
Effects of High Doses of Selenate, Selenite and Nano-Selenium on Biometrical Characteristics, Yield and Biofortification Levels of Vicia faba L. Cultivars
 ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
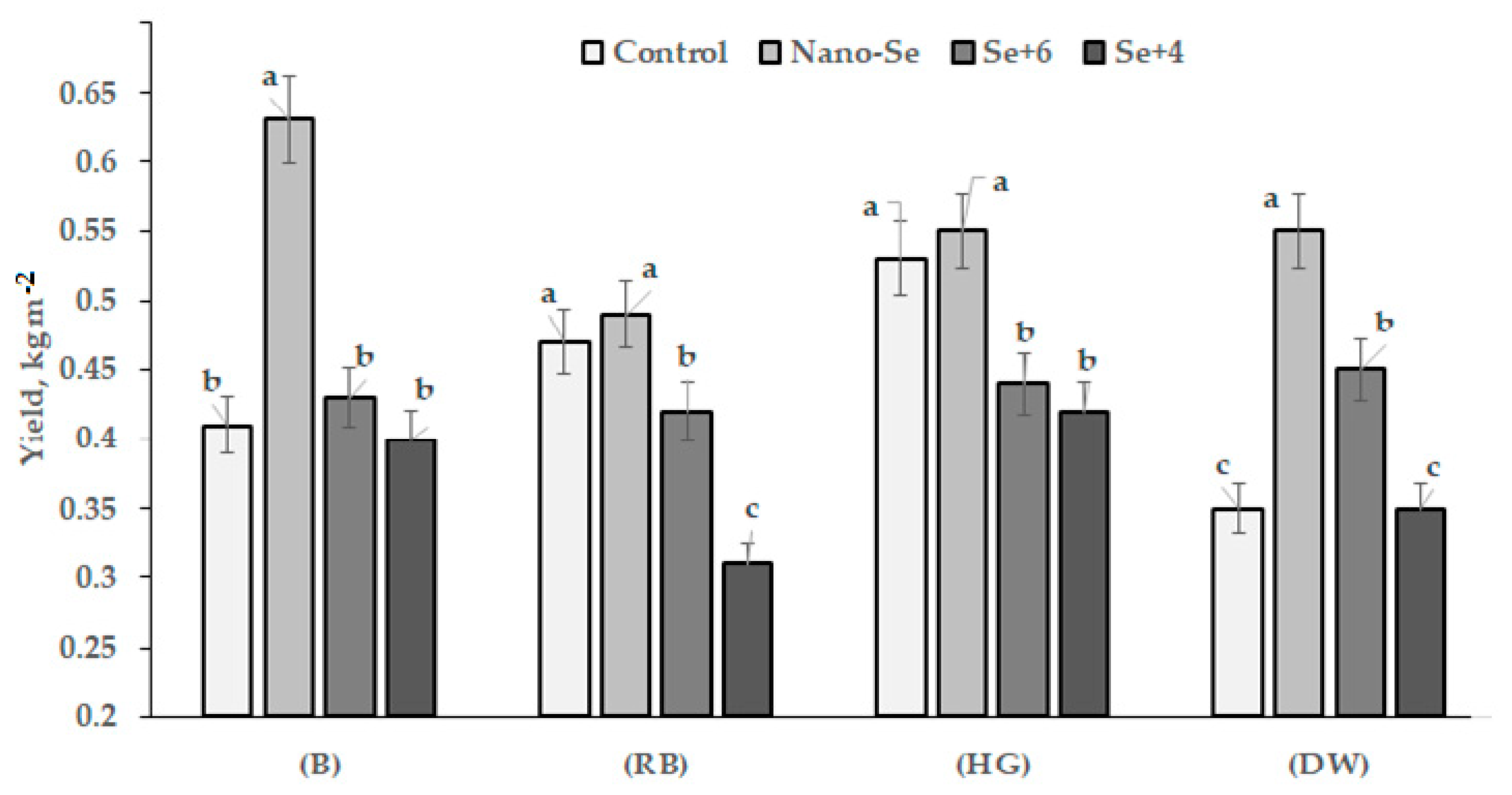
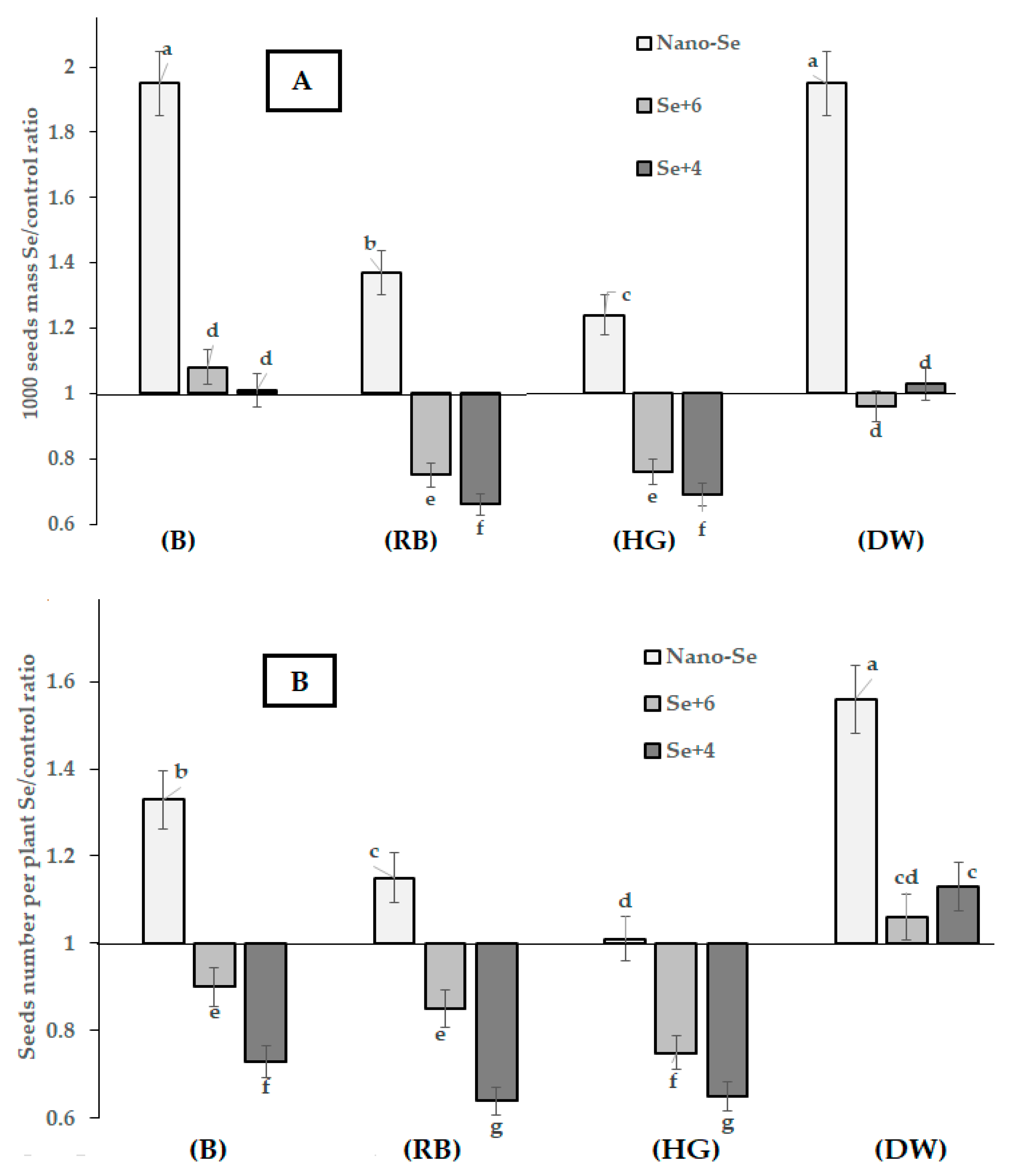
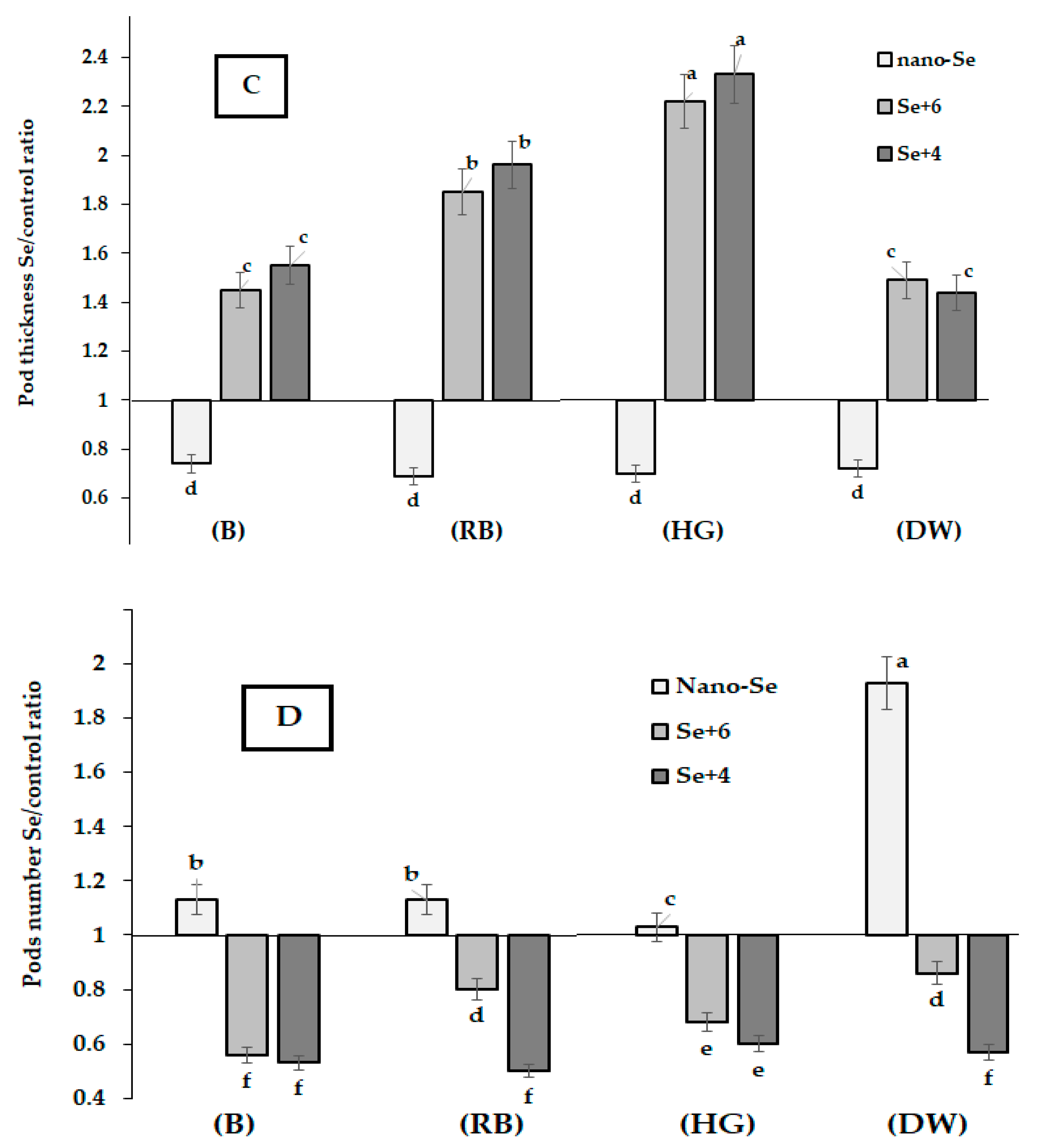
2.1. Yield and Biometrical Characteristics
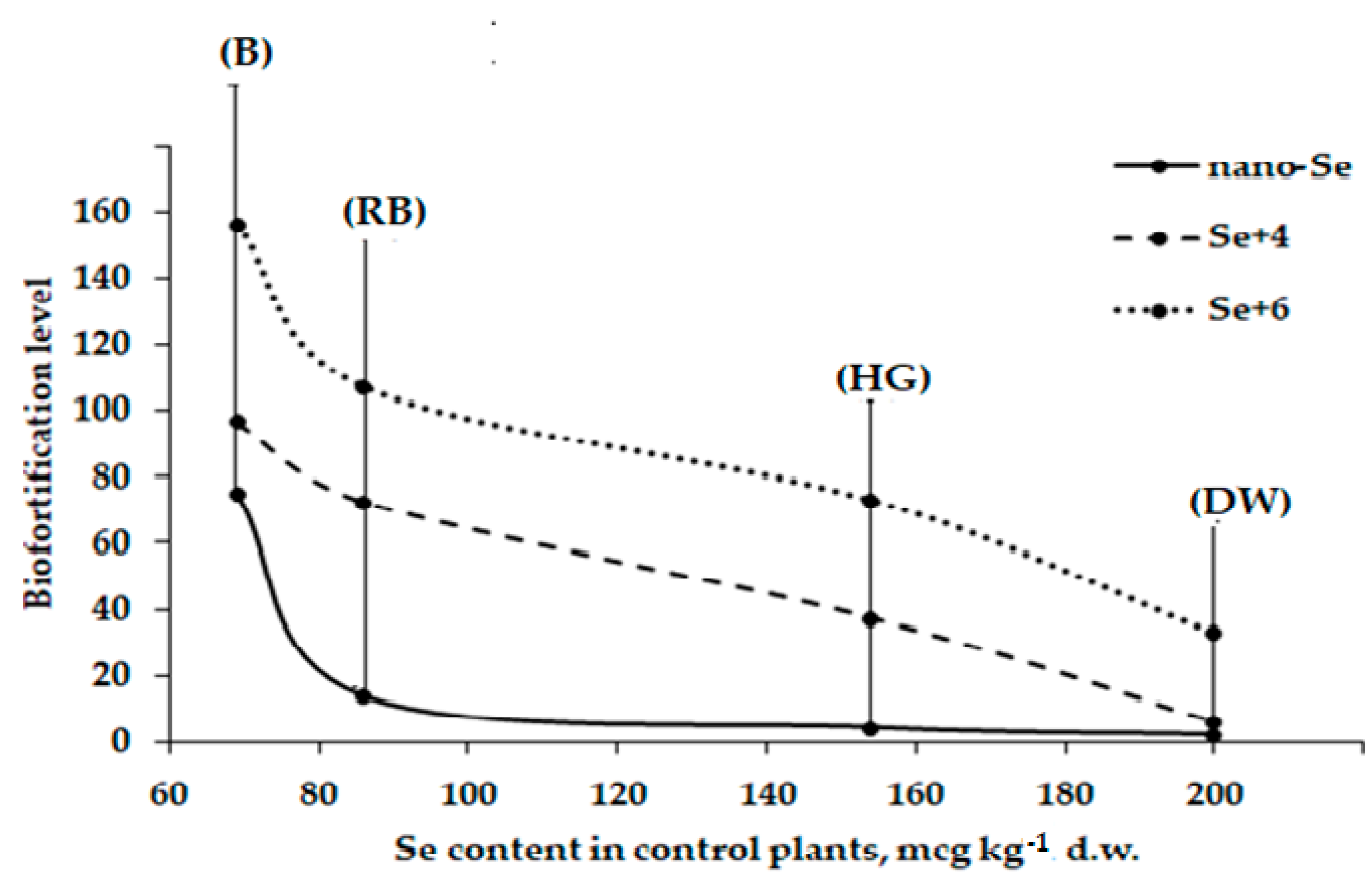
2.2. Se Accumulation
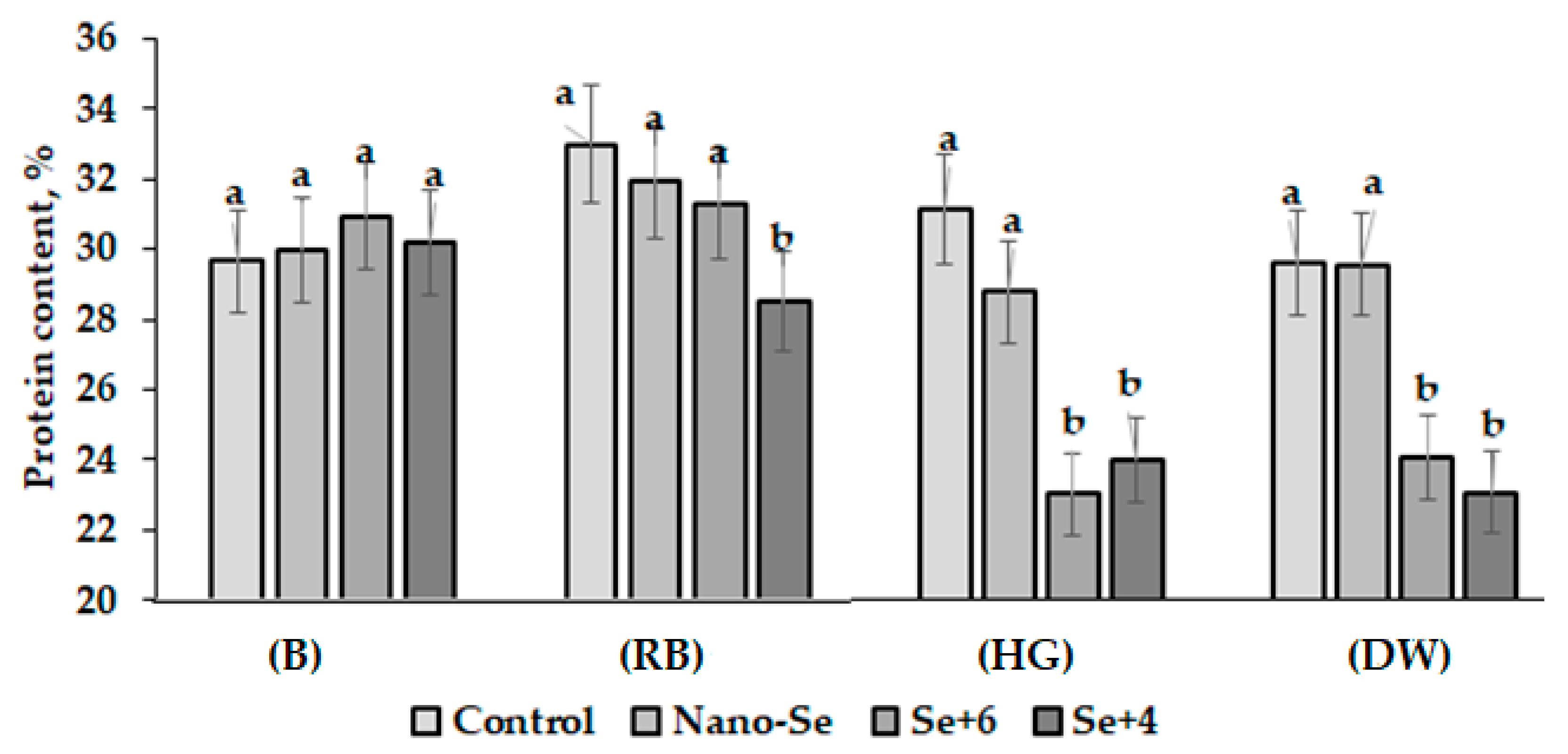
2.3. Protein
3. Material and Methods
3.1. Growing Conditions and Experimental Protocol
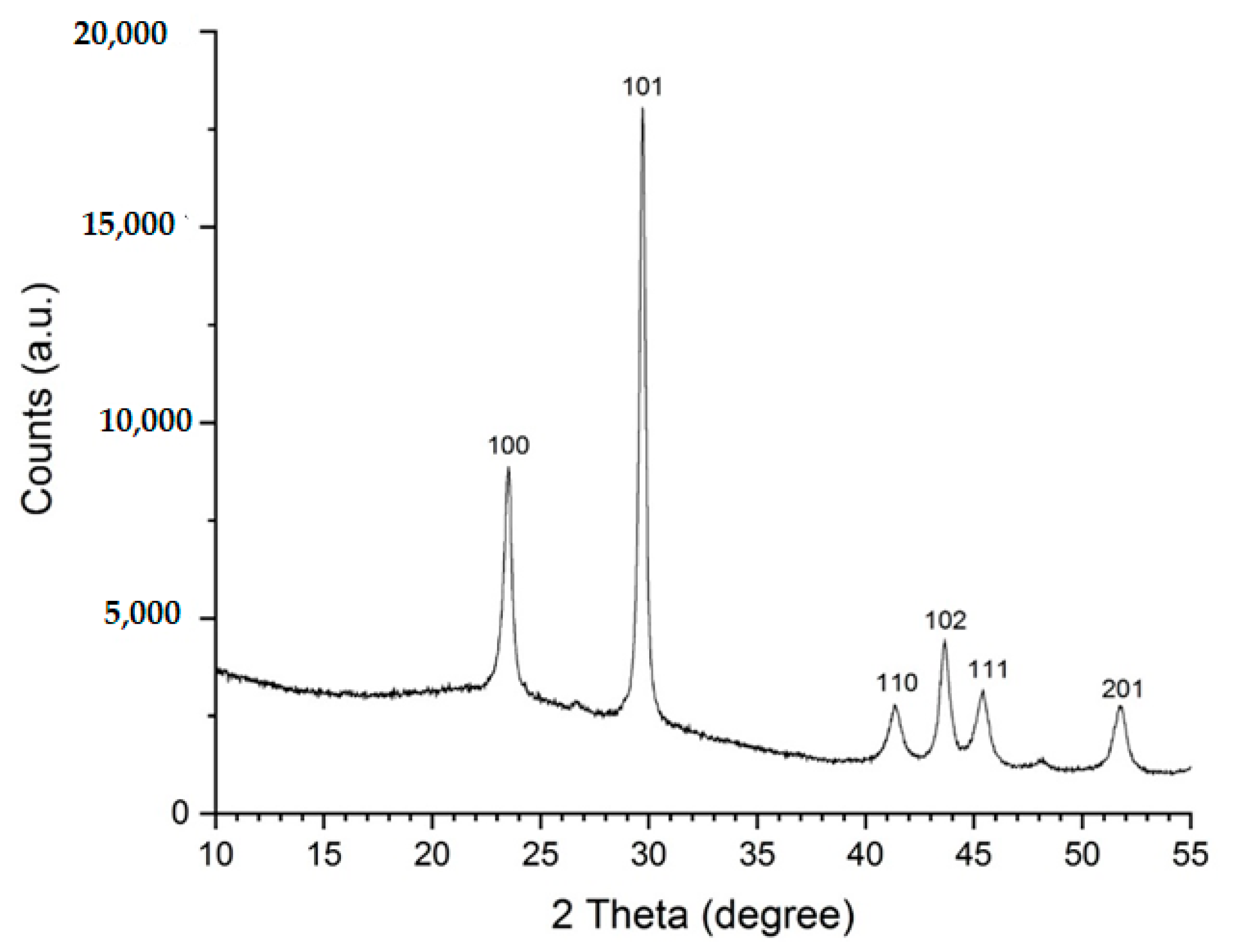
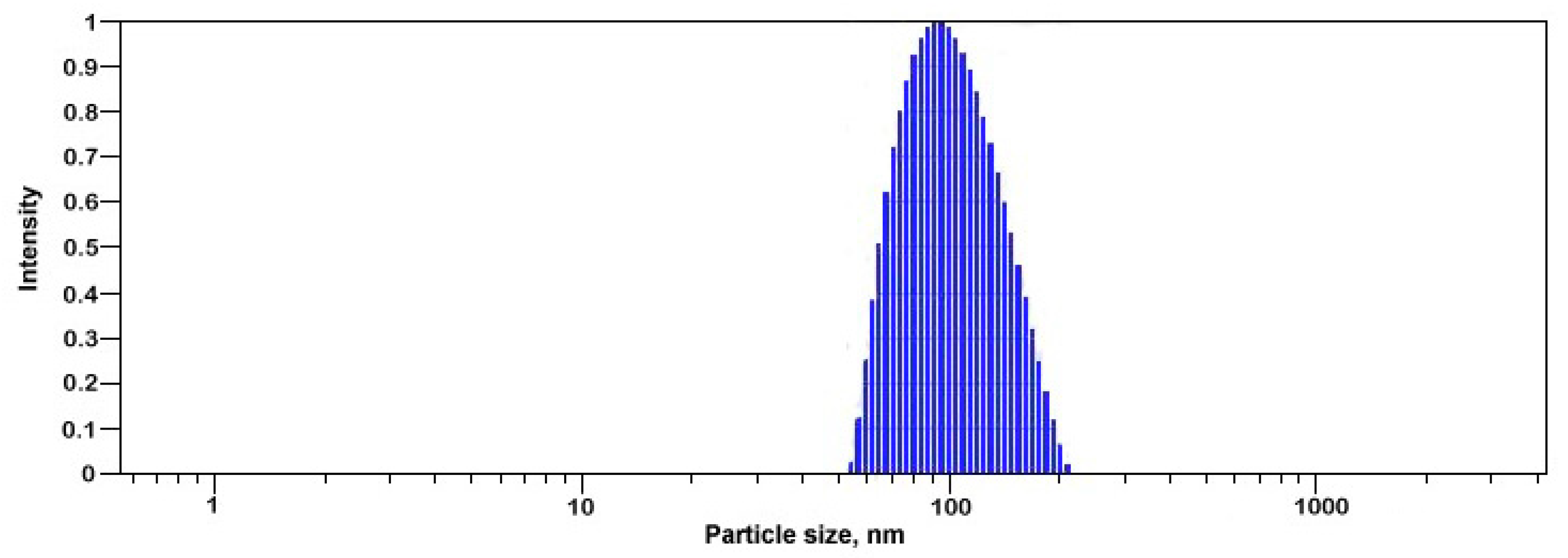
3.2. Preparation and Characterization of Selenium Colloidal Solution
3.3. Selenium
3.4. Protein Content
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, M.; Zhou, F.; Cheng, N.; Chen, P.; Ma, Y.; Zhai, H.; Qi, M.; Liu, N.; Liu, Y.; Meng, L. Bañuelos GS and Liang D Soil and foliar selenium application: Impact on accumulation, speciation, and bioaccessibility of selenium in wheat (Triticum aestivum L.). Front. Plant Sci. 2022, 13, 988627. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangjaidee, P.; Swedlund, P.; Xiang, J.; Yin, H.; Quek, S.Y. Selenium-enriched plant foods: Selenium accumulation, speciation, and health functionality. Front. Nutr. 2023, 9, 962312. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, Z.; Lehotai, N.; Molnár, Á.; Feigl, G. “The roots” of selenium toxicity: A new concept. Plant Signal Behav. 2016, 11, e1241935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Nahar, K.; Fujita, M. Selenium toxicity in plants and environment: Biogeochemistry and remediation possibilities. Plants 2020, 9, 1711. [Google Scholar] [CrossRef]
- Genchi, G.; Lauria, G.; Catalano, A.; Sinicropi, M.S.; Carocci, A. Biological activity of selenium and its impact on human health. Int. J. Mol. Sci. 2023, 24, 2633. [Google Scholar] [CrossRef]
- Xiao, T.; Boada, R.; Llugany, M.; Valiente, M. Co-application of Se and a biostimulant at different wheat growth stages: Influence on grain development. Plant Physiol. Biochem. 2021, 160, 184–192. [Google Scholar] [CrossRef]
- Subirana, M.A.; Boada, R.; Xiao, T.; Llugany, M.; Valiente, M. Direct and indirect selenium speciation in biofortified wheat: A tale of two techniques. Physiol. Plant. 2023, 175, e13843. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, Q.; Chen, H.; Xiong, H. Distribution and effects of natural selenium in soybean proteins and its protective role in soybean β-conglycinin (7S globulins) under AAPH-induced oxidative stress. Food Chem. 2019, 272, 201–209. [Google Scholar] [CrossRef]
- Smrkolj, P.; Osvald, M.; Osvald, J.; Stibilj, V. Selenium uptake and species distribution in selenium-enriched bean (Phaseolus vulgaris L.) seeds obtained by two different cultivations. Eur. Food Res. Technol. 2007, 225, 233–237. [Google Scholar] [CrossRef]
- Danso, O.P.; Asante-Badu, B.; Zhang, Z.; Song, J.; Wang, Z.; Yin, X.; Zhu, R. Selenium biofortification: Strategies, progress and challenges. Agriculture 2023, 13, 416. [Google Scholar] [CrossRef]
- Debnath, S.; Agarwal, A.; Kumar, N.R.; Bedi, A. Selenium-based drug development for antioxidant and anticancer activity. Future Pharmacol. 2022, 2, 595–607. [Google Scholar] [CrossRef]
- Dhull, S.B.; Kidwai, M.K.; Noor, R.; Chawla, P. A review of nutritional profile and processing of faba bean (Vicia faba L.). Leg. Sci. 2022, 4, e129. [Google Scholar] [CrossRef]
- Crépon, K.; Marget, P.; Peyronnet, C.; Carrouée, B.; Arese, P.; Duc, G. Nutritional value of faba bean (Vicia faba L.) seeds for food and feed. Field Crops Res. 2010, 115, 329–339. [Google Scholar] [CrossRef]
- Feedipedia. Faba Bean (Vicia Faba). 2018. Available online: https://www.feedipedia.org/node/4926 (accessed on 21 January 2020).
- Robinson, G.H.J.; Balk, J.; Domoney, C. Improving pulse crops as a source of protein, starch and micronutrients. Nutr. Bull. 2019, 44, 202–215. [Google Scholar] [CrossRef]
- Khazaei, H.; Vandenberg, A. Seed mineral composition and protein content of faba beans (Vicia faba L.) with contrasting tannin contents. Agronomy 2020, 10, 511. [Google Scholar] [CrossRef] [Green Version]
- Rahate, K.A.; Madhumita, M.; Prabhakar, P.K. Nutritional composition, anti-nutritional factors, pre-treatments-cum-processing impact and food formulation potential of faba bean (Vicia faba L.): A comprehensive review. LWT Food Sci. Technol. 2020, 138, 110796. [Google Scholar] [CrossRef]
- Boghdady, M.S.; Desoky, E.M.; Azoz, S.N.; Nassar, D.M.A. Effect of selenium on growth, physiological aspects and productivity of faba bean (Vicia faba L.). Egypt. J. Agron. 2017, 39, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Ravello, R.A.V.; de Oliveira, C.; Lessa, J.; Boas, L.V.V.; de Castro, E.M.; Guilherme, L.R.G.; Lopes, G. Selenium application influenced selenium biofortification and physiological traits in water-deficit common bean plants. Crop Pasture Sci. 2021, 73, 44–55. [Google Scholar] [CrossRef]
- Patel, P.; Trivedi, G.; Bhatt, S.; Patel, H.; Saraf, M. Development of sustainable biofortification strategy in Phraseolus vulgaris through selenorhizobacteria under selenium deficient region. Biosci. Biotechnol. Res. Comm. 2020, 13, 1–9. [Google Scholar]
- Hermosillo-Cereceres, M.A.; Sánchez-Chávez, E.; Guevara-Aguilar, A.; Muñoz-Márquez, E.; García-Bañuelos, M.L. Biofortification and distribution patterns of selenium in bean: Response to selenate and selenite. J. Food Agric. Environ. 2013, 11, 421–426. [Google Scholar]
- Silva, M.A.; de Sousa, G.F.; Corguinha, A.P.B.; de Lima Lessa, J.H.; Dinali, G.S.; Oliveira, C.; Lopes, G.l.; Amaral, D.; Brown, P.; Guilherme, L.R.G. Selenium biofortification of soybean genotypes in a tropical soil via Se-enriched phosphate fertilizers. Front. Plant Sci. 2022, 13, 988140. [Google Scholar] [CrossRef] [PubMed]
- Jakabová, S.; Čurlej, J.; Fikselová, M.; Harangozo, L.; Kozelová, D.; Hegedűs, O.; Hegedűsová, A. The use of biofortification for production of selenium enriched garden pea. Slovak J. Food Sci. 2020, 14, 437–442. [Google Scholar] [CrossRef]
- Golubkina, N.; Gomez, L.; Kekina, H.; Hallam, R.; Tallarita, A.; Cozzolino, E.; Torino, V.; Koshevarov, A.; Cuciniello, A.; Maiello, R.; et al. Joint selenium-iodine supply and arbuscular mycorrhizal fungi inoculation affect yield and quality of chickpea seeds and residual biomass. Plants 2020, 9, 804. [Google Scholar] [CrossRef]
- Bano, I.; Skalickova, S.; Sajjad, H.; Skladanka, J.; Horky, P. Uses of selenium nanoparticles in the plant production. Agronomy 2021, 11, 2229. [Google Scholar] [CrossRef]
- Silva, M.A.; Sousa, G.F.d.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; Corguinha, A.P.B.; Bueno, J.M.M.; Brunetto, G.; Leite, J.M.; Santos, A.A.d.; Lopes, G. Foliar application of selenium associated with a multi-nutrient fertilizer in soybean: Yield, grain quality, and critical selenium threshold. Plants 2023, 12, 2028. [Google Scholar] [CrossRef]
- Khan, Z.; Thounaojam, T.C.; Chowdhury, D.; Upadhyaya, H. The role of selenium and nano selenium on physiological responses in plant: A review. Plant Growth Regul. 2023, 100, 409–433. [Google Scholar] [CrossRef]
- Terry, N.; Zayed, A.M.; De Souza, M.P.; Tarun, A.S. Selenium in higher plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, J.; Yu, H. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes: Comparison with selenomethionine in mice. Free Radic. Biol. Med. 2011, 42, 1524–1533. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-induced upregulation of the antioxidant defense and methylglyoxal detoxification system reduces salinity induced damage in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 43, 1704–1721. [Google Scholar] [CrossRef]
- Samynathan, R.; Venkidasamy, B.; Ramya, K.; Muthuramalingam, P.; Shin, H.; Kumari, P.S.; Thangavel, S.; Sivanesan, I. A recent update on the impact of nano-selenium on plant growth, metabolism, and stress tolerance. Plants 2023, 12, 853. [Google Scholar] [CrossRef]
- El Lateef Gharib, F.A.; Zeid, I.M.; Ghazi, S.M.; Ahmed, E.Z. The response of cowpea (Vigna unguiculata L.) plants to foliar application of sodium selenate and selenium nanoparticles (SeNPs). J. Nanomater. Mol. Nanotechnol. 2019, 8, 4. [Google Scholar]
- Amina, Z.; Samar, O. Nano selenium: Reduction of severe hazards of Atrazine and promotion of changes in growth and gene expression patterns on Vicia faba seedlings. Afr. J. Biotechnol. 2019, 18, 502–510. [Google Scholar] [CrossRef] [Green Version]
- De Cillis, F.; Leoni, B.; Massaro, M.; Renna, M.; Santamaria, P. Yield and quality of faba bean (Vicia faba L. var. major) genotypes as a vegetable for freshcConsumption: A comparison between Italian landraces and commercial varieties. Agriculture 2019, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- El-Ramady, H.; Shedeed, S.I.; Fawzy, Z.F.; El Bassiony, A.E.-M.M.; El-Sawy, S.M.; Mahmoud, S.H.; Prokisch, J. Biofortification of Vegetables under Stress Conditions Using Biological Nano-Selenium: A Mini-Review. Environ. Biodiv. Soil Secur. 2023, 7, 23–35. [Google Scholar] [CrossRef]
- Li, L.; Xiong, Y.; Wang, Y.; Wu, S.; Xiao, C.; Wang, S.; Cheng, S.; Cheng, H. Effect of Nano-Selenium on Nutritional Quality of Cowpea and Response of ABCC Transporter Family. Molecules 2023, 28, 1398. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.A.; Folmanis, G.E.; Tananaev, I.G. Comparative evaluation of spinach biofortification with selenium nanoparticles and ionic forms of the element. Nanotechnol. Russia 2017, 12, 569–576. [Google Scholar] [CrossRef]
- Hu, T.; Li, H.; Li, J.; Zhao, G.; Wu, W.; Liu, L.; Wang, Q.; Guo, Y. Absorption and bio-transformation of selenium nanoparticles by wheat seedlings (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 597. [Google Scholar] [CrossRef] [Green Version]
- Lei, Z.; Li, Q.; Tang, Y.; Zhang, H.; Han, C.; Wang, X.; Zhao, X.; Shi, G. Selenium enhanced nitrogen accumulation in legumes in soil with rhizobia bacteria. J. Clean. Prod. 2022, 380, 134960. [Google Scholar] [CrossRef]
- Alfthan, G.V. A micromethod for the determination of selenium in tissues and biological fluids by single-test-tube fluorimetry. Anal. Chim. Acta 1984, 165, 187–194. [Google Scholar] [CrossRef]
- AOAC. Crude protein in cereal grains and oil seeds. In Official Methods of Analysis of Association of Official Analytical Chemists, 17th ed.; Method 992.23; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Nano-Se (Se°) | Selenate (Se+6) | Selenite (Se+4) |
|---|---|---|---|---|
| Stem length (cm) | 81.3 b | 85.6 a | 81.4 b | 80.5 b |
| Nodes number up to the first pod | 3.5 b | 4.4 a | 2.6 c | 2.5 c |
| Number of fruiting nodes | 7.5 a | 8.5 a | 3.6 b | 3.4 b |
| Pod number per plant | 7.5 a | 8.5 a | 4.2 b | 4.0 b |
| Pod length (cm) | 9.0 b | 9.6 ab | 10.4 ab | 11.0 a |
| Pod width (cm) | 2.0 a | 2.0 a | 1.8 a | 2.0 a |
| Pod thickness (cm) | 3.8 b | 2.8 c | 5.5 a | 5.9 a |
| Seed number per pod | 3.2 a | 3.6 a | 3.4 a | 3.3 a |
| Seed number per plant | 16.0 b | 21.3 a | 14.4 b | 11.7 c |
| Seed weight per plant (g) | 35.7 a | 35.1 a | 24.9 b | 18.2 c |
| Weight of 1000 seeds (g) | 162.3 b | 316.0 a | 174.8 b | 164.4 b |
| Parameter | Control | Nano-Se (Se°) | Selenate (Se+6) | Selenite (Se+4) |
|---|---|---|---|---|
| Stem length (cm) | 71.4 b | 81.1 a | 77.3 a | 73.7 b |
| Nodes number up to the first pod | 2.5 b | 3.5 a | 3.2 a | 2.5 b |
| Number of fruiting nodes | 7.8 c | 8.1 b | 4.2 a | 3.8 d |
| Pod number per plant | 9.0 b | 10.2 ab | 7.2 a | 4.5 c |
| Pod length (cm) | 7.0 a | 8.0 a | 6.8 a | 7.6 a |
| Pod width (cm) | 1.6 b | 1.7 ab | 1.9 a | 1.7 ab |
| Pod thickness (cm) | 2.6 c | 1.8 d | 4.8 b | 5.1 a |
| Seed number per pod | 3.0 a | 2.7 a | 2.6 a | 2.8 a |
| Seed number per plant | 17.7 ab | 20.4 a | 15.1 b | 11.3 c |
| Seed weight per plant (g) | 21.0 b | 28.8 a | 15.7 c | 13.8 c |
| Weight of 1000 seeds (g) | 189.2 b | 260.0 a | 142.0 c | 124.6 c |
| Parameter | Control | Nano-Se (Se°) | Selenate (Se+6) | Selenite (Se+4) |
|---|---|---|---|---|
| Stem length (cm) | 80.3 a | 80.8 a | 76.2 b | 72.4 c |
| Nodes number up to the first pod | 3.4 a | 3.7 a | 3.5 a | 2.7 b |
| Number of fruiting nodes | 5.8 a | 5.3 a | 3.5 b | 3.5 b |
| Pod number per plant | 6.0 a | 6.2 a | 4.1 b | 3.6 b |
| Pod length (cm) | 10.8 a | 9.6 a | 10.8 a | 11.6 a |
| Pod width (cm) | 2.0 a | 1.9 a | 2.2 a | 2.0 a |
| Pod thickness (cm) | 2.7 b | 1.9 c | 6.0 a | 6.3 a |
| Seed number per pod | 3.7 ab | 4.1 a | 3.3 b | 3.5 ab |
| Seed number per plant | 16.2 a | 16.4 a | 12.2 b | 10.6 b |
| Seed weight per plant (g) | 28.7 a | 25.5 ab | 21.7 bc | 19.7 c |
| Weight of 1000 seeds (g) | 259.0 b | 320.0 a | 196.0 c | 178.0 c |
| Parameter | Control | Nano-Se (Se°) | Selenate (Se+6) | Selenite (Se+4) |
|---|---|---|---|---|
| Stem length (cm) | 81.0 b | 85.6 a | 77.5 bc | 73.6 c |
| Nodes number up to the first pod | 2.8 b | 4.4 a | 2.0 c | 2.4 bc |
| Number of fruiting nodes | 4.2 b | 8.5 a | 2.1 d | 3.0 c |
| Pod number per plant | 4.4 b | 8.5 a | 3.8 b | 2.5 c |
| Pod length (cm) | 12.6 a | 9.6 b | 11.2 ab | 13.3 a |
| Pod width (cm) | 2.2 a | 2.0 ab | 1.8 b | 1.9 ab |
| Pod thickness (cm) | 3.9 bc | 2.8 c | 5.8 a | 5.6 a |
| Seed number per pod | 3.2 b | 3.6 ab | 3.9 | 4.1 a |
| Seed number per plant | 10.2 b | 15.9 a | 10.8 | 11.5 b |
| Seed weight per plant (g) | 17.9 b | 35.1 a | 17.2 b | 20.0 b |
| Weight of 1000 seeds (g) | 162.3 b | 316.0 a | 155.0 b | 167 b |
| Cultivar | Se Content (mg kg−1 d.w.) | |||
|---|---|---|---|---|
| Control | Nano-Se | Sodium Selenite | Sodium Selenate | |
| Russian Black | 0.086 ± 0.004 c | 1.180 ± 0.046 b | 6.231 ± 0.012 a | 9.197 ± 0.180 b |
| Belorussian | 0.069 ± 0.004 d | 5.172 ± 0.116 a | 6.655 ± 0.585 a | 10.758 ± 0.032 ab |
| Hangdown Grünkernig | 0.154 ± 0.016 b | 0.652 ± 0.084 c | 5.754 ± 0.204 a | 11.249 ± 1.510 a |
| Dreifach Weiβe | 0.200 ± 0.009 a | 0.426 ± 0.020 d | 1.195 ± 0.254 b | 6.596 ± 0.309 c |
| Month | 2019 | 2020 | 2021 | |||
|---|---|---|---|---|---|---|
| Temperature (°C) | Precipitation (mm) | Temperature (°C) | Precipitation (mm) | Temperature (°C) | Precipitation (mm) | |
| May | 12.2 | 37.8 | 17.4 | 13 | 17.3 | 13 |
| June | 15.5 | 85.3 | 16.1 | 45 | 16.9 | 45 |
| July | 20.4 | 28.9 | 21.2 | 33 | 20.6 | 33 |
| August | 18.0 | 40.5 | 19.4 | 43 | 19.1 | 43 |
| September | 10.8 | 48.2 | 11.4 | 44 | 10.7 | 44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sindireva, A.; Golubkina, N.; Bezuglova, H.; Fedotov, M.; Alpatov, A.; Erdenotsogt, E.; Sękara, A.; Murariu, O.C.; Caruso, G. Effects of High Doses of Selenate, Selenite and Nano-Selenium on Biometrical Characteristics, Yield and Biofortification Levels of Vicia faba L. Cultivars. Plants 2023, 12, 2847. https://doi.org/10.3390/plants12152847
Sindireva A, Golubkina N, Bezuglova H, Fedotov M, Alpatov A, Erdenotsogt E, Sękara A, Murariu OC, Caruso G. Effects of High Doses of Selenate, Selenite and Nano-Selenium on Biometrical Characteristics, Yield and Biofortification Levels of Vicia faba L. Cultivars. Plants. 2023; 12(15):2847. https://doi.org/10.3390/plants12152847
Chicago/Turabian StyleSindireva, Anna, Nadezhda Golubkina, Helene Bezuglova, Mikhail Fedotov, Andrey Alpatov, Erdene Erdenotsogt, Agnieszka Sękara, Otilia Cristina Murariu, and Gianluca Caruso. 2023. "Effects of High Doses of Selenate, Selenite and Nano-Selenium on Biometrical Characteristics, Yield and Biofortification Levels of Vicia faba L. Cultivars" Plants 12, no. 15: 2847. https://doi.org/10.3390/plants12152847