
Genome-Wide Identification and Abiotic-Stress-Responsive Expression of CKX Gene Family in Liriodendron chinense
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Identification of CKX Gene Family in L. chinense Genome
2.2. Phylogenetic Analysis of LcCKX Proteins
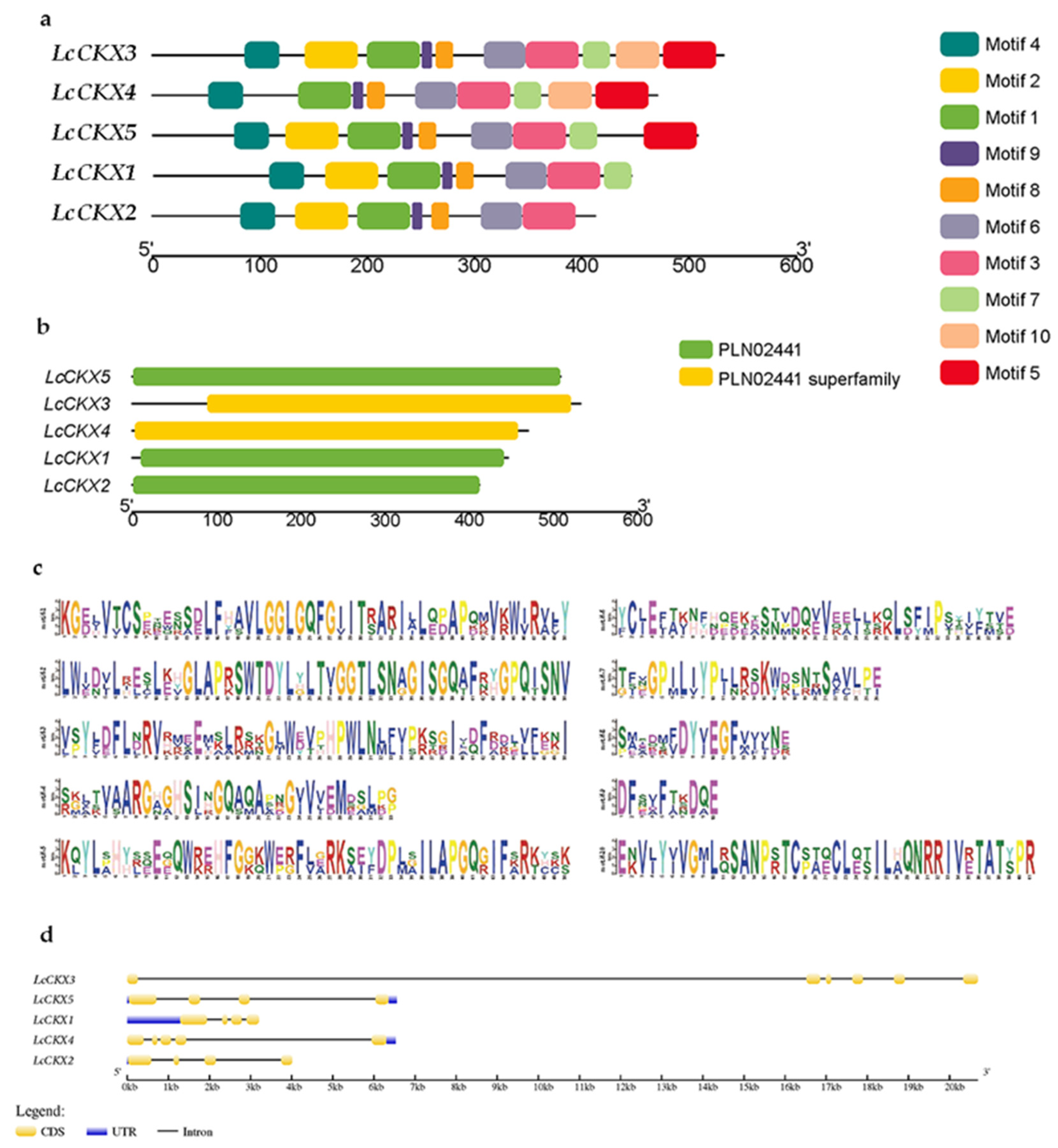
2.3. Analysis of Structure and Conserved Motifs of LcCKX Genes
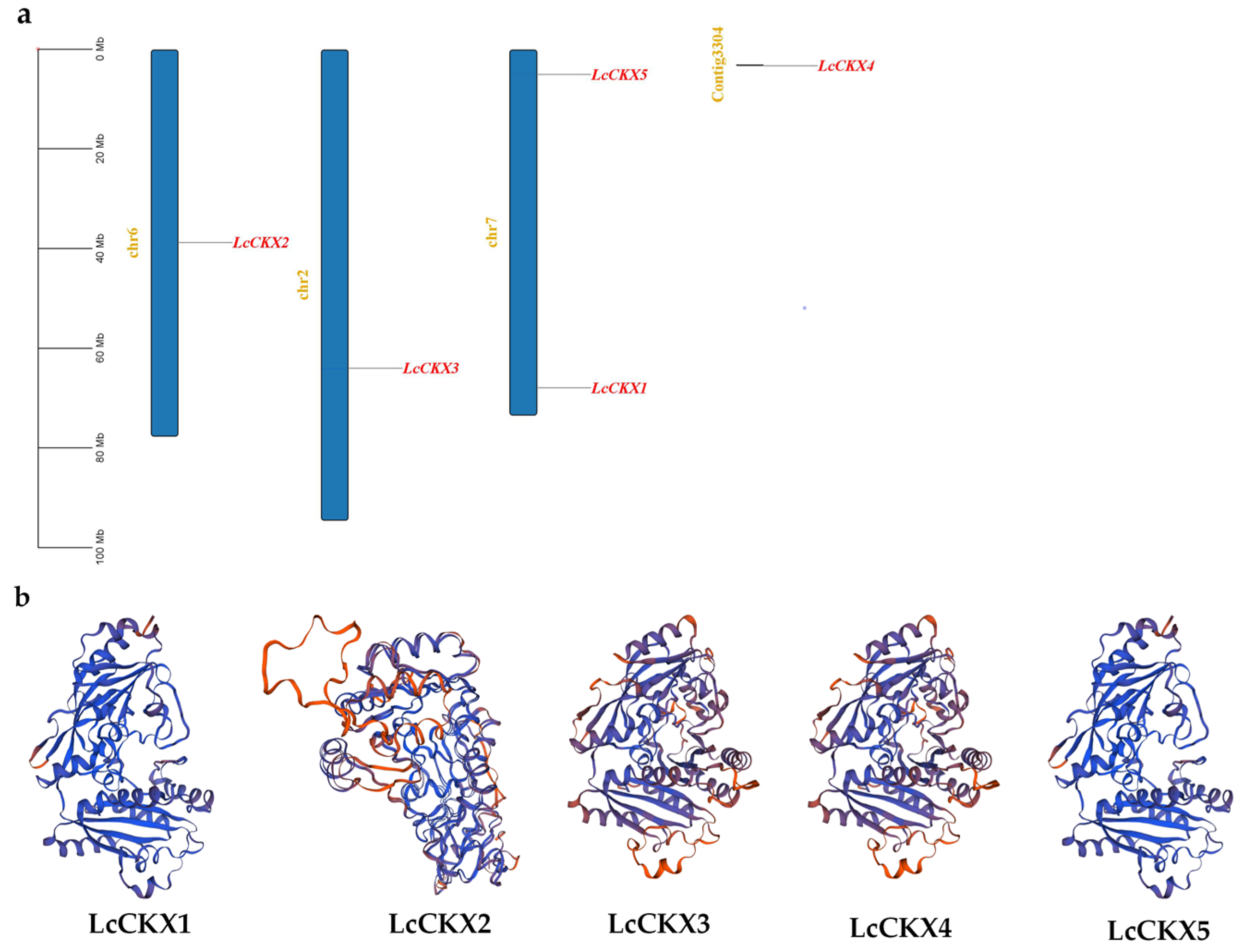
2.4. Chromosome Localization and Tertiary Structure of LcCKX Genes
2.5. Prediction of Cis-Acting Elements in LcCKX Promoter Regions
2.6. Gene Expression Pattern Analysis of LcCKX Family
2.7. Expression Analysis of LcCKX5 under Abiotic Stress
3. Discussion
4. Materials and Methods
4.1. Identification of CKX Genes in L. chinense
4.2. Protein Physicochemical Properties and Tertiary Structure Prediction of CKX Genes in L. chinense
4.3. Phylogenetic Analysis of CKX Genes in L. chinense
4.4. Gene Structure and Conserved Motif Analysis of CKX Genes in L. chinense
4.5. Cis-Acting Elements of CKX Genes in L. chinensis
4.6. RNA-seq Analysis of LcCKX Gene Expression Levels in Different Organs and under Multiple Stresses
4.7. Plant Materials and Abiotic Stress Treatment
4.8. RNA Extraction and Quantitative Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Lv, Y.; Zhang, M.; Liu, Y.; Kong, L.; Zou, M.; Lu, G.; Cao, J.; Yu, X. Identification, expression, and comparative genomic analysis of the IPT and CKX gene families in Chinese cabbage (Brassica rapa ssp. pekinensis). BMC Genom. 2013, 14, 594. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Bartrina, I.; Novak, O.; Strnad, M.; Werner, T.; Schmulling, T. The Cytokinin Status of the Epidermis Regulates Aspects of Vegetative and Reproductive Development in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 613488. [Google Scholar] [CrossRef] [PubMed]
- Geng, L.; Li, Q.; Jiao, L.; Xiang, Y.; Deng, Q.; Zhou, D.X.; Zhao, Y. WOX11 and CRL1 act synergistically to promote crown root development by maintaining cytokinin homeostasis in rice. New. Phytol. 2022, 237, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Schaller, G.E.; Street, I.H.; Kieber, J.J. Cytokinin and the cell cycle. Curr. Opin. Plant Biol. 2014, 21, 7–15. [Google Scholar] [CrossRef]
- Khuman, A.; Kumar, V.; Chaudhary, B. Evolutionary expansion and expression dynamics of cytokinin-catabolizing CKX gene family in the modern amphidiploid mustard (Brassica sp.). 3 Biotech. 2022, 12, 233. [Google Scholar] [CrossRef]
- Ramireddy, E.; Nelissen, H.; Leuendorf, J.E.; Van Lijsebettens, M.; Inzé, D.; Schmülling, T. Root engineering in maize by increasing cytokinin degradation causes enhanced root growth and leaf mineral enrichment. Plant Mol. Biol. 2021, 106, 555–567. [Google Scholar] [CrossRef]
- Gu, R.; Fu, J.; Guo, S.; Duan, F.; Wang, Z.; Mi, G.; Yuan, L. Comparative Expression and Phylogenetic Analysis of Maize Cytokinin Dehydrogenase/Oxidase (CKX) Gene Family. J. Plant Growth Regul. 2010, 29, 428–440. [Google Scholar] [CrossRef]
- Mahto, R.K.; Ambika; Singh, C.; Chandana, B.S.; Singh, R.K.; Verma, S.; Gahlaut, V.; Manohar, M.; Yadav, N.; Kumar, R. Chickpea Biofortification for Cytokinin Dehydrogenase via Genome Editing to Enhance Abiotic-Biotic Stress Tolerance and Food Security. Front. Genet. 2022, 13, 900324. [Google Scholar] [CrossRef]
- Noman, A.; Fahad, S.; Aqeel, M.; Ali, U.; Amanullah; Anwar, S.; Baloch, S.K.; Zainab, M. miRNAs: Major modulators for crop growth and development under abiotic stresses. Biotechnol. Lett. 2017, 39, 685–700. [Google Scholar] [CrossRef]
- Khan, A.; Tan, D.K.Y.; Afridi, M.Z.; Luo, H.; Tung, S.A.; Ajab, M.; Fahad, S. Nitrogen fertility and abiotic stresses management in cotton crop: A review. Environ. Sci. Pollut. R. 2017, 24, 14551–14566. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, Z.; Gu, Y.; Li, W.; Wang, W.; Yuan, X.; Zhang, Y.; Yuan, M.; Du, J.; Zhao, Q. Genome-wide identification of the soybean cytokinin oxidase/dehydrogenase gene family and its diverse roles in response to multiple abiotic stress. Front. Plant Sci. 2023, 14, 1163219. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, G.; Yi, M.; Gao, T.; Song, C.; Gao, Q.; Xia, Q.; Xiang, H. Whole-genome identification and salt- and ABA-induced expression trends of the Nicotiana tabacum CKX gene family. Plant Biotechnol. Rep. 2023. [Google Scholar] [CrossRef]
- Jain, P.; Singh, A.; Iquebal, M.A.; Jaiswal, S.; Kumar, S.; Kumar, D.; Rai, A. Genome-Wide Analysis and Evolutionary Perspective of the Cytokinin Dehydrogenase Gene Family in Wheat (Triticum aestivum L.). Front. Genet. 2022, 13, 931659. [Google Scholar] [CrossRef]
- Li, M.; Zhou, J.; Gong, L.; Zhang, R.; Wang, Y.; Wang, C.; Du, X.; Luo, Y.; Zhang, Y.; Wang, X.; et al. Identification and Expression Analysis of CKX Gene Family in Brassica juncea var. tumida and Their Functional Analysis in Stem Development. Horticulturae 2022, 8, 705. [Google Scholar] [CrossRef]
- Wu, W.; Zhu, S.; Zhu, L.; Wang, D.; Liu, Y.; Liu, S.; Zhang, J.; Hao, Z.; Lu, Y.; Cheng, T.; et al. Characterization of the Liriodendron Chinense MYB Gene Family and Its Role in Abiotic Stress Response. Front. Plant Sci. 2021, 12, 641280. [Google Scholar] [CrossRef]
- Wu, W.; Zhu, S.; Xu, L.; Zhu, L.; Wang, D.; Liu, Y.; Liu, S.; Hao, Z.; Lu, Y.; Yang, L.; et al. Genome-wide identification of the Liriodendron chinense WRKY gene family and its diverse roles in response to multiple abiotic stress. BMC Plant Biol. 2022, 22, 25. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, M.; Luo, Q.; Wang, J.; Li, H. De novo transcriptome analysis of Liriodendron chinense petals and leaves by Illumina sequencing. Gene 2014, 534, 155–162. [Google Scholar] [CrossRef]
- Zhong, Y.D.; Zhang, X.; Li, Y.Q.; Liu, L.P.; Yu, F.X. Genome Survey of Liriodendron chinense (Hemsl.) Sarg. Mol. Plant Breed. 2017, 15, 507–512. [Google Scholar] [CrossRef]
- Dash, P.K.; Rai, R. Green revolution to grain revolution: Florigen in the frontiers. J. Biotechnol. 2022, 343, 38–46. [Google Scholar] [CrossRef]
- Durán-Medina, Y.; Díaz-Ramírez, D.; Marsch-Martínez, N. Cytokinins on the Move. Front. Plant Sci. 2017, 8, 146. [Google Scholar] [CrossRef]
- Blume, R.; Yemets, A.; Korkhovyi, V.; Radchuk, V.; Rakhmetov, D.; Blume, Y. Genome-wide identification and analysis of the cytokinin oxidase/dehydrogenase (ckx) gene family in finger millet (Eleusine coracana). Front. Genet. 2022, 13, 963789. [Google Scholar] [CrossRef] [PubMed]
- Zhaodong, H. Liriodendron Genome Evolution and Thegenetic Basis of Flower Color Variation. Doctor Dissertation, Nanjing Forestry University, Nanjing, China, 2020. [Google Scholar]
- Nguyen, H.N.; Kambhampati, S.; Kisiala, A.; Seegobin, M.; Emery, R. The soybean (Glycine max L.) cytokinin oxidase/dehydrogenase multigene family; Identification of natural variations for altered cytokinin content and seed yield. Plant Direct 2021, 5, e308. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, H.; Zhu, H.; Ji, W.; Hou, Y.; Meng, Y.; Wen, J.; Mysore, K.S.; Li, X.; Lin, H. Genome-wide identification and characterization of cytokinin oxidase/dehydrogenase family genes in Medicago truncatula. J. Plant Physiol. 2021, 256, 153308. [Google Scholar] [CrossRef]
- Galuszka, P.; Frebortova, J.; Werner, T.; Yamada, M.; Strnad, M.; Schmulling, T.; Frebort, I. Cytokinin oxidase/dehydrogenase genes in barley and wheat: Cloning and heterologous expression. Eur. J. Biochem. 2004, 271, 3990–4002. [Google Scholar] [CrossRef] [PubMed]
- Kollmer, I.; Novak, O.; Strnad, M.; Schmulling, T.; Werner, T. Overexpression of the cytosolic cytokinin oxidase/dehydrogenase (CKX7) from Arabidopsis causes specific changes in root growth and xylem differentiation. Plant J. 2014, 78, 359–371. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, S.; Liang, Y.; Zhang, R.; Liu, L.; Qin, P.; Zhang, Z.; Wang, Y.; Zhou, J.; Tang, X.; et al. Loss-function mutants of OsCKX gene family based on CRISPR-Cas systems revealed their diversified roles in rice. Plant Genome 2023, e20283. [Google Scholar] [CrossRef]
- Yu, K.; Yu, Y.; Bian, L.; Ni, P.; Ji, X.; Guo, D.; Zhang, G.; Yang, Y. Genome-wide identification of cytokinin oxidases/dehydrogenase (CKXs) in grape and expression during berry set. Sci. Hortic-Amst. 2021, 280, 109917. [Google Scholar] [CrossRef]
- Zhang, C. Functional Studies on Cytokinin Oxidase/Dehydrogenase Gene in Mulberry. Masteral Dissertation, Southwest University, Chongqing, China, 2016. [Google Scholar]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef]
- Santner, A.; Estelle, M. Recent advances and emerging trends in plant hormone signalling. Nature 2009, 459, 1071–1078. [Google Scholar] [CrossRef]
- Zhengnan, L. Bioinformatics Analysis of Maize CKX Gene Family and Selection of Cas9 Transgenic Maize Lines. Masteral Dissertation, Yantai University, Yantai, China, 2021. [Google Scholar]
- Sharma, A.; Prakash, S.; Chattopadhyay, D. Killing two birds with a single stone—Genetic manipulation of cytokinin oxidase/dehydrogenase (CKX) genes for enhancing crop productivity and amelioration of drought stress response. Front. Genet. 2022, 13, 941595. [Google Scholar] [CrossRef]
- Kurepa, J.; Smalle, J.A. Auxin/Cytokinin Antagonistic Control of the Shoot/Root Growth Ratio and Its Relevance for Adaptation to Drought and Nutrient Deficiency Stresses. Int. J. Mol. Sci. 2022, 23, 1933. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zheng, H.; Zhang, X.; Sui, N. Cytokinins as central regulators during plant growth and stress response. Plant Cell Rep. 2021, 40, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Zu, S.H.; Jiang, Y.T.; Chang, J.H.; Zhang, Y.J.; Xue, H.W.; Lin, W.H. Interaction of brassinosteroid and cytokinin promotes ovule initiation and increases seed number per silique in Arabidopsis. J. Integr. Plant Biol. 2022, 64, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, M.; Meng, Z.; Wang, B.; Chen, M. Research Progress on the Roles of Cytokinin in Plant Response to Stress. Int. J. Mol. Sci. 2020, 21, 6574. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, Y.; Lu, S.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H.; Fang, Z.; Hou, X. Genome-wide identification and analysis of cytokinin dehydrogenase/oxidase (CKX) family genes in Brassica oleracea L. reveals their involvement in response to Plasmodiophora brassicae infections. Hortic. Plant J. 2022, 8, 68–80. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, L.; Fu, Q.; Xu, Z. Identification and expression analysis of cytokinin metabolic genes IPTs, CYP735A and CKXs in the biofuel plant Jatropha curcas. PeerJ 2018, 6, e4812. [Google Scholar] [CrossRef]
- Tan, M.; Li, G.; Qi, S.; Liu, X.; Chen, X.; Ma, J.; Zhang, D.; Han, M. Identification and expression analysis of the IPT and CKX gene families during axillary bud outgrowth in apple (Malus domestica Borkh.). Gene 2018, 651, 106–117. [Google Scholar] [CrossRef]
- Štefan, Č.; Alexandra, B.; Juraj, K. Relative quantification of mRNA: Comparison of methods currently used for real-time PCR data analysis. Bmc Mol. Biol. 2007, 8, 113. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Length (aa) | Molecular Weight (kDa) | Isoelectric Point (pl) | GRAVY |
|---|---|---|---|---|---|
| LcCKX1 | Lchi24464 | 446 | 49,620.79 | 8.8 | −0.136 |
| LcCKX2 | Lchi34027 | 412 | 45,216.77 | 6.1 | 0.044 |
| LcCKX3 | Lchi12502 | 532 | 59,699.25 | 6.64 | −0.196 |
| LcCKX4 | Lchi32870 | 470 | 53,131.66 | 5.98 | −0.164 |
| LcCKX5 | Lchi19652 | 508 | 56,314.36 | 5.7 | −0.089 |
| Gene Name | Plant Hormone | Environmental Stress | MYB Binding Site | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ABA | ZT | MeJA | GA | SA | Light | Defense | Circadian | Low Temperature | Drought Inducibility | |
| LcCKX1 | √ | √ | √ | √ | √ | √ | ||||
| LcCKX2 | √ | √ | √ | √ | √ | |||||
| LcCKX3 | √ | √ | √ | √ | √ | √ | √ | |||
| LcCKX4 | √ | √ | √ | √ | √ | √ | √ | |||
| LcCKX5 | √ | √ | √ | √ | √ | √ | ||||
| Gene Name | qRT-PCR Primer | |
|---|---|---|
| LcCKX5 | R | AAATGGCCTTCCTCTCGACG |
| F | CTTCGTTTCGGCCGTTCATC | |
| 18S | R | CTGCCTTCCTTGGATGTGGT |
| F | GCCCGTCGCTCTGATGAT | |
| Acting97 | R | TGGTCGCACAACTGGTATCG |
| F | TTCCCGTTCAGCAGTGGTCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Zhu, L.; Hao, Z.; Wu, W.; Xu, L.; Yang, Y.; Zhang, J.; Lu, Y.; Shi, J.; Chen, J. Genome-Wide Identification and Abiotic-Stress-Responsive Expression of CKX Gene Family in Liriodendron chinense. Plants 2023, 12, 2157. https://doi.org/10.3390/plants12112157
Sun X, Zhu L, Hao Z, Wu W, Xu L, Yang Y, Zhang J, Lu Y, Shi J, Chen J. Genome-Wide Identification and Abiotic-Stress-Responsive Expression of CKX Gene Family in Liriodendron chinense. Plants. 2023; 12(11):2157. https://doi.org/10.3390/plants12112157
Chicago/Turabian StyleSun, Xiao, Liming Zhu, Zhaodong Hao, Weihuang Wu, Lin Xu, Yun Yang, Jiaji Zhang, Ye Lu, Jisen Shi, and Jinhui Chen. 2023. "Genome-Wide Identification and Abiotic-Stress-Responsive Expression of CKX Gene Family in Liriodendron chinense" Plants 12, no. 11: 2157. https://doi.org/10.3390/plants12112157