
Evaluation of Trichoderma spp. on Fusarium oxysporum f. sp. asparagi and Fusarium wilt Control in Asparagus Crop



Abstract
:1. Introduction
2. Results
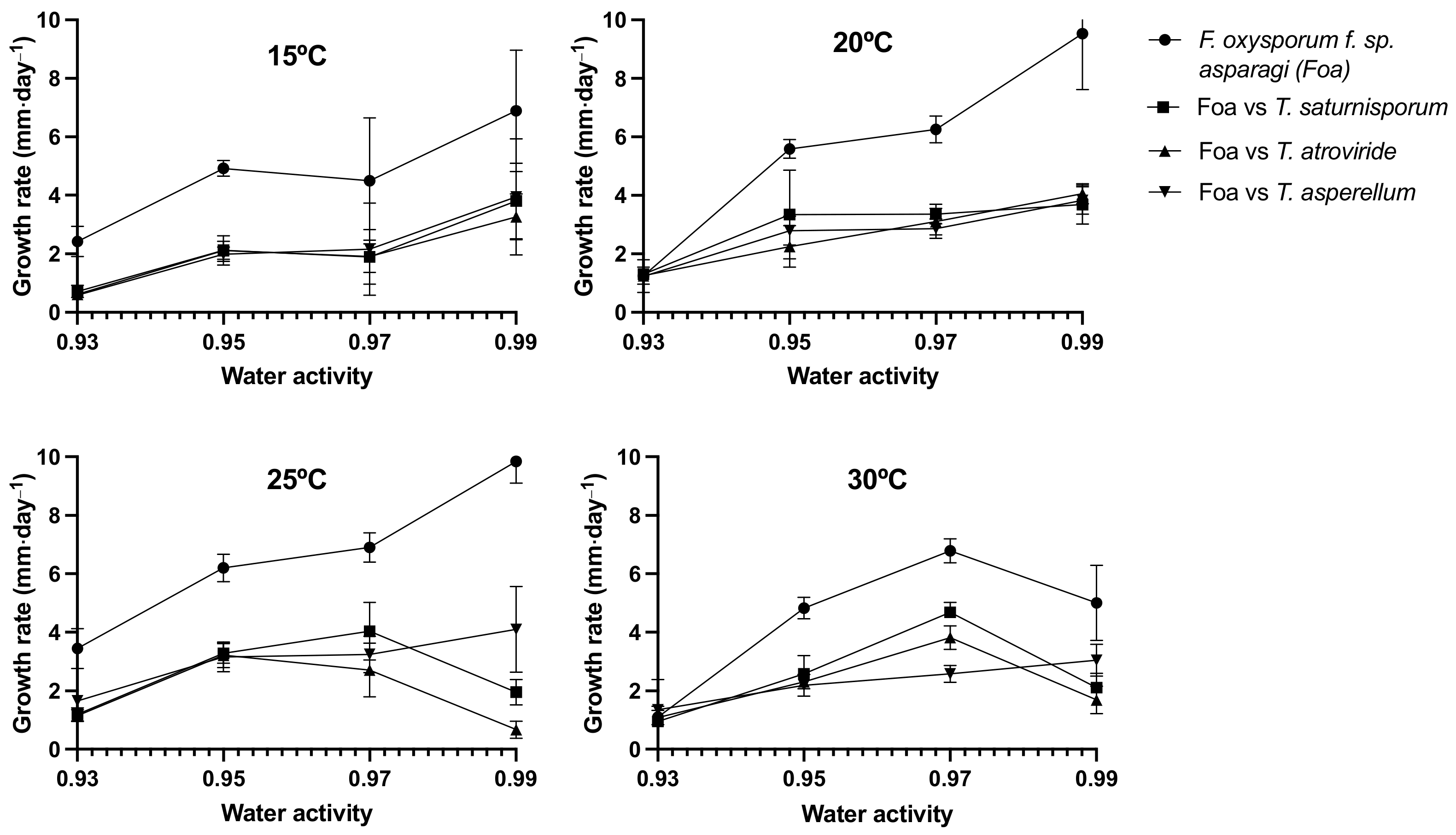
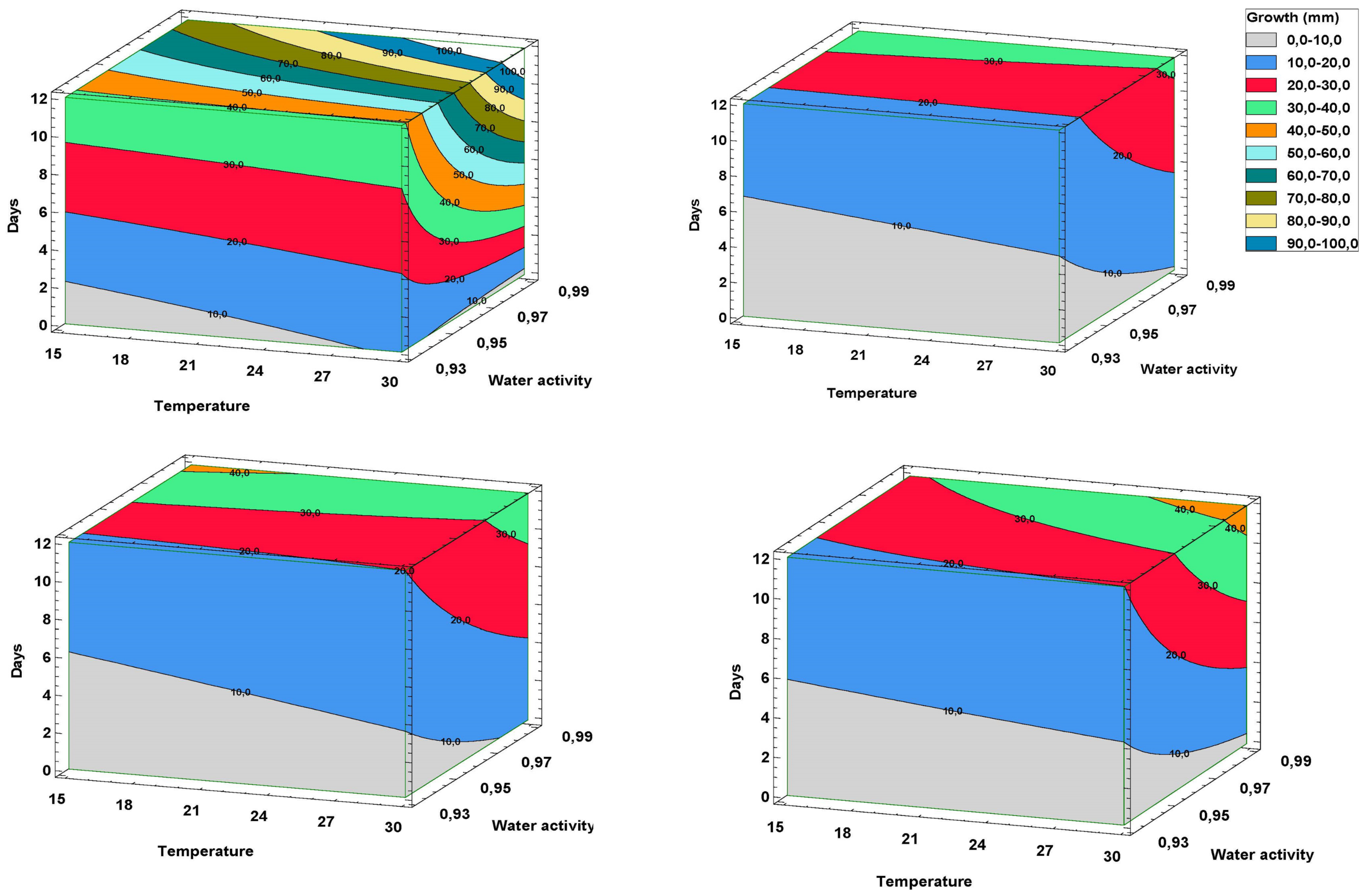
2.1. Effect of Different Environmental Variables on the Antagonistic Capacity In Vitro
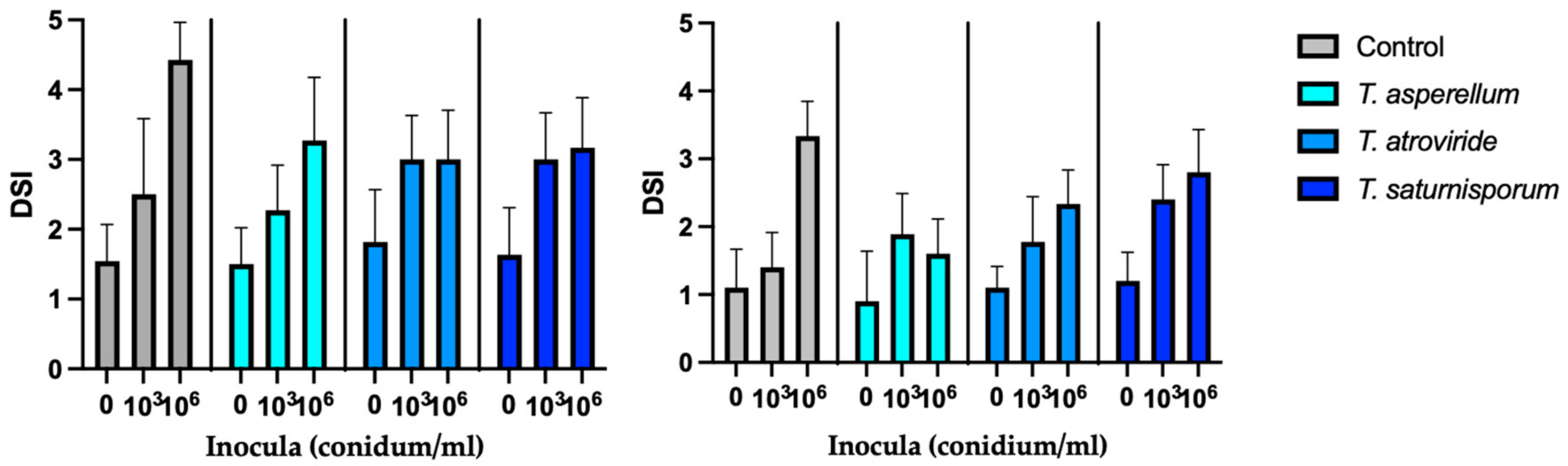
2.2. Effect of the Fungal Inoculum Density on the Efficacy of Trichoderma spp.
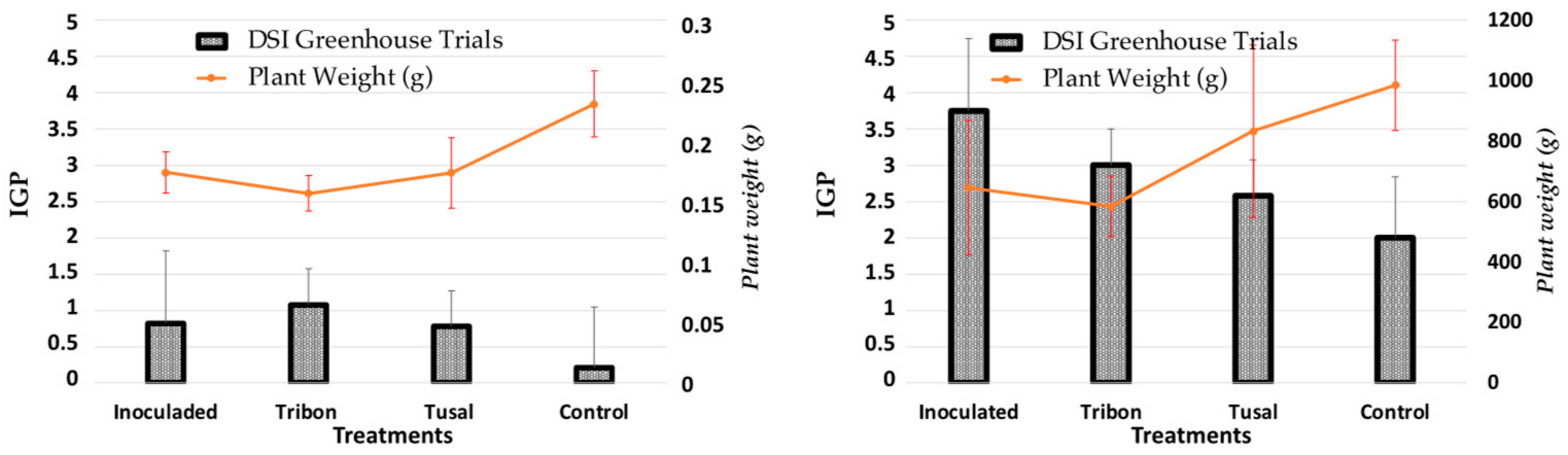
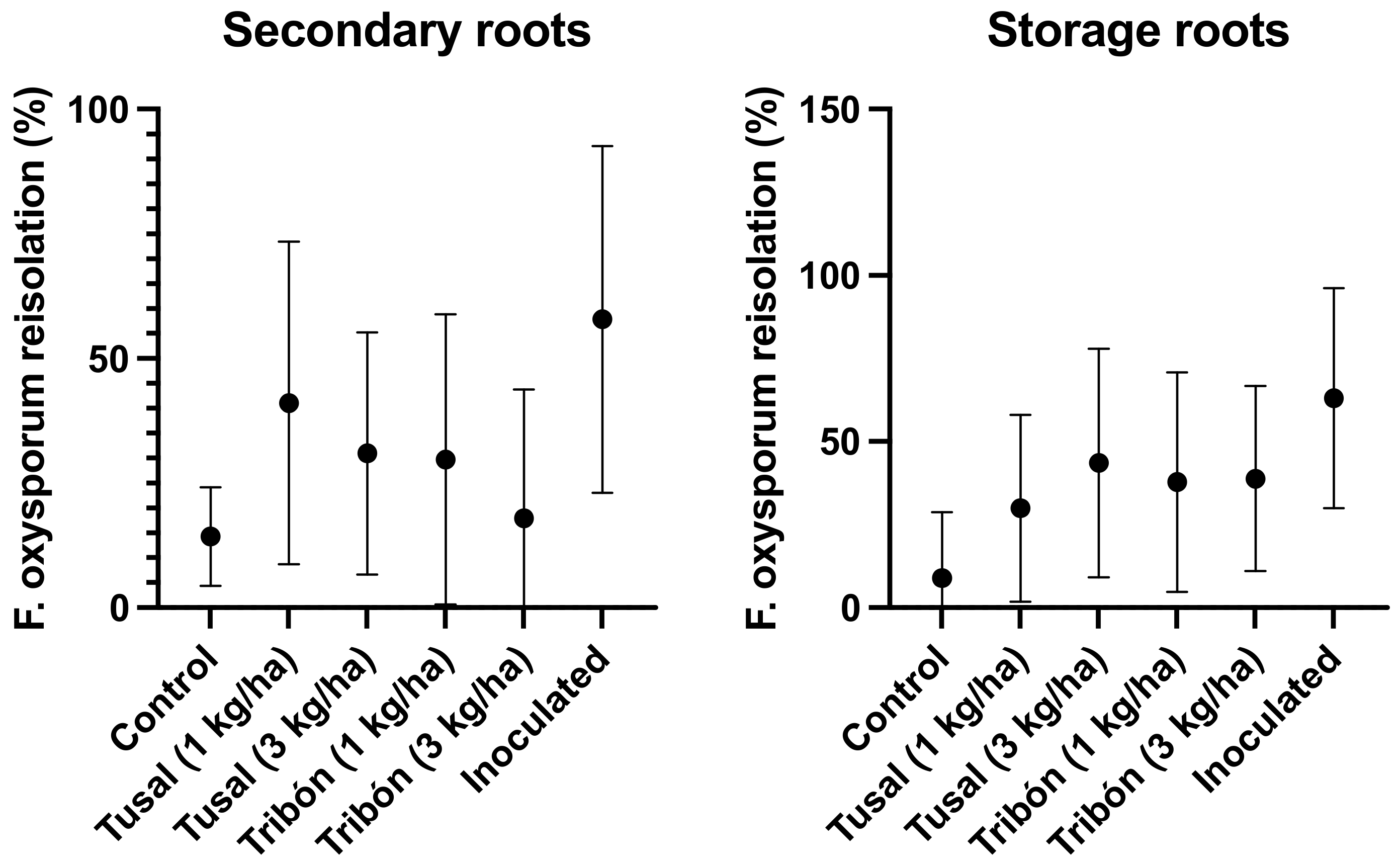
2.3. Effectiveness of Trichoderma spp. on Commercial Products under Greenhouse and Semi-Field Conditions
3. Discussion
4. Materials and Methods
4.1. Effect of Climatic Variables on the In Vitro Antagonism of Trichoderma spp. and Fusarium oxysporum f. sp. asparagi (FOA)
4.2. Antagonistic Effect of BCAs on Asparagus Seedlings Inoculated with Fusarium oxysporum f. sp. asparagi (FOA)
4.3. Efficacy of Commercial Biopesticides
4.3.1. Greenhouse Trials
4.3.2. Semi-Field Trials
4.4. Statistical Analysis of Data
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. 2021. Available online: https://www.fao.org/faostat/es/#rankings/countries_by_commodity (accessed on 14 March 2023).
- MAPA. Superficies y Producciones Anuales de Cultivos. 2022. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/superficies-producciones-anuales-cultivos/ (accessed on 14 March 2023).
- Blok, W.J.; Bollen, G.J. Fungi on roots and stem bases of asparagus in the Netherlands: Species and pathogenicity. Eur. J. Plant Pathol. 1995, 101, 15–24. [Google Scholar] [CrossRef]
- Brizuela, A.M.; De la Lastra, E.; Marín-Guirao, J.I.; Gálvez, L.; de Cara-García, M.; la Lastra, C.N.E.D.; Marín-Guirao, J.I.; Gálvez, L.; de Cara-García, M.; Capote, N.; et al. Fusarium consortium populations associated with asparagus crop in Spain and their role on field decline syndrome. J. Fungi 2020, 6, 336. [Google Scholar] [CrossRef] [PubMed]
- Andrzejak, R.; Janowska, B. The occurrence of Fusarium species in white spears of Asparagus officinalis. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12559. [Google Scholar] [CrossRef]
- Blok, W.J.; Bollen, G.J. Etiology of asparagus replant-bound early decline. Eur. J. Plant Pathol. 1996, 102, 87–98. [Google Scholar] [CrossRef]
- Fiume, F.; Fiume, G. Field response of some asparagus varieties to rust, Fusarium crown root rot, and violet root rot. Commun. Agric. Appl. Biol. Sci. 2003, 68, 659–671. [Google Scholar] [PubMed]
- Elmer, W. Asparagus decline and replant problem: A look back and a look forward at strategies for mitigating losses. Acta Hortic. 2018, 1223, 195–204. [Google Scholar] [CrossRef]
- Corpas-Hervias, C.; Melero-Vara, J.M.; Molinero-Ruiz, M.L.; Zurera-Muñoz, C.A.; Basallote-Ureba, M.J. Characterization of Isolates of Fusarium spp. obtained from Asparagus in Spain. Plant Dis. 2006, 90, 1441–1451. [Google Scholar] [CrossRef] [Green Version]
- Falavigna, A.; Alberti, P.; Casali, P.E.; Toppino, L.; Mennella, G.; Wang, H. Interspecific hybridization for asparagus breeding in Italy. Acta Hortic. 2008, 776, 291–297. [Google Scholar] [CrossRef]
- Mercati, F.; Riccardi, P.; Harkess, A.; Sala, T.; Abenavoli, M.R.; Leebens-Mack, J.; Sunseri, F. Single nucleotide polymorphism-based parentage analysis and population structure in garden asparagus, a worldwide genetic stock classification. Mol. Breed. 2015, 35, 59. [Google Scholar] [CrossRef]
- Apaza, W.; Talledo, A.; Casas, A. Reaction of 10 Asparagus cultivars to infested soil with Fusarium oxysporum f. sp. asparagi and Meloidogyne incognita. Acta Hortic. 2018, 1223, 233–238. [Google Scholar] [CrossRef]
- Plath, S.; Krämer, R.; Lantos, E.; Nothnagel, T. Breeding programs to transmit Asparagus virus 1 resistance. Acta Hortic. 2018, 1223, 17–24. [Google Scholar] [CrossRef]
- Kathe, L.; Krämer, R.; Budahn, H.; Pillen, K.; Rabenstein, F.; Nothnagel, T. Development of a bioassay to assess resistance to Fusarium oxysporum (Schlecht.) in asparagus (Asparagus officinalis L.). J. Phytopathol. 2019, 167, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Jacobi, J.; Budahn, H.; Nothnagel, T.; König, J. Studies on the identification of resistance to Fusarium oxysporum (Schlecht.) in different genetic backgrounds of Asparagus officinalis (L.) and its defense responses. Horticulturae 2023, 9, 158. [Google Scholar] [CrossRef]
- Reid, T.C.; Hausbeck, M.K. Use of fungicides and biological controls in the suppression of Fusarium crown and root rot of asparagus under greenhouse and growth chamber conditions. Plant Dis. 2002, 86, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Ferreira, F.V.; Musumeci, M.A. Trichoderma as biological control agent: Scope and prospects to improve efficacy. World J. Microbiol. Biotechnol. 2021, 37, 90. [Google Scholar] [CrossRef]
- Akram, S.; Ahmed, A.; He, P.; He, P.; Liu, Y.; Wu, Y.; Munir, S.; He, Y. Uniting the role of endophytic fungi against plant pathogens and their interaction. J. Fungi 2023, 9, 72. [Google Scholar] [CrossRef]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Barbosa, J.Z.; Hungria, M.; Prior, S.A.; Moura, M.C.; Poggere, G.; Motta, A.C.V. Improving yield and health of legume crops via co-inoculation with rhizobia and Trichoderma: A global meta-analysis. Appl. Soil Ecol. 2022, 176, 104493. [Google Scholar] [CrossRef]
- Çakar, G.; Tozlu, E. The biological control of Fusarium oxysporum, the causal agent of potato rot. Gesunde Pflanz. 2022, 74, 305–315. [Google Scholar] [CrossRef]
- Jadhav, S.R.; Ingle, Y.V.; Paithanar, D.H.; Bhange, G.P. Laboratory evaluation of fungicides and microbial antagonists against Fusarium solani and Rhizoctonia bataticola using damping-off and root rot disease in acid lime seedlings. J. Plant Dis. Sci. 2022, 17, 8–12. [Google Scholar] [CrossRef]
- Patil, S.V.; Raja, J. Antagonism of Trichoderma species against major soil borne plant pathogens. J. Plant Dis. Sci. 2022, 17, 39–43. [Google Scholar] [CrossRef]
- Rabiey, M.; Hailey, L.E.; Roy, S.R.; Grenz, K.; Al-Zadjali, M.A.S.; Barrett, G.A.; Jackson, R.W. Endophytes vs tree pathogens and pests: Can they be used as biological control agents to improve tree health? Eur. J. Plant Pathol. 2019, 155, 711–729. [Google Scholar] [CrossRef] [Green Version]
- Howell, C.R. Mechanisms employed by Trichoderma species in the Biological Control of Plant Diseases: The History and Evolution of Current Concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segaran, G.; Sathiavelu, M. Biocatalysis and agricultural biotechnology fungal endophytes: A potent biocontrol agent and a bioactive metabolites reservoir. Biocatal. Agric. Biotechnol. 2019, 21, 101–284. [Google Scholar] [CrossRef]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Benhamou, N.; Kapulnik, Y.; Chet, I. Concomitant induction of systemic resistance to Pseudomonas syringae pv. lachrymans in cucumber by Trichoderma asperellum (T-203) and accumulation of phytoalexins. Appl. Environ. Microbiol. 2003, 69, 7343–7353. [Google Scholar] [CrossRef] [Green Version]
- Belhadj Benyahia, F.; Kthiri, Z.; Hamada, W.; Boureghda, H. Trichoderma atroviride induces biochemical markers associated with resistance to Fusarium culmorum, the main crown rot pathogen of wheat in Algeria. Biocontrol. Sci. Technol. 2021, 31, 357–372. [Google Scholar] [CrossRef]
- Karuppiah, V.; He, A.; Lu, Z.; Wang, X.; Li, Y.; Chen, J. Trichoderma asperellum GDFS1009—Mediated maize resistance against Fusarium graminearum stalk rot and mycotoxin degradation. Biol. Control 2022, 174, 105026. [Google Scholar] [CrossRef]
- Qin, C.; Hu, Z.; Xiong, Y.; Chen, M.; Li, C.; Ding, W. A new sesquiterpene derivative from the mangrove endophytic fungus Trichoderma harzianum (Strain No. R1). Chem. Nat. Compd. 2021, 57, 267–269. [Google Scholar] [CrossRef]
- Singh, G.; Tiwari, A.; Choudhir, G.; Kumar, A.; Kumar, S.; Hariprasad, P.; Sharma, S. Deciphering the role of Trichoderma sp. bioactives in combating the wilt causing cell wall degrading enzyme polygalacturonase produced by Fusarium oxysporum: An in-silico approach. Microb. Pathog. 2022, 168, 105610. [Google Scholar] [CrossRef]
- Harman, G.E.; Lorito, M.; Lynch, J.M. Uses of Trichoderma spp. to alleviate or remediate soil and water pollution. Adv. Appl. Microbiol. 2004, 56, 313–330. [Google Scholar]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In vivo study of Trichoderma-pathogen-plant interactions, using constitutive and inducible green fluorescent protein reporter systems. Appl. Environ. Microbiol. 2004, 70, 3073–3081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunner, K.; Zeilinger, S.; Ciliento, R.; Woo, S.L.; Lorito, M.; Kubicek, C.P.; Mach, R.L. Improvement of the fungal biocontrol agent Trichoderma atroviride to enhance both antagonism and induction of plant systemic disease resistance. Appl. Environ. Microbiol. 2005, 71, 3959–3965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, F.; Razdan, V.K.; Mohiddin, F.A.; Bhat, K.A.; Banday, S. Potential of Trichoderma species as biocontrol agents of soil borne fungal propagules. J. Phytol. 2010, 2, 38–41. [Google Scholar]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carretero, F.; Ruiz, L.A.; Berenguel, M.; Martínez, F.D.; Marín, F.F.; Martinez, M.; Yau, J.A.; Santos, M. Trichoderma saturnisporum, nuevo agente de control biológico y bioestimulante sobre diferentes cultivos hortícolas. Phytohemeroteca 2013, 246, 32–39. [Google Scholar]
- Li, Y.-T.; Hwang, S.-G.; Huang, Y.-M.; Huang, C.-H. Effects of Trichoderma asperellum on nutrient uptake and Fusarium wilt of tomato. Crop Prot. 2018, 110, 275–282. [Google Scholar] [CrossRef]
- He, A.; Liu, J.; Wang, X.; Zhang, Q.; Song, W.; Chen, J. Soil application of Trichoderma asperellum GDFS1009 granules promotes growth and resistance to Fusarium graminearum in maize. J. Integr. Agric. 2019, 18, 599–606. [Google Scholar] [CrossRef]
- Quiroga-Rojas, L.I.; Ruiz-Quiñones, N.; Muñoz-Motta, G.; Lozano-Tovar, M.D. Rhizosphere microorganism, potential antagonists of Fusarium sp. causing agent of root rot in passion fruit (Passiflora edulis Sims). Acta Agron. 2012, 61, 244–250. [Google Scholar]
- Mannai, S.; Boughalleb, M.N. In vitro and in planta potential effect of some indigenous antagonists against Fusarium and pythiaceous species associated with peach seedlings decline. Egypt. J. Biol. Pest Control 2022, 32, 60. [Google Scholar] [CrossRef]
- Cuervo-Parra, J.A.; Pérez España, V.H.; Zavala-González, E.A.; Peralta-Gil, M.; Aparicio Burgos, J.E.; Romero-Cortes, T. Trichoderma asperellum strains as potential biological control agents against Fusarium verticillioides and Ustilago maydis in maize. Biocontrol. Sci. Technol. 2022, 32, 624–647. [Google Scholar] [CrossRef]
- Davari, M.; Ezazi, R. Mycelial inhibitory effects of antagonistic fungi, plant essential oils and propolis against five phytopathogenic Fusarium species. Arch. Microbiol. 2022, 204, 480. [Google Scholar] [CrossRef]
- Rawat, K.; Tripathi, S.B.; Kaushik, N.; Bishnu, B.M. Management of bakanae disease of rice using biocontrol agents and insights into their biocontrol mechanisms. Arch. Microbiol. 2022, 204, 401. [Google Scholar] [CrossRef]
- Tiwari, R.; Chandra, K.; Shukla, S.K.; Jaiswal, V.P.; Amaresan, N.; Srivastava, A.K.; Tiwari, R.K. Interference of bio-control Trichoderma to enhance physical and physiological strength of sugarcane during Pokkah boeng infection. World J. Microbiol. Biotechnol. 2022, 38, 139. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytother. Res. 2020, 34, 2835–2842. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L. Fungi vs fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Elazouni, I.; Abdel, S.; Amira, A. Microbial efficacy as biological agents for potato enrichment as well as bio-controls against wilt disease caused by Ralstonia solanacearum. World J. Microbiol. Biotechnol. 2019, 35, 30. [Google Scholar] [CrossRef]
- Zaim, S.; Bekkar, A.A.; Belabid, L. Efficacy of Bacillus subtilis and Trichoderma harzianum combination on chickpea Fusarium wilt caused by F. oxysporum f. sp. ciceris. Arch. Phytopathol. Plant Prot. 2018, 51, 217–226. [Google Scholar] [CrossRef]
- Yendyo, S.; Ramesh, G.C.; Pandey, B.R. Evaluation of Trichoderma spp. Pseudomonas fluorescence and Bacillus subtilis for biological control of Ralstonia wilt of tomato. F1000Research 2017, 6, 2028. [Google Scholar] [CrossRef]
- González-Domínguez, E.; Molinero-Ruíz, M.L.; Sánchez-Jimenez, V.; Vela-Delgado, M.D.; Basallote-Ureba, M.J. Utilización de Trichoderma spp. en el control de la podredumbre de raíz y cuello de espárrago. In Proceedings of the XV Congreso de la Sociedad Española de Fitopatología, Vitoria-Gasteiz, Spain, 1 October 2010; Volume 195. Available online: http://hdl.handle.net/10261/28906 (accessed on 12 May 2023).
- Arriola, L.L.; Hausbeck, M.K.; Rogers, J.; Safir, G.R. The effect of Trichoderma harzianum and arbuscular mycorrhizae on Fusarium root rot in asparagus. Hort. Technol. 2000, 10, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Pérez, E.; Molinero-Ruíz, M.L.; Melero-Vara, J.M.; Basallote-Ureba, M.J. Selection of potential antagonists against Asparagus crown and root rot caused by Fusarium spp. Commun. Agric. Appl. Biol. Sci. 2008, 73, 203–206. [Google Scholar] [PubMed]
- Rao, Y.; Zeng, L.; Jiang, H.; Mei, L.; Wang, Y. Trichoderma atroviride LZ42 releases volatile organic compounds promoting plant growth and suppressing Fusarium wilt disease in tomato seedlings. BMC Microbiol. 2022, 22, 88. [Google Scholar] [CrossRef]
- Diánez Martínez, F.; Santos, M.; Carretero, F.; Marín, F. Trichoderma saturnisporum, a new biological control agent. J. Sci. Food Agric. 2016, 96, 1934–1944. [Google Scholar] [CrossRef] [PubMed]
- Royse, D.; Ries, S. The influence of fungi isolated from peach twigs on the pathogenicity of Cytospora cincta. Phytopathology 1978, 68, 603–607. [Google Scholar] [CrossRef]
- Ezziyyani, M.; Requena, M.E.; Egea-Gilabert, C.; Candela, M.E. Biological control of Phytophthora root rot of pepper using Trichoderma harzianum and Streptomyces rochei in combination. J. Phytopathol. 2007, 155, 342–349. [Google Scholar] [CrossRef]
- Andrade, H.P.; Luna-Cruz, A.; Osorio-Hernández, E.; Molina-Gayosso, E.; Landero-Valenzuela, N.; Barrales-Cureño, H. Antagonism of Trichoderma spp. vs fungi associated with wilting of chilli. Rev. Mex. Cienc. Agríc. 2019, 10, 1259–1272. [Google Scholar] [CrossRef] [Green Version]
- Romero-Arenas, O.; Amaro, J.L.; Damián, M.A.; de Ita, M.A.V.; Rivera, A.H.M. Trichoderma spp. biopreparates for the biological control of Phytophthora capsici in the tomato crop of Puebla, Mexico. ITEA 2017, 113, 313–324. [Google Scholar] [CrossRef]
- Bell, D.K.; Wells, H.D.; Markham, C.R. In Vitro Antagonism of Trichoderma species Against Six Fungal Plant Pathogens. Phytopathology 1982, 72, 379–382. [Google Scholar] [CrossRef]
- Stankovic, S.; Levic, J.; Petrovic, T.; Logrieco, A.; Moretti, A. Pathogenicity and mycotoxin production by Fusarium proliferatum isolated from onion and garlic in Serbia. Eur. J. Plant Pathol. 2007, 118, 165–172. [Google Scholar] [CrossRef]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium Species. An Illustrated Manual for Identification; The Pennsylvania State University Press: Madison, WI, USA, 1983; p. 193. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006; p. 388. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biocontrol Agents | Temperature | Water Activity | |||
|---|---|---|---|---|---|
| 0.99 | 0.97 | 0.95 | 0.93 | ||
| T. saturnisporum | 15 | 1 | 1.5 | 2.6 | 3 |
| 20 | 1.6 | 1 | 1.1 | 2 | |
| 25 | 1 | 1.8 | 2 | 3 | |
| p-Value = 0.00 | 30 | 1 | 1 | 1 | 1.8 |
| T. atroviride | 15 | 1 | 1 | 2 | 3.8 |
| 20 | 1 | 1 | 1 | 3.5 | |
| 25 | 1 | 1 | 2 | 4 | |
| p-Value = 0.00 | 30 | 3 | 3.6 | 2 | 4 |
| T. asperellum | 15 | 1.6 | 2.8 | 3.6 | 4 |
| 20 | 2 | 3 | 3 | 4 | |
| 25 | 1 | 2.6 | 3 | 4 | |
| p-Value = 0.01 | 30 | 1 | 1 | 3 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brizuela, A.M.; Gálvez, L.; Arroyo, J.M.; Sánchez, S.; Palmero, D. Evaluation of Trichoderma spp. on Fusarium oxysporum f. sp. asparagi and Fusarium wilt Control in Asparagus Crop. Plants 2023, 12, 2846. https://doi.org/10.3390/plants12152846
Brizuela AM, Gálvez L, Arroyo JM, Sánchez S, Palmero D. Evaluation of Trichoderma spp. on Fusarium oxysporum f. sp. asparagi and Fusarium wilt Control in Asparagus Crop. Plants. 2023; 12(15):2846. https://doi.org/10.3390/plants12152846
Chicago/Turabian StyleBrizuela, Alexandri María, Laura Gálvez, Juan Manuel Arroyo, Silvia Sánchez, and Daniel Palmero. 2023. "Evaluation of Trichoderma spp. on Fusarium oxysporum f. sp. asparagi and Fusarium wilt Control in Asparagus Crop" Plants 12, no. 15: 2846. https://doi.org/10.3390/plants12152846