 | 11.70 | 0.00251 | Predicted Protein |
| 13 | TRINITY_DN40097_c0_g1 | 440.62 | 297.68 | 1698.09 | 0 | 3.39 | 0.69 | 11.62 | 0.00080 | Predicted Protein |
| 14 | TRINITY_DN2463_c0_g1 | 301.76 | 196.16 | 568.86 | 0 | 0.04 | 0.11 | 11.56 | 0.00056 | Predicted Protein |
| 15 | TRINITY_DN4524_c0_g3 | 64.05 | 74.91 | 115.6 | 0 | 0 | 0 | 11.54 | 0.00002 | Predicted Protein |
| 16 | TRINITY_DN792_c0_g1 | 149.05 | 126.17 | 86.37 | 0 | 0.03 | 0 | 11.54 | 0.00004 | ACT domain-containing protein ACR4 (Protein ACT DOMAIN REPEATS 4) |
| 17 | TRINITY_DN792_c0_g1 | 149.05 | 126.17 | 86.37 | 0 | 0.03 | 0 | 11.54 | 0.00004 | ACT domain-containing protein ACR5 (Protein ACT DOMAIN REPEATS 5) |
| 18 | TRINITY_DN129489_c0_g1 | 125.97 | 40.97 | 102.59 | 0 | 0 | 0 | 11.53 | 0.00085 | Predicted Protein |
| 19 | TRINITY_DN2914_c0_g1 | 134.07 | 79.52 | 144.69 | 0 | 0.03 | 0 | 11.51 | 0.00014 | Protein TIFY 10b, OsTIFY10b (Jasmonate ZIM domain-containing protein 7, OsJAZ7) (OsJAZ6) |
| 20 | TRINITY_DN2914_c0_g1 | 134.07 | 79.52 | 144.69 | 0 | 0.03 | 0 | 11.51 | 0.00014 | Protein TIFY 3B (Jasmonate ZIM domain-containing protein 12) |
| 21 | TRINITY_DN3536_c0_g1 | 51.58 | 119.55 | 84.19 | 0 | 0 | 0 | 11.51 | 0.00001 | Predicted Protein |
| 22 | TRINITY_DN1537_c0_g1 | 64.53 | 90.63 | 76.46 | 0 | 0 | 0 | 11.44 | 0.00000 | Predicted Protein |
| 23 | TRINITY_DN2075_c1_g1 | 81.81 | 56.05 | 84.87 | 0 | 0 | 0 | 11.38 | 0.00005 | Predicted Protein |
| 24 | TRINITY_DN12750_c0_g1 | 93.87 | 62.85 | 64.95 | 0 | 0 | 0 | 11.37 | 0.00005 | Predicted Protein |
| 25 | TRINITY_DN3685_c0_g2 | 524.13 | 169.45 | 298.36 | 0.01 | 0.58 | 0 | 11.33 | 0.00171 | Copia protein (Gag-int-pol protein) [Cleaved into: Copia VLP protein; Copia protease, EC 3.4.23.] |
| 0 | TRINITY_DN1118_c0_g1 | 0 | 0 | 0 | 27.63 | 15.12 | 24.7 | −11.36 | 4.86 × 10−5 | Flavonol synthase/flavanone 3-hydroxylase, FLS, EC 1.14.11.9, EC 1.14.20.6 |
| 1 | TRINITY_DN26931_c0_g1 | 0.16 | 0 | 0 | 65.61 | 45.82 | 36.39 | −11.19 | 9.47 × 10−5 | Probable aquaporin PIP2-8 (Plasma membrane intrinsic protein 2-8, AtPIP2;8) (Plasma membrane intrinsic protein 3b, PIP3b) |
| 2 | TRINITY_DN432_c0_g1 | 0 | 0.3 | 0 | 77.54 | 17.58 | 69.88 | −11.15 | 0.002533 | Predicted Protein |
| 3 | TRINITY_DN4059_c0_g1 | 0 | 0 | 0 | 20.09 | 12.1 | 19.58 | −10.99 | 4.10 × 10−5 | Predicted Protein |
| 4 | TRINITY_DN30654_c0_g1 | 0 | 0 | 0 | 14.69 | 11.78 | 14.4 | −10.67 | 1.63 × 10−5 | Predicted Protein |
| 5 | TRINITY_DN2314_c0_g1 | 0.03 | 0.13 | 0 | 40.88 | 14.56 | 52.04 | −10.43 | 0.001066 | Predicted Protein |
| 6 | TRINITY_DN69830_c0_g4 | 0 | 0 | 0 | 10.13 | 7.29 | 18.59 | −10.38 | 0.000101 | Predicted Protein |
| 7 | TRINITY_DN129793_c0_g1 | 0 | 0 | 0 | 8.28 | 13.37 | 9.36 | −10.22 | 9.45 × 10−6 | Putative UPF0481 protein At3g02645 |
| 8 | TRINITY_DN40558_c0_g1 | 0.04 | 0 | 0.05 | 36.31 | 14.48 | 19.64 | −10.08 | 0.000432 | Predicted Protein |
| 9 | TRINITY_DN522_c0_g3 | 0 | 0 | 0 | 8.24 | 4.71 | 17.93 | −10.05 | 0.000408 | Predicted Protein |
| 10 | TRINITY_DN1550_c0_g1 | 0 | 0.07 | 0 | 18.5 | 9.11 | 17.44 | −9.99 | 0.000209 | Predicted Protein |
| 11 | TRINITY_DN113586_c0_g1 | 0 | 0 | 0 | 7.25 | 5.74 | 13.28 | −9.94 | 8.70 × 10−5 | Predicted Protein |
| 12 | TRINITY_DN25689_c0_g1 | 0.06 | 0.09 | 0 | 26.01 | 16.36 | 31.07 | −9.92 | 0.000136 | Predicted Protein |
| 13 | TRINITY_DN26605_c0_g1 | 0 | 0 | 0 | 6.61 | 7.11 | 10.35 | −9.87 | 2.28 × 10−5 | Predicted Protein |
| 14 | TRINITY_DN31123_c0_g2 | 0 | 0 | 0 | 6.35 | 8.14 | 8.67 | −9.82 | 1.28 × 10−5 | Predicted Protein |
| 15 | TRINITY_DN4890_c0_g1 | 0 | 0 | 0.17 | 15.59 | 12.05 | 25.46 | −9.82 | 0.000174 | Predicted Protein |
| 16 | TRINITY_DN5062_c0_g2 | 0 | 0 | 0 | 10.4 | 7.97 | 3.99 | −9.72 | 0.000193 | Predicted Protein |
| 17 | TRINITY_DN3390_c0_g1 | 0 | 0 | 0 | 9.96 | 3.94 | 7.77 | −9.69 | 0.000273 | Predicted Protein |
| 18 | TRINITY_DN6314_c0_g1 | 0 | 0 | 0 | 7.61 | 6.68 | 5.98 | −9.66 | 2.86 × 10−5 | Predicted Protein |
| 19 | TRINITY_DN2507_c0_g1 | 0 | 0 | 0.61 | 32.43 | 13.12 | 23.67 | −9.65 | 0.000952 | Predicted Protein |
| 20 | TRINITY_DN53932_c0_g1 | 0.01 | 0 | 0.2 | 17.81 | 11.58 | 17.09 | −9.61 | 0.00016 | Predicted Protein |
| 21 | TRINITY_DN20386_c0_g1 | 0 | 0 | 0 | 7.77 | 6 | 5.13 | −9.55 | 5.04 × 10−5 | Predicted Protein |
| 22 | TRINITY_DN17540_c0_g1 | 0 | 0 | 0 | 10.32 | 6.62 | 3.33 | −9.55 | 0.000363 | Predicted Protein |
| 23 | TRINITY_DN51950_c1_g1 | 0 | 0 | 0 | 6.24 | 5.15 | 7.26 | −9.53 | 3.46 × 10−5 | Predicted Protein |
| 24 | TRINITY_DN59077_c1_g1 | 0 | 0.2 | 0 | 11.37 | 9.17 | 20.15 | −9.52 | 0.000196 | Predicted Protein |
| 25 | TRINITY_DN26_c1_g1 | 0 | 0 | 0 | 5.86 | 4.64 | 7.86 | −9.49 | 5.04 × 10−5 | Alpha-galactosidase, EC 3.2.1.22 (Alpha-D-galactoside galactohydrolase) (Melibiase) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moy, A.; Czajka, K.; Michael, P.; Nkongolo, K. Transcriptome Analysis Reveals Changes in Whole Gene Expression, Biological Process, and Molecular Functions Induced by Nickel in Jack Pine (Pinus banksiana). Plants 2023, 12, 2889. https://doi.org/10.3390/plants12152889
Moy A, Czajka K, Michael P, Nkongolo K. Transcriptome Analysis Reveals Changes in Whole Gene Expression, Biological Process, and Molecular Functions Induced by Nickel in Jack Pine (Pinus banksiana). Plants. 2023; 12(15):2889. https://doi.org/10.3390/plants12152889
Chicago/Turabian StyleMoy, Alistar, Karolina Czajka, Paul Michael, and Kabwe Nkongolo. 2023. "Transcriptome Analysis Reveals Changes in Whole Gene Expression, Biological Process, and Molecular Functions Induced by Nickel in Jack Pine (Pinus banksiana)" Plants 12, no. 15: 2889. https://doi.org/10.3390/plants12152889