
Pigment Biosynthesis and Molecular Genetics of Fruit Color in Pepper


Abstract
:1. Introduction
2. Biosynthetic Pathways of Pigments in Pepper Fruits
2.1. Classification of Pigments in Pepper Fruits
2.2. Pathway of Pigment Biosynthesis in Pepper Fruits
2.2.1. Biosynthetic Pathway of Chlorophyll
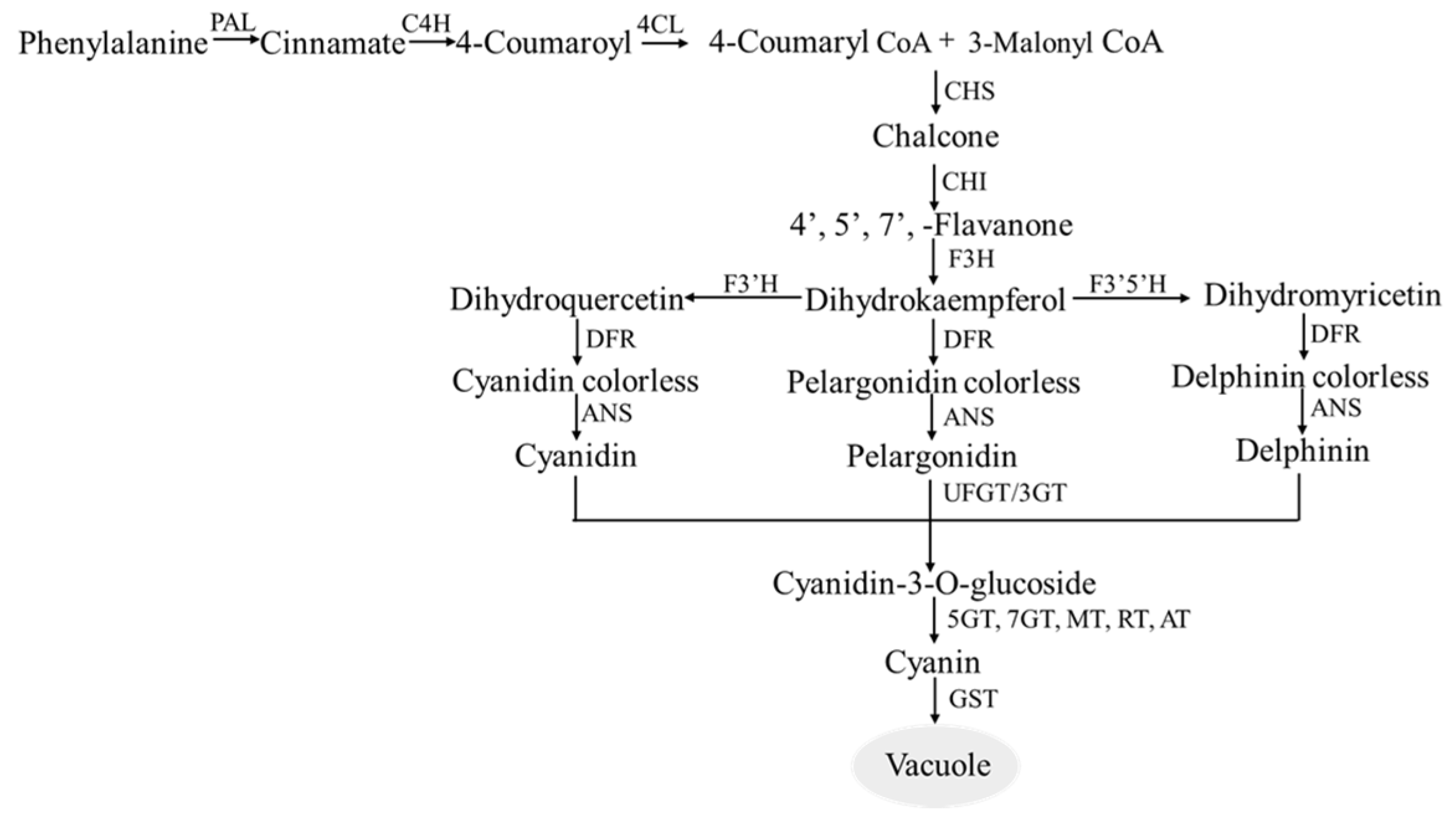
2.2.2. Biosynthetic Pathway of Anthocyanin
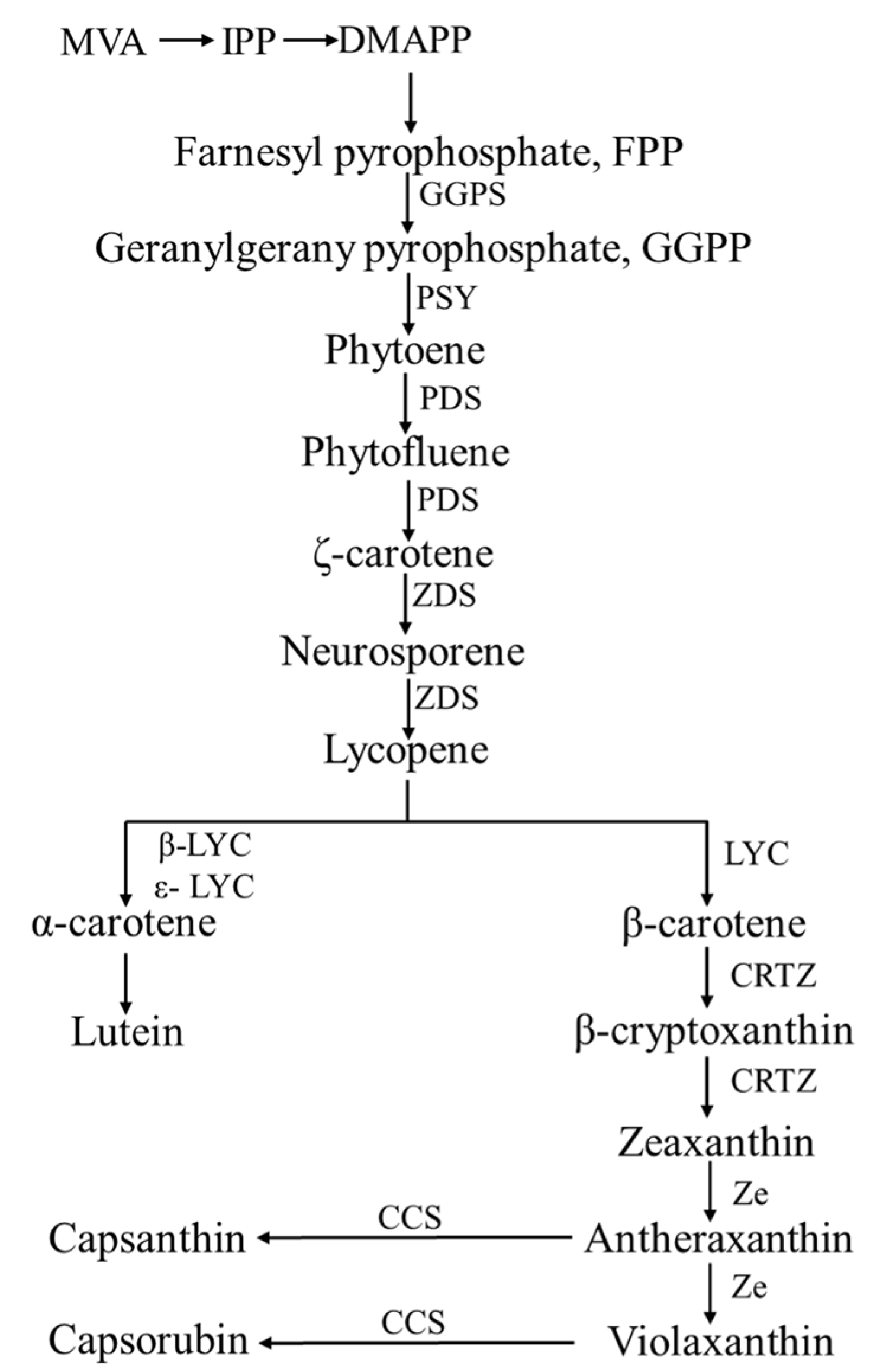
2.2.3. Biosynthetic Pathway of Carotenoid
2.3. Relationship between Fruit Color and Pigment Formation of Pepper
3. Research of Immature Fruit Color in Pepper
3.1. Research of Green and White Immature Fruit Related Genes in Pepper
3.2. Research of Violet and Nearly Black Immature Fruit Related Genes in Pepper
4. Research of the Mature Fruit Color in Pepper
4.1. Research of Red, Yellow, and Orange Mature Fruit Related Genes in Pepper
4.2. Research of White, Green, and Brown Mature Fruit Related Genes in Pepper
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parisi, M.; Alioto, D.; Tripodi, P. Overview of biotic stresses in pepper (Capsicum spp.): Sources of genetic resistance, molecular breeding and genomics. Int. J. Mol. Sci. 2020, 21, 2587. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.J.; Peng, J.; Li, Y.J. Recent advances in the study on capsaicinoids and capsinoids. Eur. J. Pharmacol. 2011, 650, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.D.; Silva, B.P.; Lopes, Â.C.; Carvalho, L.C.; Gomes, R.L. Selection of pepper accessions with ornamental potential. Rev. Caatinga 2019, 32, 566–574. [Google Scholar] [CrossRef]
- Sharoni, Y.; Linnewiel-Hermoni, K.; Khanin, M.; Salman, H.; Veprik, A.; Danilenko, M.; Levy, J. Carotenoids and apocarotenoids in cellular signaling related to cancer: A review. Mol. Nutr. Food Res. 2012, 56, 259–269. [Google Scholar] [CrossRef]
- Sporn, M.B.; Liby, K.T. Is lycopene an effective agent for preventing prostate cancer? Cancer Prev. Res 2013, 6, 384–386. [Google Scholar] [CrossRef]
- Walter, M.H.; Floss, D.S.; Strack, D. Apocarotenoids: Hormones, mycorrhizal metabolites and aroma volatiles. Planta 2010, 232, 1–17. [Google Scholar] [CrossRef]
- Jeong, H.B.; Jang, S.J.; Kang, M.Y.; Kim, S.; Kwon, J.K.; Kang, B.C. Candidate gene analysis reveals that the fruit color locus C1 corresponds to PRR2 in pepper (Capsicum frutescens). Front. Plant Sci. 2020, 11, 399. [Google Scholar] [CrossRef]
- Egea, I.; Barsan, C.; Bian, W.; Purgatto, E.; Latché, A.; Chervin, C.; Bouzayen, M.; Pech, J.C. Chromoplast differentiation: Current status and perspectives. Plant Cell Physiol. 2010, 51, 1601–1611. [Google Scholar] [CrossRef]
- Kahlau, S.; Bock, R. Plastid transcriptomics and translatomics of tomato fruit development and chloroplast-to-chromoplast differentiation: Chromoplast gene expression largely serves the production of a single protein. Plant Cell 2008, 20, 856–874. [Google Scholar] [CrossRef]
- Barsan, C.; Zouine, M.; Maza, E.; Bian, W.; Egea, I.; Rossignol, M.; Pech, J.C. Proteomic analysis of chloroplast-to-chromoplast transition in tomato reveals metabolic shifts coupled with disrupted thylakoid biogenesis machinery and elevated energy-production components. Plant Physiol. 2012, 160, 708–725. [Google Scholar] [CrossRef]
- Suzuki, M.; Takahashi, S.; Kondo, T.; Dohra, H.; Ito, Y.; Kiriiwa, Y.; Hayashi, M.; Kamiya, S.; Kato, M.; Fujiwara, M.; et al. Plastid proteomic analysis in tomato fruit development. PLoS ONE 2015, 10, e0137266. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Liang, B.; Chang, B.; Yan, J.; Liu, L.; Wang, Y.; Yang, Y.; Zhao, Z. Transcriptome profiling of anthocyanin biosynthesis in the peel of ‘Granny Smith’apples (Malus domestica) after bag removal. BMC Genom. 2019, 20, 353. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.; Zachariae, A.; Lenz, F.; Eckhardt, G. Carotenoid changes in the peel of the «Golden delicious» apple during ripening and storage. Zeitschrift für Pflanzenphysiologie 1978, 89, 321–332. [Google Scholar] [CrossRef]
- Ramirez, D.A.; Tomes, M.L. Relationship between chlorophyll and carotenoid biosynthesis in dirty-red (green-flesh) mutant in tomato. Bot. Gaz. 1964, 125, 221–226. [Google Scholar] [CrossRef]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.P.; Matros, A.; Peterek, S.; Schijlen, E.G.W.M.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Z.; Guan, L.; Zheng, T.; Jiu, S.; Zhu, X.; Jia, H.; Fang, J. Changes of anthocyanin component biosynthesis in‘Summer Black’ grape berries after the red flesh mutation occurred. J. Agri. Food Chem. 2018, 66, 9209–9218. [Google Scholar] [CrossRef]
- Niu, S.S.; Xu, C.J.; Zhang, W.S.; Zhang, B.; Li, X.; Wang, L.K.; Ferguson, I.B.; Allan, A.C.; Chen, K.S. Coordinated regulation of anthocyanin biosynthesis in Chinese bayberry (Myrica rubra) fruit by a R2R3 MYB transcription factor. Planta 2010, 231, 887–899. [Google Scholar] [CrossRef]
- Fanciullino, A.L.; Dhuique-Mayer, C.; Froelicher, Y.; Talón, M.; Ollitrault, P.; Morillon, R. Changes in carotenoid content and biosynthetic gene expression in juice sacs of four orange varieties (Citrus sinensis) differing in flesh fruit color. J. Agri. Food Chemis. 2008, 56, 3628–3638. [Google Scholar] [CrossRef]
- Borovsky, Y.; Oren-Shamir, M.; Ovadia, R.; De Jong, W.; Paran, I. The A locus that controls anthocyanin accumulation in pepper encodes a MYB transcription factor homologous to Anthocyanin2 of Petunia. Theor. Appl. Genet. 2004, 109, 23–29. [Google Scholar] [CrossRef]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Qiao, Y.M.; Li, Y.; Yu, Y.N.; Gong, Z.H. Identification of Fruit Traits Related QTLs and a Candidate Gene, CaBRX, Controlling Locule Number in Pepper (Capsicum annuum L.). Horticulturae 2022, 8, 146. [Google Scholar] [CrossRef]
- Rout, S.S.; Rout, P.; Uzair, M.; Kumar, G.; Nanda, S. Genome-wide identification and expression analysis of CRK gene family in chili pepper (Capsicum annuum L.) in response to Colletotrichum truncatum infection. J. Hortic. Sci. Biotechnol. 2023, 98, 194–206. [Google Scholar] [CrossRef]
- Wu, Y.; Popovsky-Sarid, S.; Tikunov, Y.; Borovsky, Y.; Baruch, K.; Visser, R.G.; Paran, I.; Bovy, A.G. CaMYB12-like underlies a major QTL for flavonoid content in pepper (Capsicum annuum) fruit. New Phytol. 2023, 237, 2255–2267. [Google Scholar] [CrossRef] [PubMed]
- Chae, S.Y.; Lee, K.; Do, J.W.; Hong, S.C.; Lee, K.; Cho, M.C.; Yang, E.Y.; Yoon, J.B. QTL Mapping of Resistance to Bacterial Wilt in Pepper Plants (Capsicum annuum) Using Genotyping-by-Sequencing (GBS). Horticulturae 2022, 8, 115. [Google Scholar] [CrossRef]
- Shu, H.; He, C.; Mumtaz, M.A.; Hao, Y.; Zhou, Y.; Jin, W.; Zhu, J.; Bao, W.; Cheng, S.; Zhu, G.; et al. Fine mapping and identification of candidate genes for fruit color in pepper (Capsicum chinense). Sci. Hortic. 2023, 310, 111724. [Google Scholar] [CrossRef]
- Fraser, P.D.; Bramley, P.M. The biosynthesis and nutritional uses of carotenoids. Prog. Lipid Res. 2004, 43, 228–265. [Google Scholar] [CrossRef]
- Bley, K.; Boorman, G.; Mohammad, B.; McKenzie, D.; Babbar, S. A comprehensive review of the carcinogenic and anticarcinogenic potential of capsaicin. Toxicol. Pathol. 2012, 40, 847–873. [Google Scholar] [CrossRef]
- Gómez-García, M.D.R.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in Chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025–19053. [Google Scholar] [CrossRef]
- Shifriss, C.; Pilovsky, M. Studies of the inheritance of mature fruit color in Capsicum annuum L. Euphytica 1992, 60, 123–126. [Google Scholar] [CrossRef]
- Lightbourn, G.J.; Griesbach, R.J.; Novotny, J.A.; Clevidence, B.A.; Rao, D.; Stommel, J.R. Effects of anthocyanin and carotenoid combinations on foliage and immature fruit color of Capsicum annuum L. J. Hered. 2008, 99, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.L.; Li, L.; Chai, W.; Shah, S.M.; Gong, Z. Effects of silencing key genes in the capsanthin biosynthetic pathway on fruit color of detached pepper fruits. BMC Plant Biol. 2014, 14, 314. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, J.E.; Lister, C.E.; Reay, P.F.; Triggs, C.M. Influence of pigment composition on skin color in a wide range of fruit and vegetables. J. Am. Soc. Hortic. Sci. 1997, 122, 594–598. [Google Scholar] [CrossRef]
- Omori, S.; Sasanuma, T. Identification of large deletion of Ccs responsible for non-red fruit color in pepper (Capsicum annuum) and development of DNA marker to distinguish the deletion. Am. J. Plant Sci. 2022, 13, 1233–1246. [Google Scholar] [CrossRef]
- Banerjee, S.; Bhattacharjee, T.; Maurya, P.K.; Mukherjee, D.; Islam, S.M.; Chattopadhyay, A.; Hazra, P. Genetic control of qualitative and quantitative traits in bell pepper crosses involving varied fruit colors and shapes. Int. J. Veg. Sci. 2022, 28, 477–492. [Google Scholar] [CrossRef]
- Matsufuji, H.; Ishikawa, K.; Nunomura, O.; Chino, M.; Takeda, M. Anti-oxidant content of different coloured sweet peppers, white, green, yellow, orange and red (Capsicum annuum L.). Int. J. Food Sci. Technol. 2007, 42, 1482–1488. [Google Scholar] [CrossRef]
- Tian, S.L.; Li, L.L.; Shah, S.N.; Gong, Z. The relationship between red fruit colour formation and key genes of capsanthin biosynthesis pathway in Capsicum annuum. Biol. Plantarum 2015, 59, 507–513. [Google Scholar] [CrossRef]
- Smith, P.G. Inheritance of brown and green mature fruit color in peppers. J. Hered. 1950, 41, 138–140. [Google Scholar] [CrossRef]
- Roca, M.; Hornero-Méndez, D.; Gandul-Rojas, B.; Mínguez-Mosquera, M.I. Stay-green phenotype slows the carotenogenic process in Capsicum annuum (L.) fruits. J. Agricul. Food Chem. 2006, 54, 8782–8787. [Google Scholar] [CrossRef]
- Eckhardt, U.; Grimm, B.; Hörtensteiner, S. Recent advances in chlorophyll biosynthesis and breakdown in higher plants. Plant Mol. Biol. 2004, 56, 1–14. [Google Scholar] [CrossRef]
- Bollivar, D.W. Recent advances in chlorophyll biosynthesis. Photosyn. Res. 2006, 90, 173–194. [Google Scholar] [CrossRef]
- Zhang, S.; Heyes, D.J.; Feng, L.; Sun, W.; Johannissen, L.O.; Liu, H.; Levy, C.W.; Li, X.; Yang, J.E.; Yu, X.; et al. Structural basis for enzymatic photocatalysis in chlorophyll biosynthesis. Nature 2019, 574, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Cheng, S.C.; Cai, J.H.; Wei, B.D.; Zhou, X.; Zhou, Q.; Ji, S.J. Chlorophyll degradation and carotenoid biosynthetic pathways: Gene expression and pigment content in broccoli during yellowing. Food Chem. 2019, 297, 124964. [Google Scholar] [CrossRef]
- Wang, W.Q.; Tang, W.J.; Ma, T.T.; Niu, D.; Jin, J.B.; Wang, H.; Lin, R. A pair of light signaling factors FHY3 and FAR1 regulates plant immunity by modulating chlorophyll biosynthesis. J. Integ. Plant Biol. 2016, 58, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Beale, S.I. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.J.; Willows, R.D. Mechanism and regulation of Mg-chelatase. Biochem. J. 1997, 327, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Gu, T.; Khan, I.; Zada, A.; Jia, T. Research progress in the interconversion, turnover and degradation of chlorophyll. Cells 2021, 10, 3134. [Google Scholar] [CrossRef]
- Tanaka, A.; Ito, H.; Tanaka, R.; Tanaka, N.K.; Yoshida, K.I.; Okada, K. Chlorophyll a oxygenase (CAO) is involved in chlorophyll b formation from chlorophyll a. Proc. Natl. Acad. Sci. USA 1998, 95, 12719–12723. [Google Scholar] [CrossRef]
- Nagata, N.; Tanaka, R.; Satoh, S.; Tanaka, A. Identification of a vinyl reductase gene for chlorophyll synthesis in Arabidopsis thaliana and implications for the evolution of Prochlorococcus species. Plant Cell 2005, 17, 233–240. [Google Scholar] [CrossRef]
- Kobayashi, K.; Masuda, T. Transcriptional regulation of tetrapyrrole biosynthesis in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1811. [Google Scholar] [CrossRef]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef] [PubMed]
- Seren, S.; Lieberman, R.; Bayraktar, U.D.; Heath, E.; Sahin, K.; Andic, F.; Kucuk, O. Lycopene in cancer prevention and treatment. Am. J. Therapeut. 2008, 15, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, B.; Du, H.; Zhang, F.; Zhang, H.; Wang, Y.; He, H.; Geng, S.; Zhang, X. Genetic mapping of anthocyanin accumulation-related genes in pepper fruits using a combination of SLAF-seq and BSA. PLoS ONE 2018, 13, e0204690. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Al Sane, K.O.; Hesham, A.E.L. Biochemical and genetic evidences of anthocyanin biosynthesis and accumulation in a selected tomato mutant. Rend. Lincei 2015, 26, 293–306. [Google Scholar] [CrossRef]
- Zhang, Y.; Chu, G.; Hu, Z.; Gao, Q.; Cui, B.; Tian, S.; Wang, B.; Chen, G. Genetically engineered anthocyanin pathway for high health-promoting pigment production in eggplant. Mol. Breed. 2016, 36, 54. [Google Scholar] [CrossRef]
- LaFountain, A.M.; Yuan, Y.W. Repressors of anthocyanin biosynthesis. New Phytol. 2021, 231, 933–949. [Google Scholar] [CrossRef]
- He, Y.; Zhang, X.; Li, L.; Sun, Z.; Li, J.; Chen, X.; Hong, G. SPX4 interacts with both PHR1 and PAP1 to regulate critical steps in phosphorus-status-dependent anthocyanin biosynthesis. New Phytol. 2021, 230, 205–217. [Google Scholar] [CrossRef]
- Deli, J.; Matus, Z.; Tóth, G. Carotenoid composition in the Fruits of Capsicum annuum cv. Szentesi Kosszarvú during Ripening. J. Agricult. Food Chem. 1996, 44, 711–716. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef]
- Borovsky, Y.; Paran, I. Chlorophyll breakdown during pepper fruit ripening in the chlorophyll retainer mutation is impaired at the homolog of the senescence-inducible stay-green gene. Theor. Appl. Genet. 2008, 117, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, G. Diversity and origin of carotenoid biosynthesis: Its history of co-evolution towards plant photosynthesis. New Phytol. 2021, 232, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Chettry, U.; Chrungoo, N.K. A multifocal approach towards understanding the complexities of carotenoid biosynthesis and accumulation in rice grains. Brief. Funct. Genom. 2020, 19, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.B.; Kang, M.; Jung, A.; Han, K.; Lee, J.; Jo, J.; Lee, H.; An, J.; Kim, S.; Kang, B. Single-molecule real-time sequencing reveals diverse allelic variations in carotenoid biosynthetic genes in pepper (Capsicum spp.). Plant Biotechnol. J. 2019, 17, 1081–1093. [Google Scholar] [CrossRef]
- Luo, H.; Li, Z.; Straight, C.R.; Wang, Q.; Zhou, J.; Sun, Y.; Lo, C.; Yi, L.; Wu, Y.; Huang, J.; et al. Black pepper and vegetable oil-based emulsion synergistically enhance carotenoid bioavailability of raw vegetables in humans. Food Chem. 2022, 373, 131277. [Google Scholar] [CrossRef] [PubMed]
- Kuai, B.; Chen, J.; Hörtensteiner, S. The biochemistry and molecular biology of chlorophyll breakdown. J. Exp. Bot. 2018, 69, 751–767. [Google Scholar] [CrossRef]
- Li, S.; Chen, K.; Grierson, D. Molecular and hormonal mechanisms regulating fleshy fruit ripening. Cells 2021, 10, 1136. [Google Scholar] [CrossRef]
- Matus, Z.; Deli, J.; Szabolcs, J. Carotenoid composition of yellow pepper during ripening: Isolation of. beta.-cryptoxanthin 5, 6-epoxide. J. Agricul. Food Chem. 1991, 39, 1907–1914. [Google Scholar] [CrossRef]
- Lang, Y.Q.; Yanagawa, S.; Sasanuma, T.; Sasakuma, T. Orange fruit color in Capsicum due to deletion of capsanthin-capsorubin synthesis gene. Breed. Sci. 2004, 54, 33–39. [Google Scholar] [CrossRef]
- Guzman, I.; Hamby, S.; Romero, J.; Bosland, P.W.; O’Connell, M.A. Variability of carotenoid biosynthesis in orange colored Capsicum spp. Plant Sci. 2010, 179, 49–59. [Google Scholar] [CrossRef]
- Brand, A.; Borovsky, Y.; Hill, T.A.; Rahman, K.A.; Bellalou, A.; Deynze, A.V.; Paran, I. CaGLK2 regulates natural variation of chlorophyll content and fruit color in pepper fruit. Theor. Appl. Genet. 2014, 127, 2139–2148. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Liu, T.G.; Jiang, H.K.; Ding, P.P.; Yu, Q.F.; Tian, H.M.; Jia, L. Genetic analysis on white color of pepper fruit. China Veg. 2017, 37–42. [Google Scholar]
- Brand, A.; Borovsky, Y.; Meir, S.; Rogachev, I.; Aharoni, A.; Paran, I. pc8.1, a major QTL for pigment content in pepper fruit, is associated with variation in plastid compartment size. Planta 2011, 235, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Borovsky, Y.; Monsonego, N.; Mohan, V.; Shabtai, S.; Kamara, I.; Faigenboim, A.; Paran, I. The zinc-finger transcription factor CcLOL1 controls chloroplast development and immature pepper fruit color in Capsicum chinense and its function is conserved in tomato. Plant J. 2019, 99, 41–55. [Google Scholar] [CrossRef]
- Safi, A.; Medici, A.; Szponarski, W.; Ruffel, S.; Lacombe, B.; Krouk, G. The world according to GARP transcription factors. Curr. Opin. Plant Biol. 2017, 39, 159–167. [Google Scholar] [CrossRef]
- Pan, Y.; Bradley, G.; Pyke, K.; Ball, G.R.; Lu, C.; Fray, R.G.; Marshall, A.; Jayasuta, S.; Baxter, C.J.; van Wijk, R.; et al. Network inference analysis identifies an APRR2-like gene linked to pigment accumulation in tomato and pepper fruits. Plant Physiol. 2013, 161, 1476–1485. [Google Scholar] [CrossRef]
- Xue, L.B.; Wu, X.X.; Chen, J.L. Study on heredity of fruit colors of sweet pepper. Acta Hortic. Sin. 2005, 32, 513–515, (In Chinese with English Abstract). [Google Scholar]
- Chaim, A.B.; Borovsky, Y.; De Jong, W.S.; Paran, I. Linkage of the A locus for the presence of anthocyanin and fs10.1, a major fruit-shape QTL in pepper. Theor. Appl. Genet. 2003, 106, 889–894. [Google Scholar] [CrossRef]
- Aza-Gonzalez, C.; Herrera-Isidrón, L.; Núñez-Palenius, H.G.; Martínez De La Vega, O.; Ochoa-Alejo, N. Anthocyanin accumulation and expression analysis of biosynthesis-related genes during chili pepper fruit development. Biol. Plant. 2013, 57, 49–55. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Tran, G.; Nguyen, N.H. The MYB-bHLH-WDR interferers (MBWi) epigenetically suppress the MBW’s targets. Biol. Cell 2019, 111, 284–291. [Google Scholar] [CrossRef]
- Peterson, P.A. Linkage of fruit shape and color genes in capsicum. Genetics 1959, 44, 407–419. [Google Scholar] [CrossRef]
- Lefebvre, V.; Caranta, C.; Pflieger, S.; Moury, B.; Daubeze, A.M.; Blattes, A.; Palloix, A. Updated intraspecific maps of pepper. Capsicum Eggplant Newsl. 1997, 16, 36–41. [Google Scholar]
- Zhang, R.X.; Cheng, G.X.; Liu, G.T.; Chen, S.Y.; ul Haq, S.; Khan, A.; Li, Q.; Gong, Z.H. Assessing the functional role of color-related CaMYB gene under cold stress using virus-induced gene silencing in the fruit of pepper (Capsicum annuum L.). Sci. Horticul. 2020, 272, 109504. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Jin, J.; Yin, Y.; Zhang, H.; Chai, W.; Gong, Z. VIGS approach reveals the modulation of anthocyanin biosynthetic genes by CaMYB in chili pepper leaves. Front. Plant Sci. 2015, 6, 500. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Barragán, A.; Ochoa-Alejo, N. Virus-induced silencing of MYB and WD40 transcription factor genes affects the accumulation of anthocyanins in chilli pepper fruit. Biol. Plant. 2014, 58, 567–574. [Google Scholar] [CrossRef]
- Guo, Y.M.; Bai, J.J.; Duan, X.D.; Wang, J. Accumulation characteristics of carotenoids and adaptive fruit color variation in ornamental pepper. Sci. Hortic. 2021, 275, 109699. [Google Scholar] [CrossRef]
- Taller, J.; Hirata, Y.; Yagishita, N.; Kita, M.; Ogata, S. Graft-induced genetic changes and the inheritance of several characteristics in pepper (Capsicum annuum L.). Theor. Appl. Genet. 1998, 97, 705–713. [Google Scholar] [CrossRef]
- Hurtado-Hernandez, H.; Smith, P.G. Inheritance of mature fruit color in Capsicum annuum L. J. Hered. 1985, 76, 211–213. [Google Scholar] [CrossRef]
- Thorup, T.; Tanyolaç, B.; Livingstone, K.; Popovsky, S.N.; Paran, I.; Jahn, M.M. Candidate gene analysis of organ pigmentation loci in the Solanaceae. Proc. Natl. Acad. Sci. USA 2000, 97, 11192–11197. [Google Scholar] [CrossRef]
- Houlné, G.; Schantz, M.L.; Meyer, B.; Pozueta-Romero, J.; Schantz, R. A chromoplast-specific protein in Capsicum annuum: Characterization and expression of the corresponding gene. Curr. Genet. 1994, 26, 524–527. [Google Scholar] [CrossRef]
- Deruère, J.; Bouvier, F.; Steppuhn, J.; Klein, A.; Camara, B.; Kuntz, M. Structure and expression of two plant genes encoding chromoplasttranscripts. Biochem. Bioph. Res. Co. 1994, 199, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, F.; Hugueney, P.; d’Harlingue, A.; Kuntz, M.; Camara, B. Xanthophyll biosynthesis in chromoplasts: Isolation and molecular cloning of an enzyme catalyzing the conversion of 5, 6-epoxycarotenoid into ketocarotenoid. Plant J. 1994, 6, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, V.; Kuntz, M.; Camara, B.; Palloix, A. The capsanthin-capsorubin synthase gene: A candidate gene for the y locus controlling the red fruit colour in pepper. Plant Mol. Biol. 1998, 36, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Popovsky, S.N.; Paran, I. Molecular genetics of the y locus in pepper: Its relation to capsanthin-capsorubin synthase and to fruit color. Theor. Appl. Genet. 2000, 101, 86–89. [Google Scholar] [CrossRef]
- Ha, S.H.; Kim, J.B.; Park, J.S.; Lee, S.; Cho, K. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours: Deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper. J. Exp. Bot. 2007, 58, 3135–3144. [Google Scholar] [CrossRef]
- Li, Z.; Wang, S.; Gui, X.L.; Chang, X.B.; Gong, Z.H. A further analysis of the relationship between yellow ripe-fruit color and the capsanthin-capsorubin synthase gene in pepper (Capsicum sp.) indicated a new mutant variant in C. annuum and a tandem repeat structure in promoter region. PLoS ONE 2013, 8, e61996. [Google Scholar] [CrossRef]
- Tian, S.L.; Li, Z.; Li, L.; Shah, S.N.; Gong, Z. Analysis of tandem repeat units of the promoter of capsanthin/capsorubin synthase (Ccs) gene in pepper fruit. Physiol. Mol. Biol. Plants 2017, 23, 685–691. [Google Scholar] [CrossRef]
- Kim, O.R.; Cho, M.C.; Kim, B.; Huh, J.H. A splicing mutation in the gene encoding phytoene synthase causes orange coloration in Habanero pepper fruits. Mol. Cells 2010, 30, 569–574. [Google Scholar] [CrossRef]
- Borovsky, Y.; Tadmor, Y.; Bar, E.; Meir, A.; Lewinsohn, E.; Paran, I. Induced mutation in β-CAROTENE HYDROXYLASE results in accumulation of β-carotene and conversion of red to orange color in pepper fruit. Theor. Appl. Genet. 2013, 126, 557–565. [Google Scholar] [CrossRef]
- Lee, S.B.; Kim, J.E.; Kim, H.; Lee, G.; Kim, B.; Lee, J.M. Genetic mapping of the c1 locus by GBS-based BSA-seq revealed Pseudo-Response Regulator 2 as a candidate gene controlling pepper fruit color. Theor. Appl. Genet. 2020, 133, 1897–1910. [Google Scholar] [CrossRef]
- Barry, C.S.; McQuinn, R.P.; Chung, M.Y.; Besuden, A.; Giovannoni, J.J. Amino acid substitutions in homologs of the STAY-GREEN protein are responsible for the green-fresh and chlorophyll retainer mutations of tomato and pepper. Plant Physiol. 2008, 147, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Tian, S.L.; Shah, S.M.; Pan, B.; Diao, W.; Gong, Z. Cloning and characterization of the CarbcL gene related to chlorophyll in pepper (Capsicum annuum L.) under fruit shade stress. Front. Plant Sci. 2015, 6, 850. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Short Name | Enzyme Name | Chromosome Location | Gene ID |
|---|---|---|---|
| GluTR | Glutamyl-tRNA reductase | 4 | LOC107867325 |
| 8 | LOC107840125 | ||
| GSA-AM | Glutamate-1-semialdehyde 2,1-aminomutase | 5 | LOC107863035 |
| PBGS (ALAD) | Porphobilinogen synthase (5-Aminolevulinate dehydratase) | 1 | LOC107856090 |
| PBGD | Porphobilinogen deaminase (Hydroxymethylbilane synthase) | 7 | LOC107878895 |
| UROS | Uroporphyrinogen III synthase (Uroporphyrinogen III co-synthase) | 4 | LOC107867127 |
| UROD | Uroporphyrinogen III decarboxylase | 6 | LOC107875768 |
| 10 | LOC107845519 | ||
| CPOX | Coproporphyrinogen III oxidase | 10 | LOC107845388 |
| PPOX | Protoporphyrinogen oxidase | 1 | LOC107840071 |
| MgCh | Magnesium chelatase H subunit | 5 | LOC107870073 |
| Magnesium chelatase I subunit | 10 | LOC107845383 | |
| Magnesium chelatase D subunit | 5 | LOC107870054 | |
| MgPMT | Magnesium proto IX methyltransferase | 3 | LOC107862118 |
| MgPEC | Mg-protoporphyrin monomethylester | 10 | LOC107844048 |
| DVR | 3,8-Divinyl protochlorophyllide a 8-vinyl reductase | 1 | LOC107842746 |
| POR | Protochlorophyllide oxidoreductase | 3 | LOC107863251 |
| CHLG | Chlorophyll synthase | 9 | LOC107842693 |
| CAO | Chlorophyllide a oxygenase | 6 | LOC107872996 |
| Short Name | Enzyme Name | Gene Name | Chromosome Location | Gene ID |
|---|---|---|---|---|
| PAL | Phenylalanine ammonia-lyase | PAL1 | 9 | LOC107843092 |
| C4H | Cinnamate 4-hydroxy-lase | C4H | 6 | LOC107875406 |
| 4CL | 4-counmaric CoA ligase | 4CL | 3 | LOC107862076 |
| CHS | Chalcone synthase | CHS | 5 | LOC107871256 |
| CHI | Chalcone isomeras | CHI | 11 | LOC107852750 |
| F3H | Flavanone-3-hydroxylase | F3H | 2 | LOC107859880 |
| DFR | Dihydroflavonol-4-reductase | 2 | LOC107860031 | |
| ANS | Anthocyanidin synthase | OXR1 | 10 | LOC107843451 |
| UFGT | Flavonoid3-O-glucosyltransferase | UFGT | 10 | LOC107843659 |
| MT | Methyltransferase | PMT2 | 1 | LOC107877679 |
| RT | Rhamnosyltransferase | 3 | LOC107862351 | |
| AT | Acyltransferase | 3 | LOC107863748 | |
| GST | Glutathione S-transferase | GST | 9 | LOC107842862 |
| Short Name | Enzyme Name | Gene Name | Chromosome Location | Gene ID |
|---|---|---|---|---|
| GGPS | Geranylgeranyl pyrophosphosphate synthase | Ggps | 4 | LOC107867046 |
| PSY | Phytoene synthase | Psy | 4 | LOC107868281 |
| PDS | Phytoene desaturase | Pds | 3 | LOC107861625 |
| ZDS | ζ-carotene desaturase | Zds | 8 | LOC107839468 |
| LYC | Lycopene cyclase | CrtL | 5 | LOC107869983 |
| CRTZ | β-carotene hydroxylase | CrtZ-2 | 3 | LOC107863219 |
| CrtZ-1 | 6 | LOC107873401 | ||
| Ze | Zeaxanthin epoxidase | Ze | 2 | LOC107860302 |
| CCS | Capsanthin-capsorubin synthase | Ccs | 6 | LOC107875664 |
| Gene Name | Color | Functional Analysis | Chromosome |
|---|---|---|---|
| pc1 (pc8.1, LOL1) | Green, white | Affect chlorophyll content | 1 |
| pc10.1 (CaGLK2) | Green, white | Involved in chloroplast development | 2 |
| APRR2-Like | Green, white | Affect chlorophyll content | 2 |
| A (CaMYB) | Violet, nearly black | Regulate anthocyanin synthesis | 10 |
| MoA | Violet | Adjust the shade of violet | 11 |
| CCS (Y+) | Red, yellow | Regulate capsaicin synthesis | 6 |
| PSY (C2) | Orange | Regulate carotenoid synthesis | 7 |
| CaPRR2 (C1) | White, green, brown | Regulate carotenoid synthesis | 1 |
| CaSGR | White, green, brown | Chlorophyll degradation | 1 |
| CarbcL | White, green, brown | Regulate chlorophyll and capsaicin synthesis | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhong, Y.; Liu, J.; Ma, R.; Miao, Y.; Chen, W.; Zheng, J.; Pang, X.; Wan, H. Pigment Biosynthesis and Molecular Genetics of Fruit Color in Pepper. Plants 2023, 12, 2156. https://doi.org/10.3390/plants12112156
Wang L, Zhong Y, Liu J, Ma R, Miao Y, Chen W, Zheng J, Pang X, Wan H. Pigment Biosynthesis and Molecular Genetics of Fruit Color in Pepper. Plants. 2023; 12(11):2156. https://doi.org/10.3390/plants12112156
Chicago/Turabian StyleWang, Linlin, Yangmin Zhong, Jia Liu, Ruifang Ma, Yeminzi Miao, Wenqi Chen, Jiaqiu Zheng, Xin Pang, and Hongjian Wan. 2023. "Pigment Biosynthesis and Molecular Genetics of Fruit Color in Pepper" Plants 12, no. 11: 2156. https://doi.org/10.3390/plants12112156