
Effect of Sodium Selenate and Selenocystine on Savoy Cabbage Yield, Morphological and Biochemical Characteristics under Chlorella Supply
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
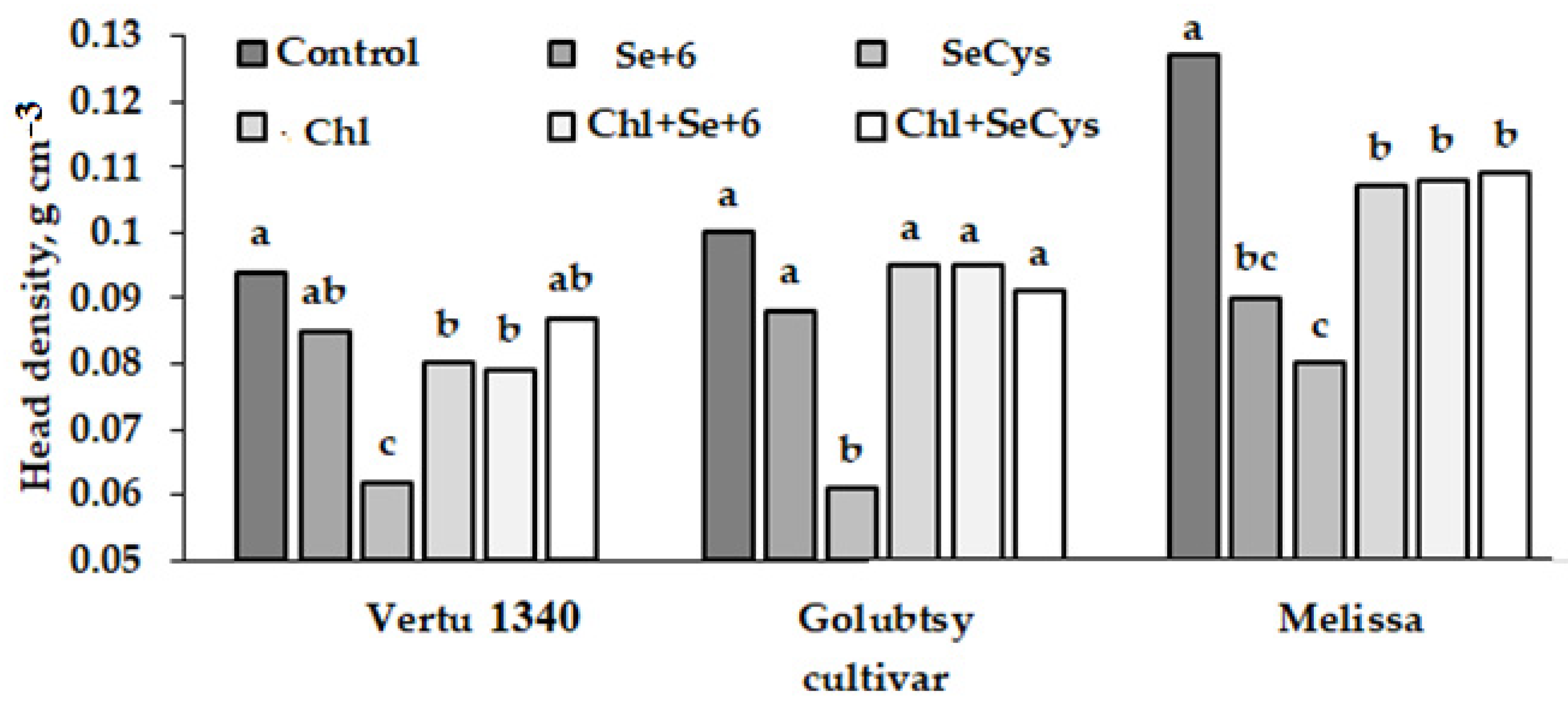
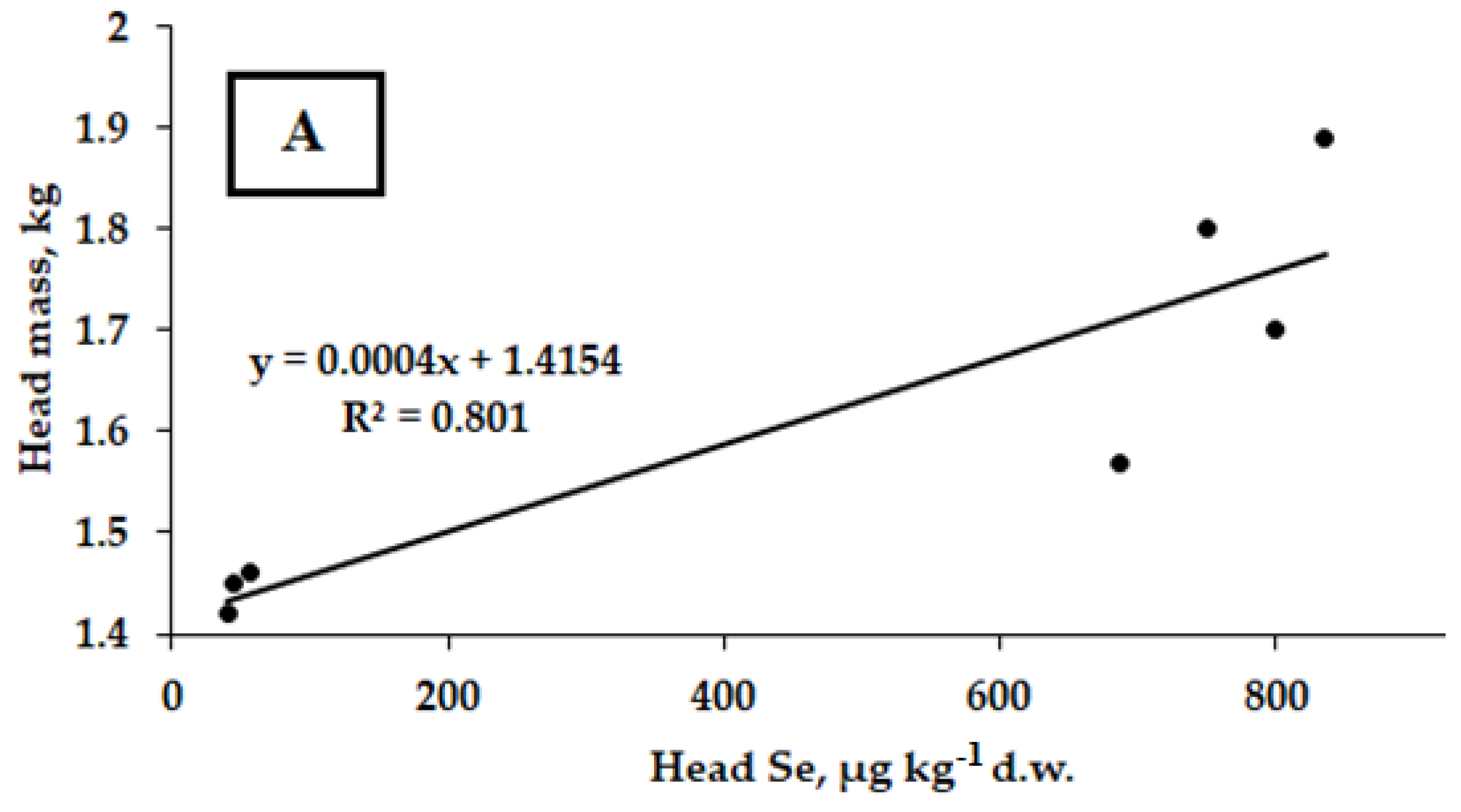
2.1. Yield and Morphological Characteristics of Savoy Cabbage
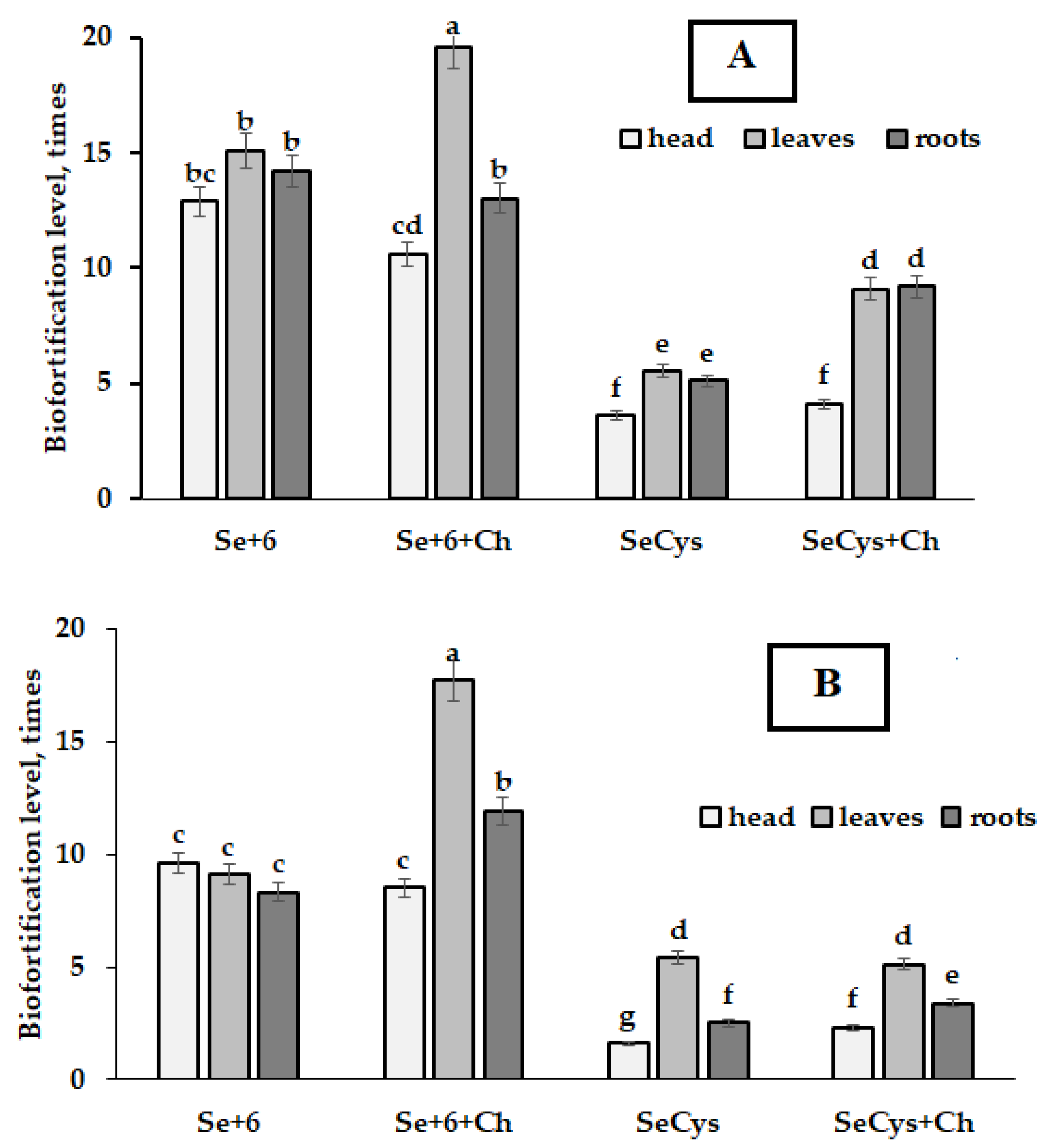
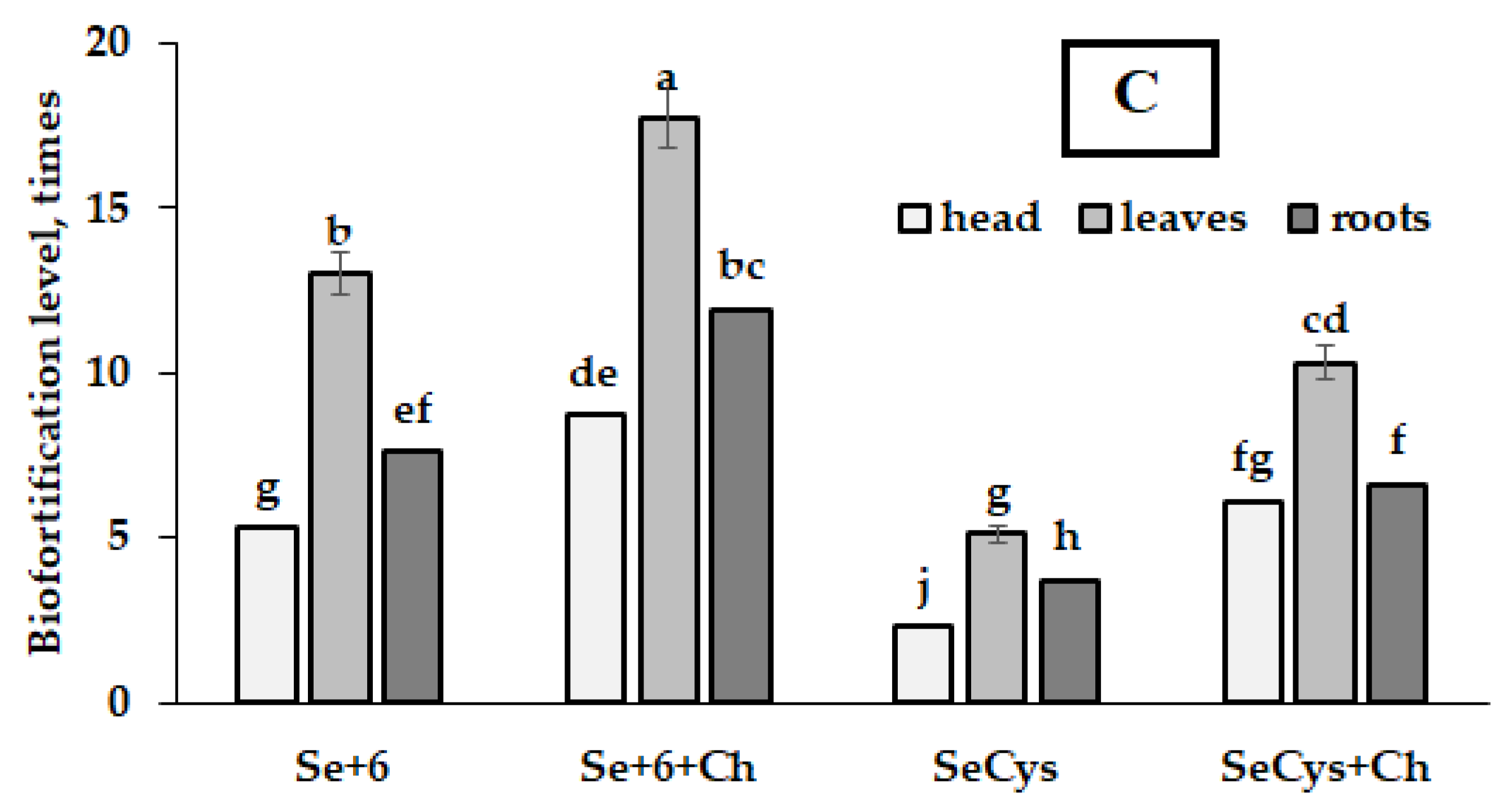
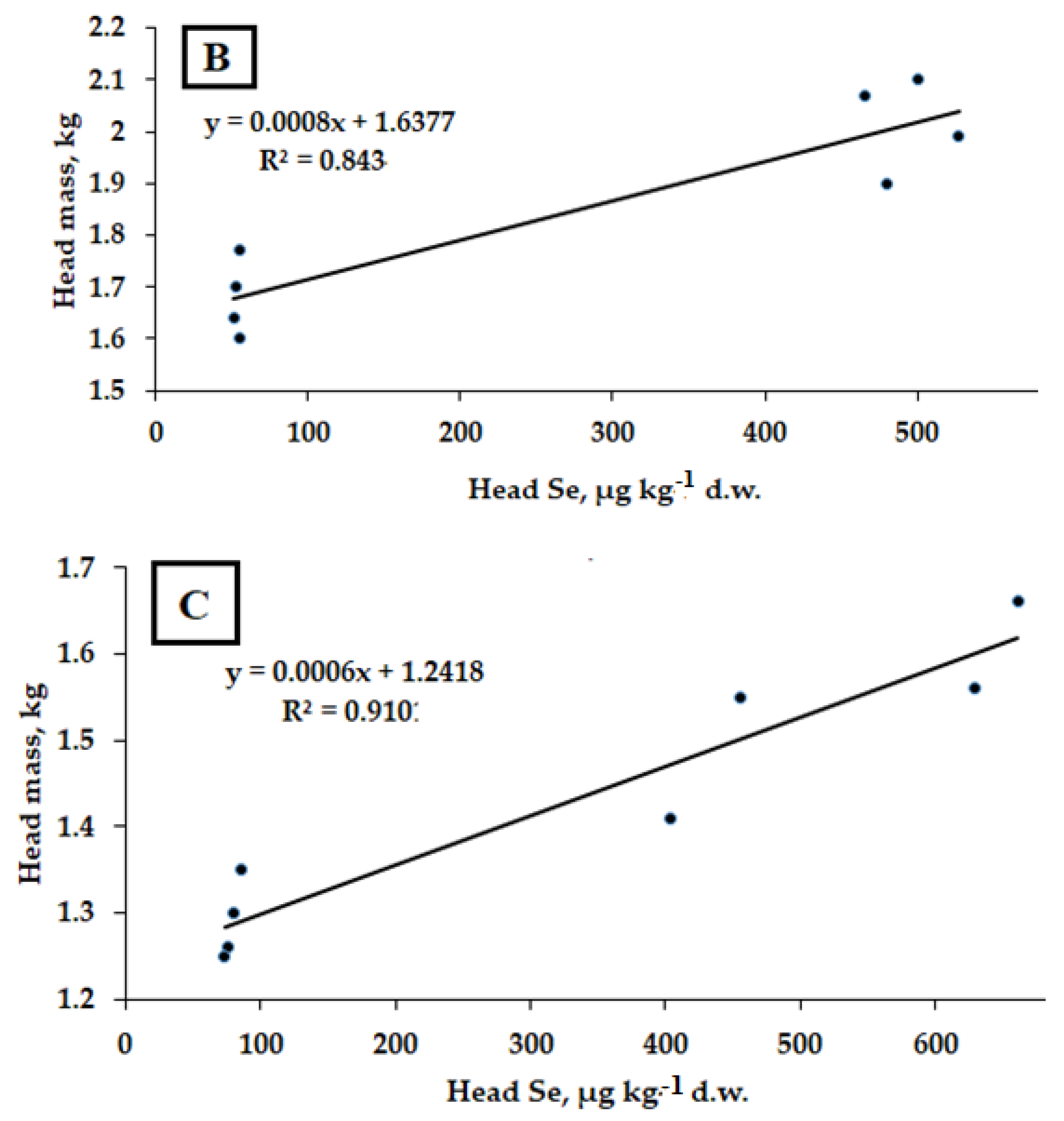
2.2. Selenium Accumulation
2.3. Antioxidants
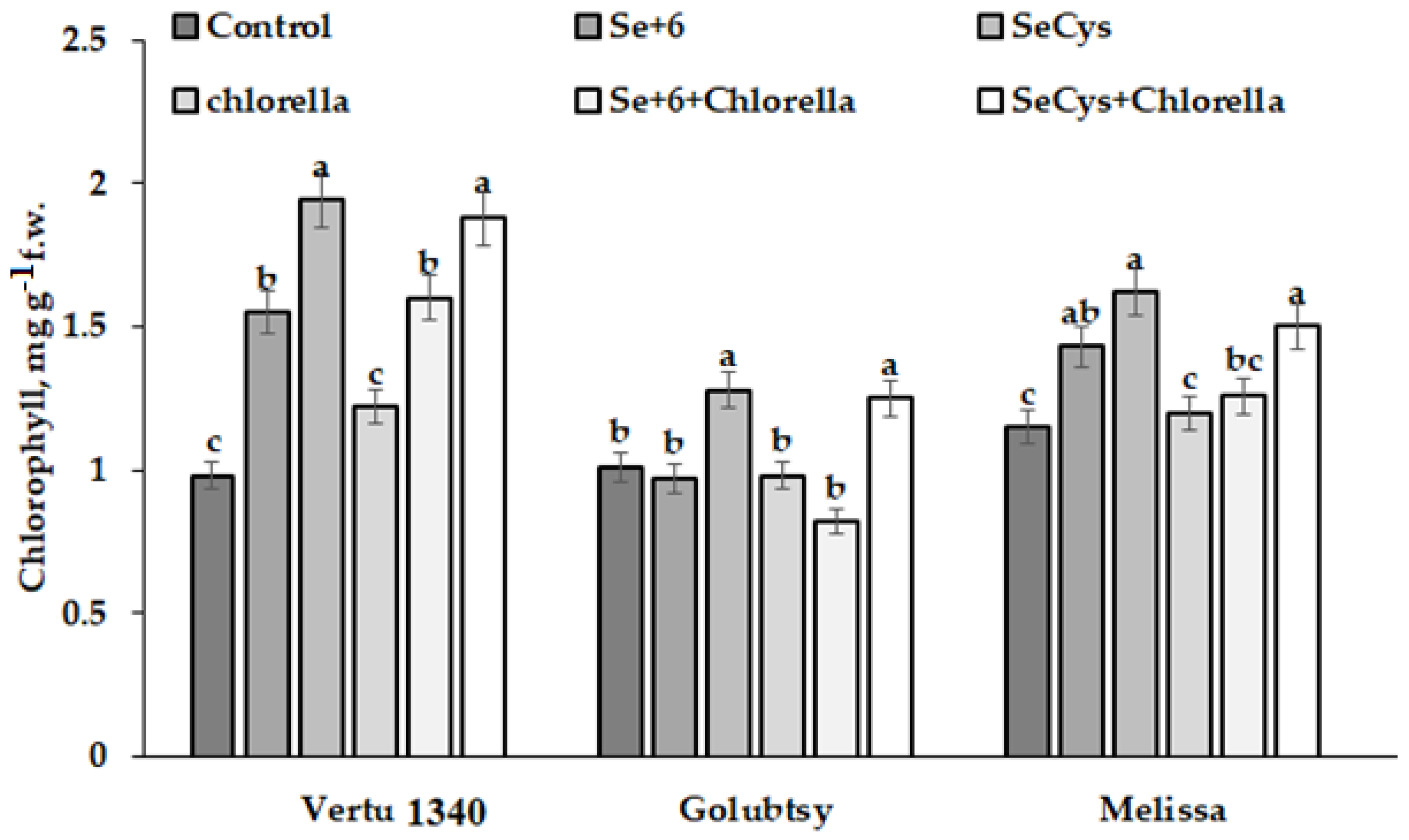
2.3.1. Photosynthetic Pigments
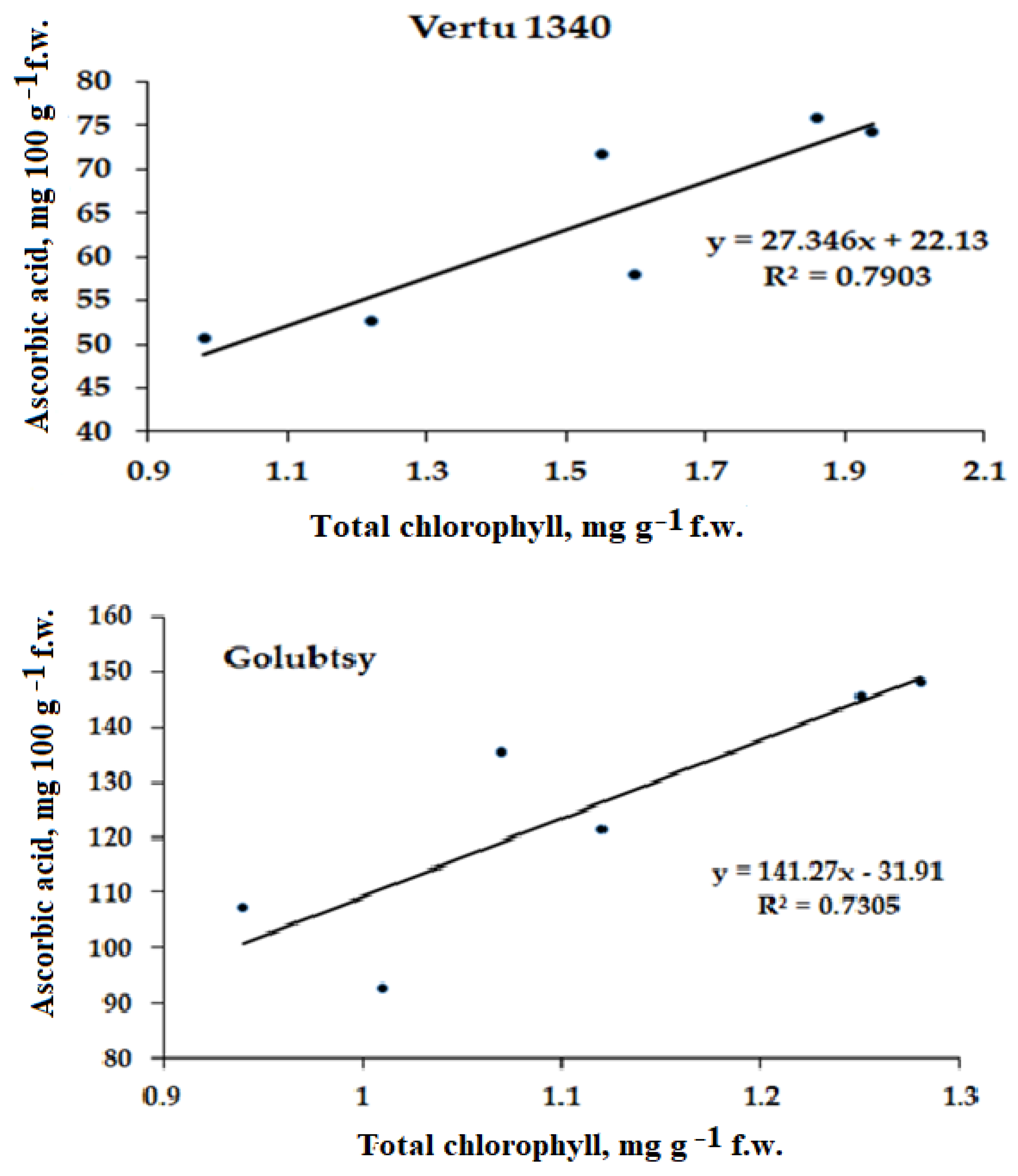
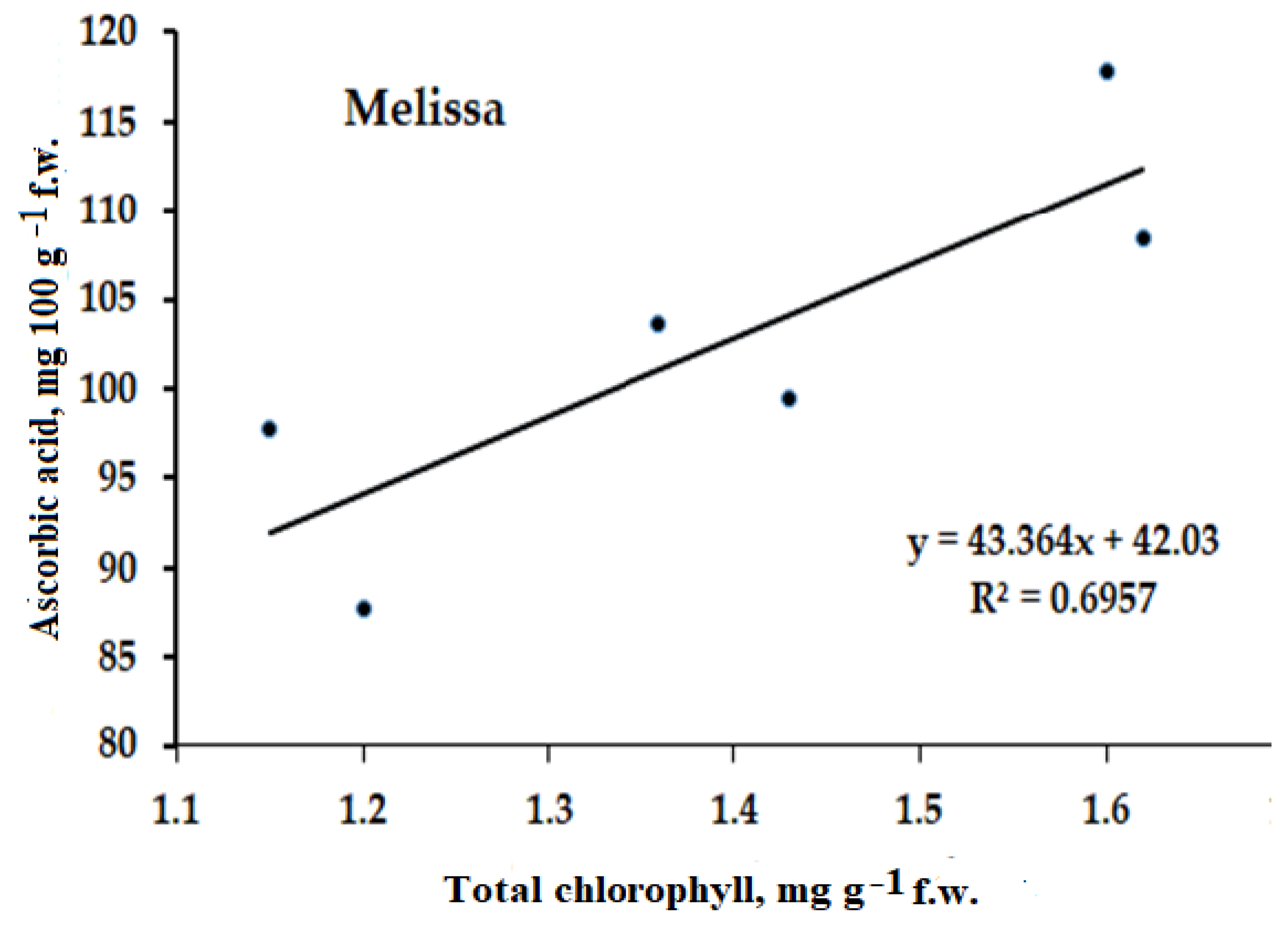
2.3.2. Ascorbic Acid
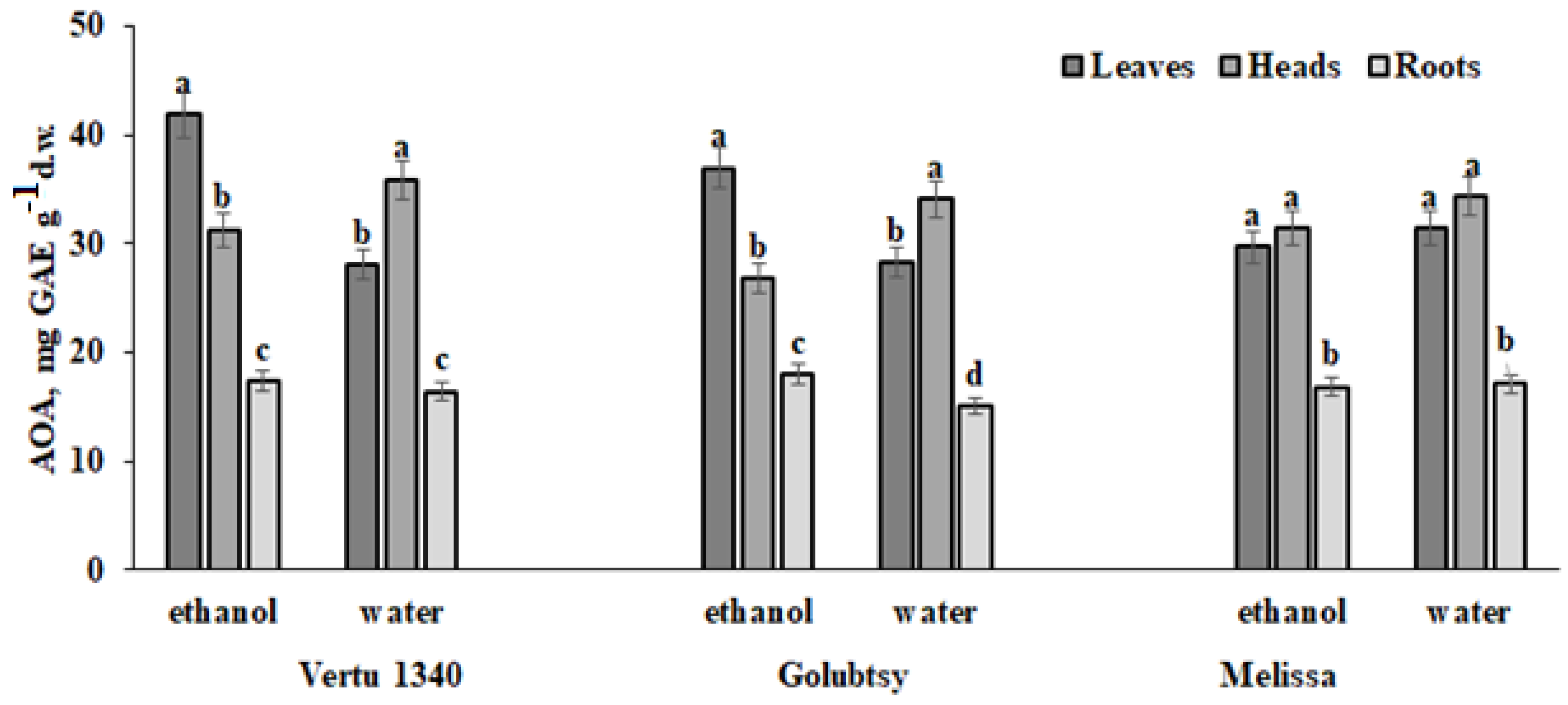
2.3.3. Total Antioxidant Activity (AOA)
2.4. Dry Matter, Nitrates, Total Dissolved Solids (TDS), Carbohydrates
3. Material and Methods
3.1. Growing Conditions and Experimental Protocol
3.2. Sample Preparation
3.3. Head Density
3.4. Dry Matter
3.5. Nitrates
3.6. Ascorbic Acid
3.7. Photosynthetic Pigments
3.8. Total Polyphenols (TP)
3.9. Antioxidant Activity (AOA)
3.10. Sugars
3.11. Selenium
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Golubkina, N.A.; Kharchenko, V.A.; Caruso, G. Selenium: Prospects of functional food production with high antioxidant activity. In Reference Series in Phytochemistry. Plant Antioxidants and Health; Ekiert, H., Ramawat, K.G., Arora, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Alfthan, G.; Eurola, M.; Ekholm, P.; Venäläinen, E.-R.; Root, T.; Korkalainen, K.; Hartikainen, H.; Salminee, P.; Hietaniemi, V.; Aspila, P.; et al. Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: From deficiency to optimal selenium status of the population. J. Trace Elem. Med. Biol. 2015, 31, 142–147. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, R.; Regni, L.; Falcinelli, B.; Mattioli, S.; Benincasa, P.; Dal Bosco, A.; Pacheco, P.; Proietti, P.; Troni, E.; Santi, C.; et al. Current knowledge on selenium biofortification to improve the nutraceutical profile of food: A comprehensive review. J. Agric. Food Chem. 2020, 68, 4075–4097. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Arroyo, I.S.; Dangi, S.R.; Zambrano, M.C. Continued selenium biofortification of carrots and broccoli grown in soils once amended with Se-enriched S. pinnata. Front. Plant Sci. 2016, 7, 1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavon, M.; Pilon-Smits, E.A. The fascinating facets of plant selenium accumulation—Biochemistry, physiology, evolution and ecology. New Phytol. 2017, 213, 1582–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, H.J.; Carmona, V.V.; Dutra, A.F.; Filho, A.B.C. Growth and physiological responses of cabbage cultivars biofortified with inorganic selenium fertilizers. Sci. Hortic. 2022, 302, 111154. [Google Scholar] [CrossRef]
- Lyi, S.M.; Heller, L.I.; Rutzke, M.; Welch, R.M.; Kochian, L.V.; Li, L. Molecular and biochemical characterization of the selenocysteine Se-methyltransferase gene and Se-methylselenocysteine synthesis in broccoli. Plant Physiol. 2005, 138, 409–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Wang, Z.; Zhang, L.; Peng, J.; Huang, T.; Yang, X.; Jeong, B.R.; Yang, Q. Seleno-amino acids in vegetables: A review of their forms and metabolism. Front. Plant Sci. 2022, 13, 804368. [Google Scholar] [CrossRef]
- Wiesner-Reinhold, M.; Schreiner, M.; Baldermann, S.; Schwarz, D.; Hanschen, F.S.; Kipp, A.P.; Rowan, D.D.; Bentley-Hewitt, K.L.; McKenzie, M.J. Mechanisms of selenium enrichment and measurement in Brassicaceous vegetables, and their application to human health. Front. Plant Sci. 2017, 8, 1365. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, M.; Matich, A.; Hunter, D.; Esfandiari, A.; Trolove, S.; Chen, R.; Lill, R. Selenium application during radish (Raphanus sativus) plant development alters glucosinolate metabolic gene expression and results in the production of 4-(methylseleno)but-3-enyl glucosinolate. Plants 2019, 8, 427. [Google Scholar] [CrossRef] [Green Version]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Sarkar, S.; Ahmad, Z.; Vemuri, H.; Garai, S.; Mondal, M.; Bhatt, R.; et al. Selenium biofortification: Roles, mechanisms, responses and prospects. Molecules 2021, 26, 881. [Google Scholar] [CrossRef]
- Muñoz, F.F.; Stoffel, M.M.; Céccoli, G.; Trod, B.S.; Daurelio, L.D.; Bouzo, C.A.; Guevara, M.G. Improving the foliar biofortification of broccoli with selenium without commercial quality losses. Crop Sci. 2021, 61, 4218–4228. [Google Scholar] [CrossRef]
- Antoshkina, M.S.; Golubkina, N.A.; Bondareva, L.L. Effect of foliar sodium selenate bio-fortification on cauliflower yield, nutritional value and antioxidant status. Veg. Crops Russ. 2020, 3, 63–68. (In Russian) [Google Scholar] [CrossRef]
- Utoiu, E.; Oancea, A.; Gaspar, A.; Seciu, A.-M.; Ștefan, L.M.; Coroiu, V.; Crăciunescu, O.; Badiu, C.D.; Oancea, F. Selenium biofortification treatment of cauliflower enhances their content in chemopreventive compounds and in vitro antitumoral activity. Sci.Bull. Ser. F Biotechnol. 2017, 21, 33–40. [Google Scholar]
- Antoshkina, M.; Golubkina, N.; Sekara, A.; Tallarita, A.; Caruso, G. Effects of selenium application on biochemical characteristics and biofortification level of kohlrabi (Brassica oleracea L. var. gongylodes) produce. Front. Biosci. 2021, 26, 533–542. [Google Scholar] [CrossRef]
- Golob, A.; Novak, T.; Maršić, N.K.; Šircelj, H.; Stibilj, V.; Jerše, A.; Kroflič, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes and increases their contents in tubers. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef]
- Kikkert, J.; Berkelaar, E. Plant uptake and translocation of inorganic and organic forms of selenium. Arch. Environ. Contam. Toxicol. 2013, 65, 458–465. [Google Scholar] [CrossRef]
- Kaur, S.; Kaur, N.; Siddique, K.H.; Nayyar, H. Beneficial elements for agricultural crops and their functional relevance in defence against stresses. Arch. Agron. Soil Sci. 2016, 62, 905–920. [Google Scholar] [CrossRef]
- Ogra, Y.; Ogihara, Y.; Anan, Y. Comparison of the metabolism of inorganic and organic selenium species between two selenium accumulator plants, garlic and Indian mustard. Metallomics 2017, 9, 61–68. [Google Scholar] [CrossRef]
- Eich-Greatorex, S.; Sogn, T.A.; Øgaard, A.F.; Aasen, I. Plant availability of inorganic and organic selenium fertiliser as influenced by soil organic matter content and pH. Nutr. Cycl. Agroecosyst. 2007, 79, 221–231. [Google Scholar] [CrossRef]
- Golubkina, N.; Zamana, S.; Seredin, T.; Poluboyarinov, P.; Sokolov, S.; Baranova, H.; Krivenkov, L.; Pietrantonio, L.; Caruso, G. Effect of selenium biofortification and beneficial microorganism inoculation on yield, quality and antioxidant properties of shallot bulbs. Plants 2019, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ali, F.; Wang, M.; Dinh, Q.T.; Zhou, F.; Bañuelos, G.S.; Liang, D. Understanding boosting selenium accumulation in wheat (Triticum aestivum L.) following foliar selenium application at different stages, forms, and doses. Environ. Sci. Pollut. Res. Int. 2020, 27, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Irish, K.P.; Harvey, M.A.; Harris, H.H.; Aart, M.G.M.; Chan, C.X.; Erskine, P.D.; Van der Ent, A. Micro-analytical and molecular approaches for understanding the distribution, biochemistry, and molecular biology of selenium in (hyperaccumulator) plants. Planta 2023, 257, 2. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhu, Y.; Lin, Z.Q.; Banuelos, G.; Li, W.; Yin, X. A novel selenocystine-accumulating plant in selenium-mine drainage area in Enshi, China. PLoS ONE 2013, 8, e65615. [Google Scholar] [CrossRef]
- Poluboyarinov, P.A.; Moiseeva, I.J.; Mikulyak, N.I.; Golubkina, N.A.; Kaplun, A.P. New synthesis of cysteine, selenocystine and their derivatives. Chem. Chem. Technol. 2022, 65, 19–29. (In Russian) [Google Scholar]
- Poluboyarinov, P.A.; Elistratov, D.G.; Shvets, V.I. Metabolism and mechanism of toxicity of selenium-containing supplements used for optimization of the human selenium status. Fine Chem. Technol. 2019, 14, 5–24. (In Russian) [Google Scholar] [CrossRef]
- Xiao, T.; Boada, R.; Llugany, M.; Valiente, M. Co-application of Se and a biostimulant at different wheat growth stages: Influence on grain development. Plant Physiol. Biochem. 2021, 160, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.; Sekara, A.; Tallarita, A.; Torino, V.; Sellitto, V.M.; Stoleru, V.; Brîndușa, S.H.; Caruso, G. Formulates based on microalgae and cyanobacteria applied to herbaceous crop systems. A review. Italus Hortus 2022, 29, 94–114. [Google Scholar] [CrossRef]
- Li, J.; Lens, P.N.L.; Ferrer, I.; Laing, G.D. Evaluation of selenium-enriched microalgae produced on domestic wastewater as biostimulant and biofertilizer for growth of selenium-enriched crops. J. Appl. Phycol. 2021, 33, 3027–3039. [Google Scholar] [CrossRef]
- Germ, M.; Stibilj, V. Selenium and plants. Acta Agric. Slov. 2007, 89, 65–71. [Google Scholar] [CrossRef]
- Hasanuzzamam, M.; Bhuyan, M.H.M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Mahmud, J.A.; Naha, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, Y.; Shi, G.; Zhang, X. Selenium delays tomato fruit ripening by inhibiting ethylene biosynthesis and enhancing the antioxidant defense system. Food Chem. 2017, 219, 179–184. [Google Scholar] [CrossRef]
- Duang, M.Y.; Cheng, S.R.; Lu, R.H.; Lai, R.F.; Zheng, A.X.; Ashraf, U.; Fan, P.S.; Du, B.; Luo, H.W.; Tang, X.R. Effect of foliar sodium selenate on leaf senescence of fragrant rice in South China. Appl. Ecol. Environ. Res. 2019, 17, 3343–3351. [Google Scholar] [CrossRef]
- Mechora, Š.; Germ, M.; Stibilj, V. Selenium compounds in selenium-enriched cabbage. Pure Appl. Chem. 2012, 84, 259–268. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An Overview of Selenium Uptake, Metabolism, and Toxicity in Plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef] [Green Version]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the new plant growth biostimulants based on amino acids on yield and grain quality of winter wheat. Molecules 2018, 23, 470. [Google Scholar] [CrossRef] [Green Version]
- El-Bassiony, A.M.; Mahmoud, S.H.; El-Sawy, S.M.; Shedeed, S.I. Stimulation of Growth and Productivity of Onion Plants by Selenium and Growth Active Substances. Middle East J. Agric. Res. 2020, 9, 637–645. [Google Scholar] [CrossRef]
- Al-Taie, A.A.G.; Aboohanah, M.A.; Issa, F.H. Effect of cysteine in stimulating some active compounds in garlic callus under different light spectra in vitro. IOP Conf. Ser. Earth Environ. Sci. 2021, 910, 012119. [Google Scholar] [CrossRef]
- Lee, S.-M.; Ryu, C.-M. Algae as new kids in beneficial plant microbiome. Front. Plant Sci. 2021, 12, 599742. [Google Scholar] [CrossRef] [PubMed]
- Hajnal-Jafari, T.; Seman, V.; Stamenov, D.; Đurić, S. Effect of Chlorella vulgaris on growth and photosynthetic pigment content in Swiss chard (Beta vulgaris L. subsp. cicla). Pol. J. Microbiol. 2020, 69, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 1991, 83, 702–708. [Google Scholar] [CrossRef]
- Turakainen, M.; Hartikainen, H.; Seppänen, M.M. Effects of selenium treatments on potato (Solanum tuberosum L.) growth and concentrations of soluble sugars and starch. J. Agric. Food Chem. 2004, 52, 5378–5382. [Google Scholar] [CrossRef]
- Breznik, B.; Germ, M.; Gaberscik, A.; Kreft, I. Combined effects of elevated UV-B radiation and the addition of selenium on common (Fagopyrum esculentum Moench) and tartary [Fagopyrum tataricum (L.) Gaertn.] buckwheat. Photosynthetica 2005, 43, 583–589. [Google Scholar] [CrossRef]
- Lyons, G.H.; Genc, Y.; Soole, K.; Stangoulis, J.; Liu, F.; Graham, R. Selenium increases seed production in Brassica. Plant Soil 2009, 318, 73–80. [Google Scholar] [CrossRef]
- Smirnoff, N.; Wheeler, G.L. Ascorbic acid in plants: Biosynthesis and function. CRC Crit. Rev. Biochem. Mol. Biol. 2000, 35, 291–314. [Google Scholar] [CrossRef]
- Golubkina, N.; Kekina, H.; Caruso, G. Yield, quality and antioxidant properties of Indian mustard (Brassica juncea L.) in response to foliar biofortification with selenium and iodine. Plants 2018, 7, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ríos, J.J.; Rosales, M.A.; Blasco, B.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. Sci. Hortic. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Sabatino, L.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C. Effect of selenium enrichment and type of application on yield, functional quality and mineral composition of curly endive grown in a hydroponic system. Agronomy 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Newman, R.; Waterland, N.; Moon, Y.; Tou, J.C. Selenium biofortification of agricultural crops and effects on plant nutrients and bioactive compounds important for human health and disease prevention—A review. Plant Foods Hum. Nutr. 2019, 74, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Bybordi, A. Effect of ascorbic acid and silicium on photosynthesis, antioxidant enzyme activity, and fatty acid contents in canola exposure to salt stress. J. Integr. Agr. 2012, 11, 1610–1620. [Google Scholar] [CrossRef]
- Ivanov, V.N. The role of ascorbic acid in photosynthesis. Biochemistry 2014, 79, 282–289. [Google Scholar] [CrossRef]
- Golubkina, N.; Molchanova, A.; Shevchuk, O.; Khlipenko, L.; Logvinenko, L. Biochemical characteristics of prospect herbs of the Nikita Botanic Gardens collection. Quest. Biol. Med. Pharm. Chem. 2018, 21, 3–9. (In Russian) [Google Scholar]
- Podsedek, A.; Sosnowska, D.; Redzynia, M.; Anders, B. Antioxidant capacity and content of Brassica oleracea dietary antioxidants. Int. J. Food Sci. Technol. 2006, 41 (Suppl. S1), 49–58. [Google Scholar] [CrossRef]
- Fernández-León, A.M.; Lozano, M.; González, D.; Ayuso, M.C.; Fernández-León, M.F. Bioactive compounds content and total antioxidant activity of two Savoy cabbages. Czech J. Food Sci. 2014, 32, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical activity and polyphenol composition of local Brassicaceae edible varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- VanEtten, C.H.; Daxenbichler, M.E.; Kwolek, W.F.; Williams, P.H. Communications distribution of glucosinolates in the pith, cambial-cortex, and leaves of the head in cabbage, Brassica oleracea L. J. Agric. Food Chem. 1979, 27, 648–650. [Google Scholar] [CrossRef]
- Rios, J.J.; Blasco, B.; Rosales, M.A.; Sanchez-Rodriguez, E.; Leyva, R.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Response of nitrogen metabolism in lettuce plants subjected to different doses and forms of selenium. J. Sci. Food Agric. 2010, 90, 1914–1919. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.; Lei, B.; Cheng, R.; Wang, Y.; Li, T.; Yang, Q. Selenium distribution and nitrate metabolism in hydroponic lettuce (Lactuca sativa L.): Effects of selenium forms and light spectra. J. Integr. Agric. 2020, 19, 133–144. [Google Scholar] [CrossRef]
- Hajiboland, R.; Sadeghzade, N. Effect of selenium on CO2 and NO3 assimilation under low and adequate nitrogen supply in wheat (Triticum aestivum L.). Photosynthetica 2014, 52, 501–510. [Google Scholar] [CrossRef]
- Ruiz, J.M.; Rivero, R.M.; Romero, L. Comparative effect of Al, Se and Mo toxicity on NO3 assimilation in sunflower (Helianthus annuus L.) plants. J. Environ. Manag. 2007, 83, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Bian, Z.; Yang, Q.; Wang, J.; Cheng, R.; Kun, L.; Liu, W.; Zhang, Y.; Fang, H.; Tong, Y. The positive function of selenium supplementation on reducing nitrate accumulation in hydroponic lettuce (Lactuca sativa L.). J. Integr. Agric 2018, 17, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M.; Abbasifar, B.; ValizadehKaji, A. Nitrate accumulation and physicochemical characteristics of lettuce as affected by sodium selenite and synthesized selenium nanoparticles. Int. J. Veg. Sci. 2022, 28, 570–582. [Google Scholar] [CrossRef]
- Munshi, C.B.; Mindy, N.I. Glycoalkaloid and nitrate content of potatoes as affected by method of selenium application. Biol. Trace Elem. Res. 1992, 33, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.A.; Kosheleva, O.V.; Krivenkov, L.V.; Dobrutskaya, H.G.; Nadezhkin, S.; Caruso, G. Intersexual differences in plant growth, yield, mineral composition and antioxidants of spinach (Spinacia oleracea L.) as affected by selenium form. Sci. Hortic. 2017, 225, 350–358. [Google Scholar] [CrossRef]
- Radovich, T.J.K.; Kleinhenz, M.D. Rapid estimation of cabbage head volume across a population varying in head shape: A test of two geometric formulae. Horttechnology 2004, 14, 388–391. [Google Scholar] [CrossRef] [Green Version]
- Kharchenko, V.A.; Moldovan, A.I.; Amagova, Z.A.; Matsadze, V.K.; Golubkina, N.A.; Caruso, G. Effect of sodium selenate foliar supplementation on Cryptotaenia japonica and Petroselinum crispum nutritional characteristics and seed quality. Veg. Crop. Russ. 2022, 4, 5–10. [Google Scholar] [CrossRef]
- AOAC Association Official Analytical Chemists. The Official Methods of Analysis of AOAC International; 22 Vitamin C; AOAC: Rockville, MD, USA, 2012. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Golubkina, N.A.; Kekina, H.G.; Molchanova, A.V.; Antoshkina, M.S.; Nadezhkin, S.M.; Soldatenko, A.V. Plants Antioxidants and Methods of Their Determination; Infra M: Moscow, Russian, 2020. [Google Scholar] [CrossRef]
- Swamy, P.M. Laboratory Manual on Biotechnology; Rastogi Publications: Meerut, India, 2008; p. 617. [Google Scholar]
- Alfthan, G.V. A micromethod for the determination of selenium in tissues and biological fluids by single-test-tube fluorimetry. Anal. Chim. Acta 1984, 165, 187–194. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Head Diameter (cm) | Head Height (cm) | Mean Head Weight (kg) | Leaves Weight (kg) | Yield (kg) | ||
|---|---|---|---|---|---|---|---|
| Marketable | Non- Marketable | Total | |||||
| Vertu 1340 | |||||||
| Control | 18.3 a | 10.8 c | 1.42 b | 0.69 c | 34.6 c | 5.2 a | 39.8 b |
| Se+6 | 19.0 a | 12.2 bc | 1.57 ab | 0.91 b | 39.2 bc | 4.8 a | 44.0 b |
| SeCys2 | 21.2 a | 15.8 a | 1.84 a | 1.19 a | 47.0 ab | 4.5 a | 51.5 a |
| Chlorella | 19.1 a | 11.9 bc | 1.46 b | 1.08 ab | 35.5 c | 5.4 a | 40.9 b |
| Se+6+ Ch | 20.6 a | 13.4 ab | 1.89 a | 1.28 a | 48.3 a | 4.6 a | 52.9 a |
| SeCys2+ Ch | 18.5 a | 11.5 bc | 1.44 b | 1.21 a | 35.6 c | 4.7 a | 40.3 b |
| M ± SD | 10.5 ± 1.2 | 12.6 ± 1.8 | 1.60 ± 0.21 | 1.06 ± 0.22 | 40.4 ± 6.1 | 4.9 ± 0.4 | 45.3 ± 5.9 |
| CV (%) | 11.3 | 14.2 | 13.1 | 20.7 | 15.1 | 8.2 | 13.1 |
| Golubtsy | |||||||
| Control | 19.3 b | 11.6 a | 1.77 ab | 0.96 c | 44.7 b | 4.9 a | 49.6 ab |
| Se+6 | 20.8 ab | 12.5 a | 1.99 ab | 1.06 bc | 51.1 ab | 4.6 a | 55.7 ab |
| SeCys2 | 24.7 a | 13.8 a | 2.16 a | 1.38 ab | 56.0 a | 4.5 a | 60.5 a |
| Chlorella | 18.6 b | 11.9 a | 1.64 b | 1.27 ab | 41.1 b | 4.8 a | 45.9 b |
| Se+6+ Ch | 20.1 ab | 12.9 a | 2.07 a | 1.49 a | 53.5 a | 4.5 a | 58.0 a |
| SeCys2+ Ch | 19.7 b | 12.4 a | 1.84 a | 1.19 b | 46.8 b | 4.7 a | 51.5 ab |
| M ± SD | 20.5 ± 2.2 | 12.5 ± 0.8 | 1.91 ± 0.20 | 1.22 ± 0.20 | 48.9 ± 5.6 | 4.5 ± 0.2 | 53.5 ± 5.5 |
| CV (%) | 10.6 | 6.2 | 10.5 | 16.4 | 11.5 | 4.4 | 10.3 |
| Melissa | |||||||
| Control | 15.0 b | 10.5 b | 1.26 c | 0.81 c | 30.3 b | 5.0 a | 35.3 b |
| Se+6 | 17.1 ab | 12.8 ab | 1.41 abc | 1.05 b | 34.8 ab | 4.7 a | 39.5 ab |
| SeCys2 | 19.2 a | 13.2 a | 1.63 ab | 1.23 a | 40.9 a | 4.7 a | 45.6 a |
| Chlorella | 15.8 ab | 12.1 ab | 1.35 bc | 1.09 ab | 32.9 b | 4.9 a | 37.8 b |
| Se+6+ Ch | 16.9 ab | 12.9 a | 1.66 a | 1.28 a | 42.0 a | 4.5 a | 46.5 a |
| SeCys2+ Ch | 16.2 ab | 11.9 ab | 1.43 abc | 1.14 ab | 35.3 ab | 4.7 a | 40.0 ab |
| M ± SD | 16.7 ± 1.4 | 12.2 ± 1.0 | 1.50 ± 0.10 | 1.10 ± 0.17 | 36.0 ± 4.6 | 4.8 ± 0.1 | 40.8 ± 4.4 |
| CV (%) | 8.4 | 8.2 | 6.7 | 15.4 | 12.7 | 2.1 | 10.8 |
| Chl a | Chl b | Car | Chl a/b | Chl/car | |
|---|---|---|---|---|---|
| Vertu 1340 | |||||
| Control | 0.59 c | 0.39 c | 0.08 c | 1.51 c | 12.25 a |
| Se+6 | 1.01 ab | 0.54 b | 0.13 b | 1.87 ab | 11.92 a |
| SeCys2 | 1.2 a | 0.74 a | 0.17 a | 1.62 bc | 11.41 a |
| Chlorella | 0.8 b | 0.42 c | 0.10 c | 1.90 a | 12.20 a |
| Se+6+ Chlorella | 1.02 ab | 0.58 b | 0.15 ab | 1.76 abc | 10.67 a |
| SeCys2+ Chlorella | 1.2 a | 0.68 a | 0.16 a | 1.76 abc | 11.75 a |
| M ± SD | 0.97 ± 0.24 | 0.56 ± 0.14 | 0.13 ± 0.03 | 1.74 ± 0.15 | 11.77 ± 0.59 |
| CV, % | 24.7 | 25.0 | 23.1 | 8.6 | 5.0 |
| Golubtsy cv. | |||||
| Control | 0.53 b | 0.48 ab | 0.10 b | 1.10 a | 10.1 ab |
| Se+6 | 0.52 b | 0.55 bc | 0.10 b | 1.16 a | 9.70 ab |
| SeCys2 | 0.71 a | 0.57 a | 0.15 a | 1.25 a | 8.53 b |
| Chlorella | 0.47 b | 0.47 b | 0.08 b | 1.00 a | 11.75 a |
| Se+6+ Chlorella | 0.53 b | 0.59 c | 0.09 b | 1.10 a | 9.11 ab |
| SeCys2+ Chlorella | 0.70 a | 0.55 a | 0.13 a | 1.27 a | 9.62 ab |
| M ± SD | 0.56 ± 0.10 | 0.48 ± 0.05 | 0.11 ± 0.02 | 1.15 ± 0.10 | 9.45 ± 1.10 |
| CV, % | 17.8 | 10.3 | 18.2 | 8.7 | 11.6 |
| Melissa cv. | |||||
| Control | 0.57 c | 0.58 b | 0.10 c | 1.00 b | 11.50 a |
| Se+6 | 0.82 ab | 0.61 ab | 0.12 bc | 1.34 a | 11.92 a |
| SeCys2 | 0.95 a | 0.67 a | 0.18 a | 1.42 a | 9.00 b |
| Chlorella | 0.70 b | 0.50 b | 0.14 b | 1.40 a | 8.57 b |
| Se+6+ Chlorella | 0.65 bc | 0.71 a | 0.09 c | 1.07 b | 14.00 a |
| SeCys2+ Chlorella | 0.88 a | 0.72 a | 0.17 a | 1.42 a | 8.82 b |
| M ± SD | 0.76 ± 0.12 | 0.59 ± 0.04 | 0.13 ± 0.03 | 1.28 ± 0.19 | 10.38 ± 2.19 |
| CV (%) | 15.8 | 6.7 | 22.5 | 14.8 | 21.1 |
| Vertu 1340 | Golubsty | Melissa | ||||
|---|---|---|---|---|---|---|
| Leaves | Head | Leaves | Head | Leaves | Head | |
| Control | 50.7 b | 42.5 b | 92.8 d | 46.5 c | 97.7 a | 50.5 ab |
| Se+6 | 71.7 a | 47.2 ab | 135.5 ab | 66.4 a | 99.4 a | 51.1 ab |
| SeCys2 | 74.2 a | 54.5 a | 148.3 a | 67.1 a | 108.4 a | 59.9 a |
| Chlorella | 52.7 b | 43.7 b | 107.4 cd | 50.2 bc | 87.7 a | 46.9 b |
| Se+6+ Chlorella | 57.9 b | 46.2 ab | 121.3 bc | 58.0 ab | 103.7 a | 57.6 a |
| SeCys2+ Chlorella | 75.8 a | 50.0 ab | 145.5 a | 66.6 a | 117.8 a | 59.3 a |
| M ± SD | 63.8 ± 11.4 | 45.7 ± 7.1 | 125.1 ± 22.1 | 59.1 ± 9.1 | 102.5 ± 10.2 | 54.2 ± 5.4 |
| CV (%) | 17.9 | 15.5 | 17.7 | 15.4 | 10.0 | 10.0 |
| Leaves | Heads | Roots | ||||
|---|---|---|---|---|---|---|
| Ethanol | Water | Ethanol | Water | Ethanol | Water | |
| Vertu | ||||||
| control | 40.4 a | 36.5 a | 37.1 a | 37.7 a | 16 b | 21.1 a |
| Se+6 | 44.4 a | 25.9 c | 30.2 b | 35.6 a | 14.9 b | 15.8 b |
| SeCys2 | 41.4 a | 25.1 c | 29 b | 37.6 a | 17.9 ab | 16.9 ab |
| Chlorella | 37.1 a | 30.3 abc | 30.5 b | 35.5 a | 16.5 b | 16.3 b |
| Se+6+ Ch | 46.3 a | 27 bc | 34.3 ab | 36 a | 18.2 a | 16.3 b |
| SeCys2+ Ch | 40.5 a | 32.3 ab | 32.6 ab | 34.6 a | 19.5 a | 16.8 b |
| M ± SD | 41.7 ± 3.3 a | 29.5 ± 4.4 b | 32.3 ± 3.0 a | 36.2 ± 1.2 a | 17.2 ± 1.7 a | 16.5 ± 0.5 a |
| Golubtsy | ||||||
| control | 29.6 b | 31.7 a | 27.4 a | 36.2 ac | 16.5 b | 24.4 a |
| Se+6 | 37.2 a | 26.9 a | 24.5 a | 42.2 a | 18.2 b | 15.6 bc |
| SeCys2 | 39.5 a | 26.1 a | 24.7 a | 33.3 bc | 17.9 b | 14.5 c |
| Chlorella | 34.8 ab | 26.6 a | 28.1 a | 32.1 b | 17.5 b | 14.2 c |
| Se+6+ Ch | 35.3 a | 29.8 a | 26.4 a | 32.1 b | 17.5 b | 15.8 bc |
| SeCys2+ Ch | 38.3 a | 31.9 a | 30.6 a | 30.6 b | 18.7 b | 15.2 c |
| M ± SD | 35.8 ± 3.5 a | 28.8 ± 2.6 b | 27.0 ± 2.3 b | 34.4 ± 4.2 a | 17.7 ± 0.7 a | 16.6 ± 3.9 a |
| Melissa | ||||||
| control | 29.2 b | 29 a | 30.3 a | 27.2 c | 15.6 b | 23.7 a |
| Se+6 | 36 a | 33 a | 28.5 a | 36.4 ab | 17.4 b | 16.8 b |
| SeCys2 | 35 a | 27.1 a | 32 a | 29.5 b | 17.7 b | 17.1 b |
| Chlorella | 34.6 a | 30.7 a | 28.7 a | 32 bc | 15.6 b | 16.0 b |
| Se+6+ Ch | 39.2 a | 30.7 a | 32 a | 34 ab | 16.7 b | 17.6 b |
| SeCys2+ Ch | 39.3 a | 27.1 a | 35.8 a | 39.9 a | 16.5 b | 18.0 b |
| M ± SD | 35.6 ± 3.7 a | 29.6 ± 2.3 b | 31.2 ± 2.7 a | 33.2 ± 4.6 a | 16.6 ± 0.9 a | 18.2 ± 2.8 a |
| Cultivar | Leaves | Head | Roots | |||
|---|---|---|---|---|---|---|
| 70% Ethanol | Water | 70% Ethanol | Water | 70% Ethanol | Water | |
| Vertu 1340 | 33.6 ± 1.7 ab | 17.1 ± 0.7 | 30.7 ± 2.3 a | 21.9 ± 1.8 | 11.4 ± 1.0 a | 9.0 ± 0.6 |
| CV (%) | 5.1 | 4.1 | 7.5 | 8.2 | 8.8 | 6.7 |
| Golubtsy | 31.7 ± 0.8 b | 17.0 ± 0.5 | 24.0 ± 1.4 b | 21.3 ± 2.0 | 9.5 ± 0.7 ab | 8.7 ± 0.8 |
| CV (%) | 2.5 | 2.9 | 5.8 | 9.4 | 7.4 | 9.2 |
| Melissa | 37.3 ± 2.3 a | 17.3 ± 0.6 | 29.5 ± 1.7 a | 21.5 ± 1.8 | 9.2 ± 0. 6 b | 10.4 ± 0.8 |
| CV (%) | 6.2 | 5.8 | 6.5 | 7.7 | ||
| Treatment | Vertu 1340 | Golubtsy | Melissa | |||
|---|---|---|---|---|---|---|
| Leaves | Heads | Leaves | Heads | Leaves | Heads | |
| Dry matter (%) | 14.0 ± 0.46 b | 8.68 ± 0.56 c | 17.7 ± 0.66 a | 10.2 ± 1.02 c | 18.4 ± 0.22 a | 10.6 ± 0.62 c |
| CV (%) | 3.3 | 6.5 | 3.7 | 10.0 | 1.2 | 5.8 |
| Nitrates, mg g−1 d.w. | 3.6 ± 0.1 b | 3.8 ± 0.3 b | 3.8 ± 0.2 b | 5.1 ± 0.6 a | 3.7 ± 0.4 b | 4.6 ± 0.3 a |
| CV (%) | 3.39 ab | 8.66 | 7.24 | 13.5 | 11.3 | 14.1 |
| TDS, mg g−1 d.w. | 89.3 ± 5.6 a | 64.1 ± 2.0 b | 92.6 ± 2.5 a | 66.6 ± 2.4 b | 93.3 ± 2.2 a | 63.1 ± 1.7 b |
| CV (%) | 6.22 | 3.19 | 2.69 | 3.61 | 2.38 | 2.71 |
| Monosaccharides (% d.w.) | traces | 47.62 + 4.67 a | traces | 36.88 + 2.07 b | traces | 33.98 + 2.00 b |
| CV (%) | - | 9.81 | - | 5.61 | - | 5.90 |
| Total sugar (% d.w.) | Traces | 50.48 + 4.53 a | traces | 39.25 + 1.41 b | traces | 41.63 + 3.10 b |
| CV (%) | - | 8.97 | - | 3.59 | - | 7.45 |
| Month | Temperature (°C) | Precipitation (mm) | ||
|---|---|---|---|---|
| 2021 | 2022 | 2021 | 2022 | |
| May | 13.8 | 10.0 | 81 | 55.5 |
| June | 21.8 | 18.6 | 20 | 24.6 |
| July | 22.0 | 20.2 | 38 | 66.1 |
| August | 19.4 | 22.3 | 36 | 13.7 |
| September | 9.1 | 9.6 | 58 | 125.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoshkina, M.; Golubkina, N.; Poluboyarinov, P.; Skrypnik, L.; Sekara, A.; Tallarita, A.; Caruso, G. Effect of Sodium Selenate and Selenocystine on Savoy Cabbage Yield, Morphological and Biochemical Characteristics under Chlorella Supply. Plants 2023, 12, 1020. https://doi.org/10.3390/plants12051020
Antoshkina M, Golubkina N, Poluboyarinov P, Skrypnik L, Sekara A, Tallarita A, Caruso G. Effect of Sodium Selenate and Selenocystine on Savoy Cabbage Yield, Morphological and Biochemical Characteristics under Chlorella Supply. Plants. 2023; 12(5):1020. https://doi.org/10.3390/plants12051020
Chicago/Turabian StyleAntoshkina, Marina, Nadezhda Golubkina, Pavel Poluboyarinov, Liubov Skrypnik, Agnieszka Sekara, Alessio Tallarita, and Gianluca Caruso. 2023. "Effect of Sodium Selenate and Selenocystine on Savoy Cabbage Yield, Morphological and Biochemical Characteristics under Chlorella Supply" Plants 12, no. 5: 1020. https://doi.org/10.3390/plants12051020