
Biomimetic Strategies for Developing Abiotic Stress-Tolerant Tomato Cultivars: An Overview

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
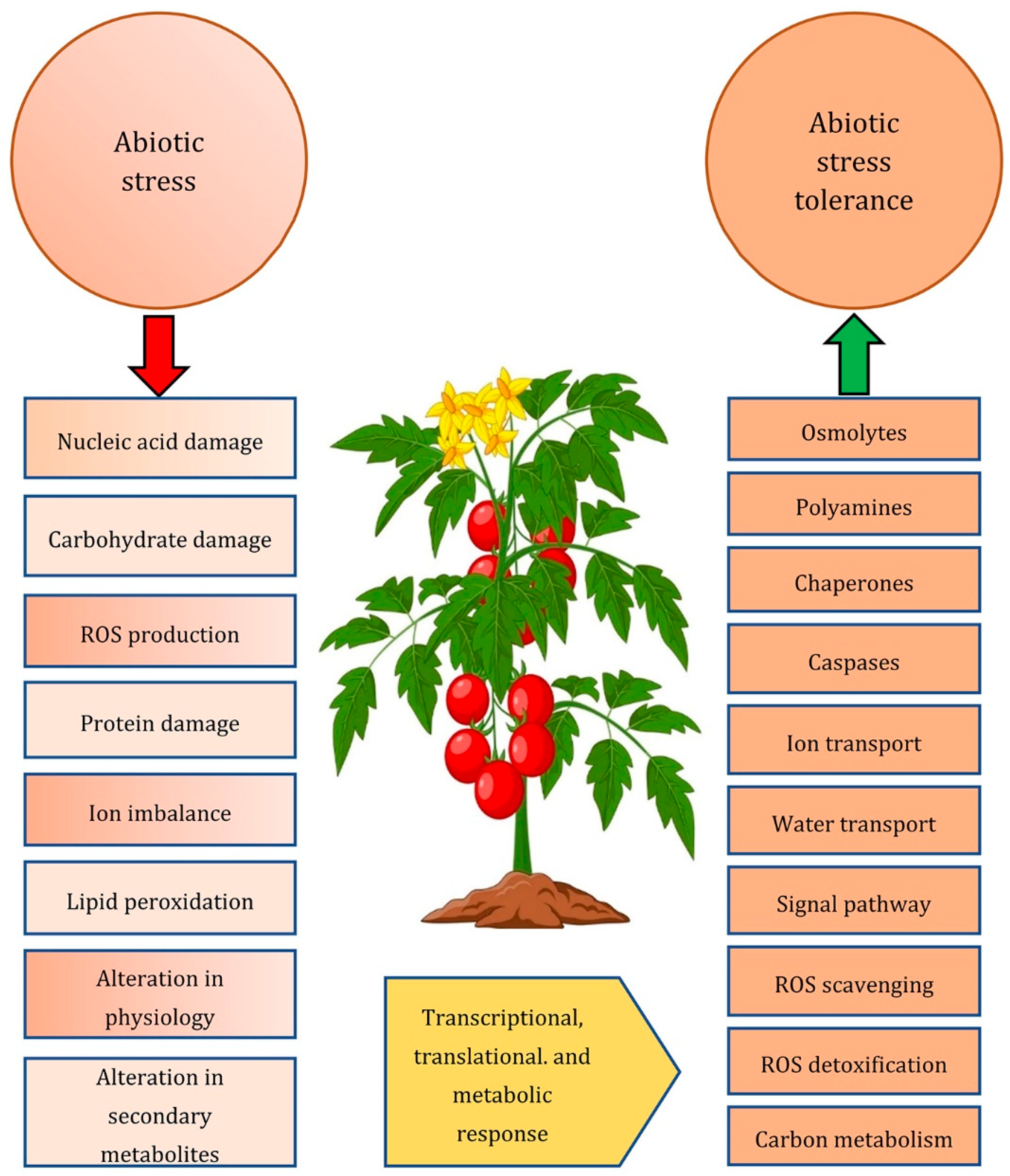
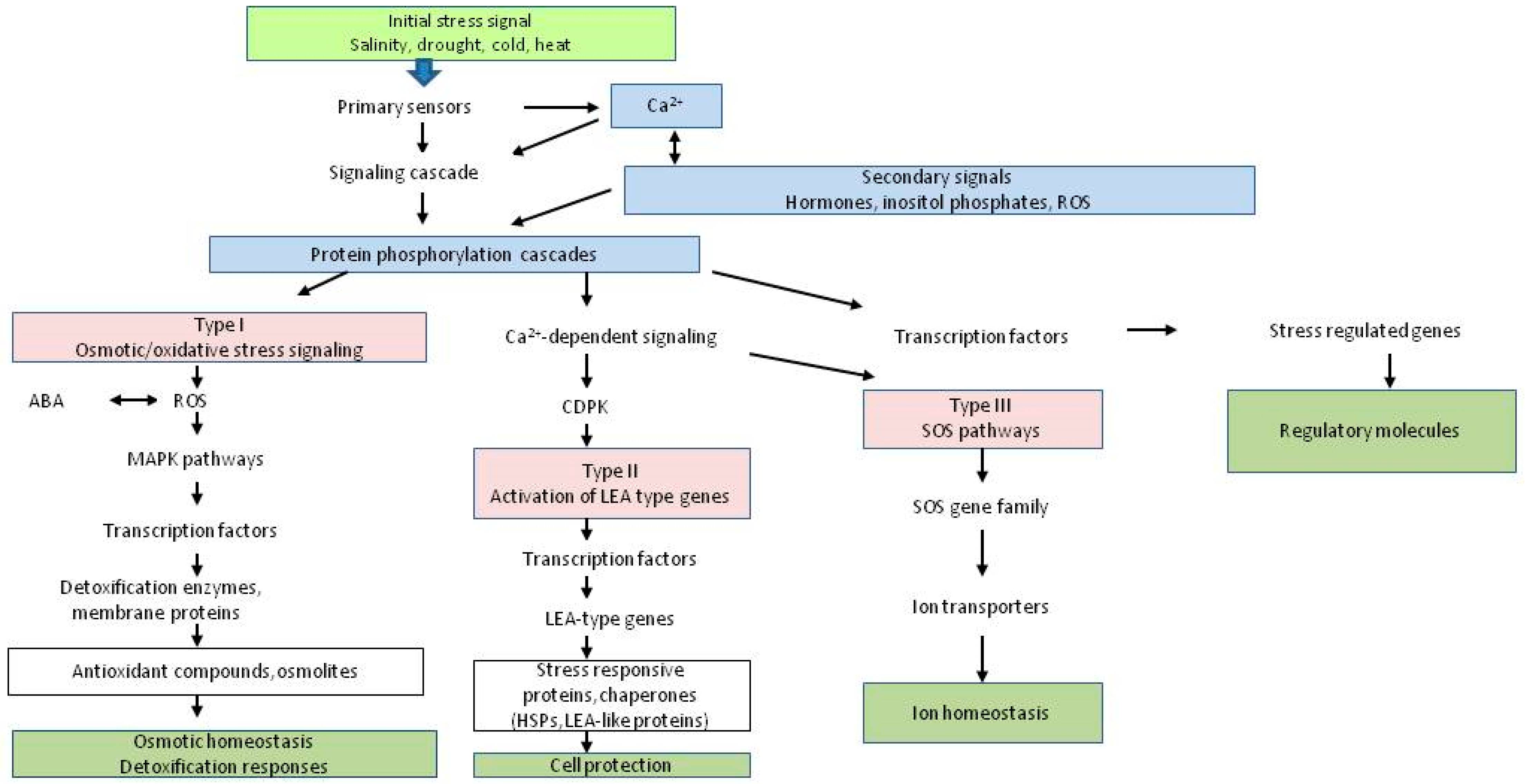
2. Abiotic Stress Resistance at the Physiological Level
3. Biotechnological Tools to Develop Tolerant Plants against Abiotic Stress
3.1. Genetic Transformation Methods in Tomato
3.2. Transformation Approaches Using rDNA Technologies (Genetic Engineering)
3.2.1. Mannitol
3.2.2. Glycine Betaine
3.2.3. Glutathione
3.2.4. Osmotin
3.2.5. Polyamines
3.2.6. Trehalose
3.2.7. Biosynthesis of Ethylene
3.2.8. Aquaporins
3.2.9. Heat Shock Proteins
3.2.10. Antioxidants
3.2.11. Ion Transport Proteins
4. Genome Editing for Ameliorating Abiotic Stresses in Tomato
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khapte, P.S.; Kumar, P.; Saxena, A.; Singh, A. Performance evaluation and character association studies in arid region greenhouse tomato hybrids. Indian J. Hortic. 2018, 75, 457–462. [Google Scholar] [CrossRef]
- FAOSTAT. 2020. Available online: http://faostat3.fao.org/home/E (accessed on 12 September 2022).
- Singh, H.; Kumar, P.; Chaudhari, S.; Edelstein, M. Tomato grafting: A global perspective. HortScience 2017, 52, 1328–1336. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, Z.; Rashid, H. An improved Agrobacterium-mediated transformation in tomato using hygromycin as a selective agent. Afr. J. Biotechnol. 2010, 9, 1882–1891. [Google Scholar]
- Sanwal, S.K.; Mann, A.; Kumar, A.; Kesh, H.; Kaur, G.; Rai, A.K.; Kumar, R.; Sharma, P.C.; Kumar, A.; Bahadur, A.; et al. Salt Tolerant Eggplant Rootstocks Modulate Sodium Partitioning in Tomato Scion and Improve Performance under Saline Conditions. Agriculture 2022, 12, 183. [Google Scholar] [CrossRef]
- Behera, T.K.; Krishna, R.; Ansari, W.A.; Aamir, M.; Kumar, P.; Kashyap, S.P.; Pandey, S.; Kole, C. Approaches Involved in the Vegetable Crops Salt Stress Tolerance Improvement: Present Status and Way Ahead. Front. Plant Sci. 2022, 12, 787292. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Kumar, P.; Cardarelli, M.; Rouphael, Y. Grafting an effective tool for abiotic stress alleviation in vegetables. In Horticulture for Food and Environment Security; Chadha, K.L., Singh, A.K., Singh, S.K., Dhillon, W.S., Eds.; West Ville Pub. House: New Delhi, India, 2013; pp. 15–28. [Google Scholar]
- Rouphael, Y.; Venema, J.H.; Edelstein, M.; Savvas, D.; Colla, G.; Ntatsi, G.; Kumar, P.; Schwarz, D. Grafting as a tool for tolerance of abiotic stress. In Vegetable Grafting: Principles and Practices; Colla, G., Perez-Alfocea, F., Schwarz, D., Eds.; CAB International: Wallingford, UK, 2017; pp. 171–215. [Google Scholar]
- Lin, K.H.; Lo, H.F.; Lee, S.P.; Kuo, C.G.; Chen, J.T. RAPD markers for the identification of yield traits in tomatoes under heat stress via bulk segregant analysis. Heredity 2006, 143, 142–154. [Google Scholar] [CrossRef]
- Ranjan, A.; Ichihashi, Y.; Sinha, N.R. The tomato genome: Implications for plant breeding, genomics and evolution. Genome Biol. 2012, 13, 167. [Google Scholar] [CrossRef]
- Chen, Q.; Lai, H. Gene delivery into plant cells for recombinant protein production. BioMed Res. Int. 2015, 2015, 932161. [Google Scholar] [CrossRef] [Green Version]
- Low, L.; Yang, S.; Kok, D.A.; Ong-Abdullah, J.; Tan, N.; Lai, K. Transgenic Plants: Gene Constructs, Vector and Transformation Method. In New Visions in Plant Science; IntechOpen: London, UK, 2018; pp. 41–61. [Google Scholar] [CrossRef]
- Kissoudis, C.; Chowdhury, R.; van Heusden, S.; van de Wiel, C.; Finkers, R.; Visser, R.G.F.; Bai, Y.; van der Linden, G. Combined biotic and abiotic stress resistance in tomato. Euphytica 2015, 202, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Vegetable grafting as a tool to improve drought resistance and water use efficiency. Front. Plant Sci. 2017, 8, 1130. [Google Scholar] [CrossRef]
- Nicky, J.A.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Zachary, J.F. Mechanisms and Morphology of Cellular Injury, Adaptation, and Death. Pathol. Basis Vet. Dis. 2017, 2017, 2–43.e19. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Rehem, B.C.; Bertolde, Z.F.; de Almeida, A.F. Regulation of gene expression in response to abiotic stress in plants. In Cell Metabolism—Cell Homeostasis and Stress Response; Bubulya, P., Ed.; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. (Ed.) In vivo chlorophyll fluorescence as a tool for stress detection in plants. In Applications of Chlorophyll Fluorescence; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 129–142. [Google Scholar]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. DREB regulons in abiotic-stress-responsive gene expression in plants. In Molecular Breeding of Forage and Turf; Yamada, T., Spangenberg, G., Eds.; Springer: New York, NY, USA, 2009; pp. 15–28. [Google Scholar] [CrossRef]
- Hoang, X.L.T.; Nhi, D.N.H.; Thu, N.B.A.; Thao, N.P.; Tran, L.P. Transcription Factors and Their Roles in Signal Transduction in Plants under Abiotic Stresses. Curr. Genom. 2017, 18, 483–497. [Google Scholar] [CrossRef]
- Ishida, Y.; Hiei, Y.; Komari, T. Tissue culture protocols for gene transfer and editing in maize (Zea mays L.). Plant Biotechnol. 2020, 37, 121–128. [Google Scholar] [CrossRef]
- Gerszberg, A.; Hnatuszko-Konka, K. Tomato tolerance to abiotic stress: A review of most often engineered target sequences. Plant Growth Regul. 2017, 83, 175–198. [Google Scholar] [CrossRef] [Green Version]
- Parmar, N.; Singh, K.H.; Sharma, D.; Singh, L.; Kumar, P.; Nanjundan, J.; Khan, Y.J.; Chauhan, D.K.; Thakur, A.K. Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: A comprehensive review. 3Biotech 2017, 7, 239. [Google Scholar] [CrossRef]
- Hnatuszko-Konka, K.; Kowalczk, T.; Gerszberg, A.; Wiktorek-Smagur, A.; Kononowicz, A.K. Phaseolus vulgaris—Recalcitrant potential. Biotechnol. Adv. 2014, 32, 1205–1215. [Google Scholar] [CrossRef]
- Yang, N.S.; Christou, P. (Eds.) Particle Bombardment Technology for Gene Transfer; Oxford University Press: New York, NY, USA, 1994; pp. 143–165. [Google Scholar]
- Mathews, H.; Wagoner, W.; Cohen, C.; Kellogg, J.; Bestwick, R. Efficient genetic transformation of red raspberry, Rubus ideaus L. Plant Cell Rep. 1995, 14, 471–476. [Google Scholar] [CrossRef]
- Hilder, V.A.; Boulter, D. Genetic engineering of crop plants for insect resistance—A critical review. Crop Prot. 1999, 18, 177–191. [Google Scholar] [CrossRef]
- Gosal, S.S.; Gosal, S.K. Genetic transformation and production of transgenic plants. In Plant Biotechnology—Recent Advances; Trivedi, P.C., Ed.; Panima Publishers: New Delhi, India, 2000; pp. 29–40. [Google Scholar]
- Chahal, G.S.; Gosal, S.S. Principles and Procedures of Plant Breeding: Biotechnological and Conventional Approaches; Narosa Publishing House: New Delhi, India, 2002. [Google Scholar]
- Altman, A. From plant tissue culture to biotechnology: Scientific revolutions, abiotic stress and new advances in crop improvement. Theor. Appl. Genet. 2003, 129, 1639-16. [Google Scholar]
- Grewal, D.K.; Gill, R.; Gosal, S.S. Genetic engineering of Oryza sativa by particle bombardment. Biol. Plant 2006, 50, 311–314. [Google Scholar] [CrossRef]
- Kerr, A. GM crops—A minireview. Australas. Plant Pathol. 2011, 40, 449–452. [Google Scholar] [CrossRef]
- Kamthan, A.; Chaudhuri, A.; Kamthan, M.; Datta, A. Genetically modified (GM) crops: Milestones and new advances in crop improvement. Theor. Appl. Genet. 2016, 129, 1639–1655. [Google Scholar] [CrossRef] [PubMed]
- Arora, L.; Narula, A. Gene editing and crop improvement using CRISPR-Cas9 system. Biol. Plant 2017, 50, 311–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanuja, P.; Kumar, A.L. Transgenic fruit crops—A review. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2030–2037. [Google Scholar] [CrossRef] [Green Version]
- McCormick, S.; Niedermeyer, J.; Fry, J.; Barnason, A.; Horsch, R.; Fraley, R. Leaf disc transformation of cultivated tomato (L. esculentum) using Agrobacterium tumefaciens. Plant Cell Rep. 1986, 2, 81–84. [Google Scholar] [CrossRef]
- Gerszberg, A.; Hnatuszko-Konka, K.; Kowalczyk, T.; Kononowicz, A.K.K. Tomato (Solanum lycopersicum L.) in the service of biotechnology. Plant Cell Tissue Organ Cult. 2015, 120, 881–902. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.H.; Ali, S.; Jan, S.A.; Din, A.U.; Ali, G.M. Piercing and incubation method of in planta transformation producing stable transgenic plants by overexpressing DREB1A gene in tomato (Solanum lycopersicum Mill.). Plant Cell Tissue Organ Cult. 2015, 120, 1139–1157. [Google Scholar] [CrossRef]
- Sun, S.X.P.; Xing, X.J.; Xu, X.Y.; Cheng, J.; Zheng, S.W.; Xing, G.M. Agrobacterium-mediated transformation of tomato (Lycopersicon esculentum L. cv. Hezuo908) with improved efficiency. Biotechnol. Biotechnol. Equip. 2015, 29, 861–868. [Google Scholar] [CrossRef]
- Chetty, V.J.; Ceballos, N.; Garcia, D.; Narvaez-Vasquez, J.; Lopez, W.; Orozco-Cardenas, M.L. Evaluation of four Agrobacterium tumefaciens strains for the genetic transformation of tomato (Solanum lycopersicum L.) cultivar Micro-Tom. Plant Cell Rep. 2013, 32, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Cueno, M.C.; Hibi, Y.; Karamatsu, K.; Yasutomi, Y.; Imai, K.; Laurena, A.C.; Okamoto, T. Preferential expression and immunogenicity of HIV-1 Tatfusion protein expressed in tomato plant. Transg. Res. 2010, 19, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Ruma, D.; Dhaliwal, M.S.; Kaur, A.; Gosal, S.S. Transformation of tomato using biolistic gun for transient expression β glucuronidase gene. Indian J. Biotechnol. 2009, 8, 363–369. [Google Scholar]
- Hasan, M.; Khan, A.J.; Khan, S.S.; Shah, A.H.; Khan, A.R.; Mirza, B. Transformation of tomato (Lycopesricon esculentum Mill.) with Arabidopsis early flowering gene APETALI (API) through Agrobacterium infiltration of ripened fruits. Pak. J. Bot. 2008, 1, 161–173. [Google Scholar]
- Guo, M.; Zhang, Y.L.; Meng, Z.J.; Jiang, J. Optimization of factors affecting Agrobacterium-mediated transformation of MicroTom tomatoes. Genet. Mol. Res. 2012, 1, 661–671. [Google Scholar] [CrossRef]
- Moghaieb, R.E.; Saneoka, H.; Fujita, K. Shoot regeneration from GUS-transformed tomato (Lycopersicon esculentum) hairy root. Cell. Mol. Biol. 2004, 9, 439–449. [Google Scholar]
- Abu-El-Heba, G.A.; Hussein, G.M.; Abdalla, N.A. A rapid and efficient tomato regeneration and transformation system. Agric. For. Res. 2008, 58, 103–110. [Google Scholar]
- Fuentes, A.D.; Ramos, P.L.; Sanchez, Y.; Callard, D.; Ferreira, A.; Tiel, K.; Cobas, K.; Rodriguez, R.; Borroto, C.; Doreste, V.; et al. A transformation procedure for recalcitrant tomato by addressing transgenic plant-recovery limiting factors. Biotechnol. J. 2008, 3, 1088–1093. [Google Scholar] [CrossRef]
- Sharma, M.K.; Solanke, A.U.; Jani, D.; Singh, Y.; Sharma, A.K. Asimple and efficient Agrobacterium-mediated procedure for transformation of tomato. J. Biosci. 2009, 3, 423–433. [Google Scholar] [CrossRef]
- Janani, C.; Girija, S.; Ranjitha Kumari, B.D. In vitro culture and Agrobacterium-mediated transformation in high altitude tomato (Lycopersicon esculentum Mill.) cultivar Shalimar. Int. J. Pharm. Teach. Pract. 2013, 4, 483–488. [Google Scholar]
- Cruz-Mendıvil, A.; Rivera-Lopez, J.; German-Baez, L.J.; Lopez-Meyer, M.; Hernandez-Verdugo, S.; Lopez-Valenzuela, J.A.; Reyes-Moreno, C.; Valdez-Ortiz, A. A simple and efficient protocol for plant regeneration and genetic transformation of tomato cv. Micro-Tom from leaf explants. HortScience 2011, 46, 1655–1660. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Bansal, K.C. Efficient production of transgenic tomatoes via Agrobacterium-mediated transformation. Biol. Plant. 2010, 54, 344–348. [Google Scholar] [CrossRef]
- El-Siddig, M.A.; El-Hussein, A.A.; Saker, M.M. Agrobacterium-mediated transformation of tomato plants expressing defensin gene. Int. J. Agric. Res. 2011, 4, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Metwali, E.M.R.; Soliman, H.I.A.; Fuller, M.P.; Almaghrabi, O.A. Improving fruit quality in tomato (Lycoper sicum esculentum Mill) under heat stress by silencing the vis 1 gene using small interfering RNA technology. Plant Cell Tissue Organ Cult. 2015, 121, 153–166. [Google Scholar] [CrossRef]
- Wang, T.; Wen, L.W.; Zhu, H.L. Effectively organ-specific virus induced gene silencing in tomato plant. J. Nat. Sci. 2015, 1, 34. [Google Scholar]
- Mishra, K.B.; Ianacone, R.; Petrozza, A.; Mishra, A.; Armentano, N.; La Vecchia, G.; Trtilek, M.; Cellini, F.; Nedbal, L. Engineered drought tolerance in tomato plants is reflected in chlorophyll fluorescence emission. Plant Sci. 2012, 182, 79–86. [Google Scholar] [CrossRef]
- Vincour, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef]
- Baranova, E.N.; Serenko, E.K.; Balachnina, T.I.; Kosobruhov, A.A.; Kurenina, L.V.; Gulevich, A.A.; Maisuryan, A.N. Activity of the photosynthetic apparatus and antioxidant enzymes in leaves of transgenic Solanum lycopersicum and Nicotiana tabacum plants, with FeSOD1 gene. Russ. Agric. Sci. 2010, 36, 242–249. [Google Scholar] [CrossRef]
- Alvarez-Gomez, T.B.; Ramirez-Trujillo, J.A.; Ramirez-Yanez, M.; Suarez-Rodriguez, R. Overexpression of SIERF3b and SIERF5 in transgenic tomato alters fruit size, number of seeds and promotes early flowering, tolerance to abiotic stress and resistance to Botrytis cinereal infection. Ann. Appl. Biol. 2021, 179, 382–394. [Google Scholar] [CrossRef]
- Dominguez, T.; Hernandez, L.M.; Pennycooke, J.C.; Jimenez, P.; Martınez-Rivas, J.M.; Sanz, C.; Stockinger, E.J.; Sanchez-Serrano, J.; Sanmartin, M. Increasing v-3 desaturase expression in tomato results in altered aroma profile and enhanced resistance to cold stress. Plant Physiol. 2010, 153, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Wang, X.; Wei, M.; Yang, F.; Li, Y.; Shi, Q. Overexpression of S-adenosylmethionine synthetase1 enhances tomato callus tolerance to alkali stress through polyamine and hydrogen peroxide cross-linked networks. Plant Cell Tissue Organ Cult. 2015, 124, 377–391. [Google Scholar] [CrossRef]
- Yang, D.Y.; Li, M.; Ma, N.N.; Yang, X.H.; Meng, Q.W. Tomato SlGGP-LIKE gene participates in plant responses to chilling stress and pathogenic infection. Plant Physiol. Biochem. 2017, 112, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Meng, X.; Cai, J.; Li, G.; Dong, T.; Li, Z. Basic leucine zipper transcription factor SlbZIP1 mediates salt and drought stress tolerance in tomato. BMC Plant Biol. 2018, 10, 83. [Google Scholar] [CrossRef] [Green Version]
- Yuling Bai, Y.B.; Sri Sunarti, S.S.; Christos Kissoudis, C.K.; Richard, G.F.; Visser, C.G.; van der Linden, C. The Role of Tomato WRKY Genes in Plant Responses to Combined Abiotic and Biotic Stresses. Plant Sci. 2018, 9, 801. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wu, M.; Teng, Y.; Jia, S.; Yu, D.; Wei, T.; Chen, C.; Song, W. Overexpression of the Glutathione Peroxidase 5 (RcGPX5) Gene from Rhodiola crenulata Increases Drought Tolerance in Salvia miltiorrhiza. Plant Sci. 2019, 9, 1950. [Google Scholar] [CrossRef] [PubMed]
- Fernando Novillo, F.N.; Joaquın Medina, J.M.; Julio Salinas, J.S. Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulation. Proc. Natl. Acad. Sci. USA 2007, 104, 21002–21007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.G.; Wang, S.H.; Luo, H.; Ma, Q.J.; Yao, Y.X.; You, C.X.; Hao, Y.J. Overexpression of MdVHA-B, a V-ATPase gene from apple, confers tolerance to drought in transgenic tomato. Sci. Hortic. 2012, 145, 94–101. [Google Scholar] [CrossRef]
- Hu, D.G.; Ma, Q.J.; Sun, C.H.; Sun, M.H.; You, C.X.; Hao, Y.J. Overexpression of MdSOS2L1, a CIPK protein kinase, increases the antioxidant metabolites to enhance salt tolerance in apple and tomato. Physiol. Plant 2015, 156, 201–214. [Google Scholar] [CrossRef]
- Rahman, M.A.; Wu, W.; Yan, Y.; Shamsul, A. Bhuiyan Overexpression of TERF1 in sugarcane improves tolerance to drought stress. Crop Pasture Sci. 2021, 72, 268–279. [Google Scholar] [CrossRef]
- Huertas, R.; Rubio, L.; Cagnac, O.; Garcia-Schanchez, M.J.; De Dios Alche, J.; Venema, K.; Fernandez, J.A.; Rodriguez-Rosales, M.P. The K+/H+ antiporter LeNHX2 increases salt tolerance by improving K+ homeostasis in transgenic tomato. Plant Cell Environ. 2013, 36, 2135–2149. [Google Scholar] [CrossRef]
- Li, Z.; Palmer, W.M.; Antony, P.; Wang, M.R.; Rainsford, F.; Jin, Y.; Patrick, J.W.; Yang, Y.; Ruan, Y.-L. High invertase activity in tomato reproductive organs correlates with enhanced sucrose import into, and heat tolerance of, young fruit. J. Exp. Bot. 2011, 63, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Khare, N.; Goyary, D.; Kumar Singh, N.; Shah, P.; Rathore, M.; Anandhan, S.; Sharma, D.; Arif, M.; Ahmed, Z. Transgenic tomato cv. Pusa uphar expressing a bacterial mannitol-1-phosphate dehydrogenase gene confers abiotic stress tolerance. Plant Cell Tissue Organ Cult. 2010, 103, 267–277. [Google Scholar] [CrossRef]
- Liu, X.; Yang, J.H.; Li, B.; Yang, X.M.; Meng, Q.W. Antisense expression of tomato chloroplast omega-3 fatty acid desaturase gene (LeFAD7) enhances the tomato high-temperature tolerance through reductions of trienoic fatty acids and alterations of physiological parameters. Photosyntheyhica 2010, 1, 59–66. [Google Scholar] [CrossRef]
- Lyu, J.I.; Min, S.R.; Lee, J.H.; Lim, Y.H.; Kim, J.K.; Bae, C.H.; Liu, J.R. Overexpression of a trehalose-6-phosphate synthase/phosphatise fusion gene enhances tolerance and photosynthesis during drought and salt stress without growth aberrations in tomato. Plant Cell Tissue Organ Cult. 2013, 112, 257–262. [Google Scholar] [CrossRef]
- Miura, K.; Shiba, H.; Ohta, M.; Seung Kang, W.; Sato, A.; Yuasa, T.; Iwaya-Inoue, M.; Kamada, H.; Ezura, H. SlICE1 encoding a MYC-type transcription factor controls cold tolerance intomato, Solanum lycopersicum. Plant Biotechnol. 2012, 29, 253–260. [Google Scholar] [CrossRef]
- Mohamed, E.A.; Iwaki, T.; Munir, I.; Tamoi, M.; Shigeoka, S.; Wadano, A. Overexpression of bacterial catalase in tomato leaf chloroplasts enhances photo-oxidative stress tolerance. Plant Cell Environ. 2003, 26, 2037–2046. [Google Scholar] [CrossRef]
- Muñoz-Mayor, M.F.; Pineda, B.; Garcia-Abellan, J.O.; Antón, T.; GarciaSogo, B.; Sanchez-Bel, P.; Flores, F.B.; Atarés, A.; Angosto, T.; Pintor-Toro, J.A.; et al. Overexpression of dehydrin tas14 gene improves the osmotic stress imposed by drought and salinity in tomato. J. Plant Physiol. 2012, 169, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Jenknic, Z.; Chen, T.H.H. Exogenous application of glycinebetaine increases chilling tolerance in tomato plants. Plant Cell Physiol. 2006, 6, 706–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, A.C.; Singh, M.; Shah, K. Engineering drought tolerant tomato plants over-expressing BcZAT12 gene encoding a C2H2 zinc finger transcription factor. Phytochemistry 2013, 85, 44–50. [Google Scholar] [CrossRef]
- Seong, E.S.; Cho, H.S.; Choi, D.; Joung, Y.H.; Lim, C.K.; Hur, J.H.; Wang, M.H. Tomato plants overexpressing CaKR1 enhanced tolerance to salt and oxidative stress. Biochem. Biophys. Res. Commun. 2007, 363, 983–988. [Google Scholar] [CrossRef]
- Shah, K.; Singh, M.; Rai, A.C. Effect of heat-shock induced oxidative stress is suppressed in BcZAT12 expressing drought tolerant tomato. Phytochemistry 2013, 95, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.Q.; Zhang, Q.F.; Liu, J.H.; Li, G.H. Overexpression of PtADC confers enhanced dehydratation and drought tolerance in transgenic tobacco and tomato: Effect on ROS elimination. Biochem. Biophys. Res. Commun. 2011, 413, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Yu, C.; Tang, X.F.; Zhu, Z.J.; Ma, N.N.; Meng, Q.W. Atomato endoplasmic reticulum (ER)-type omega-3 fatty acidd esaturase (LeFAD3) functions in early seedling tolerance to salinity stress. Plant Cell Rep. 2014, 33, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Yarra, R.; He, S.J.; Abbagani, S.; Ma, B.; Bulle, M.; Zhang, W.K. Overexpression of wheat Na+/H+ antiporter gene (TaNHX2) enhances tolerance to salt stress in transgenic tomato plants (Solanum lycopersicum L.). Plant Cell Tissue Organ Cult. 2012, 111, 49–57. [Google Scholar] [CrossRef]
- Saha, B.C.; Racine, F.M. Effects of pH and corn steep liquor variability on manitol production by Lactobaccilus. NRRL B-3693. Appl. Microb. Biotechnol. 2010, 87, 553–560. [Google Scholar] [CrossRef]
- Dai, Q.M.; Mu, W.; Zhang, T. Recent advances in the applications and biotechnological production of manitol. J. Func. Foods 2017, 36, 404–409. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Glycine betaine protects plants against abiotic stress: Mechanisms and biotechnological applications. Plant Cell Environ. 2010, 34, 1–20. [Google Scholar] [CrossRef]
- Park, E.J.; Jeknić, Z.; Sakamoto, A.; DeNoma, J.; Yuwansiri, R.; Murata, N.; Chen, T.H.H. Genetic engineering of glycinebetaine synthesis in tomato protects seeds, plants, and flowers from chilling damage. Plant J. 2004, 40, 474–487. [Google Scholar] [CrossRef]
- Hossain, M.A.; Piyatida, P.; Teixeira da Silva, J.A.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 2012, 872875. [Google Scholar] [CrossRef] [Green Version]
- Herbette, S.; Tourvieille de Labrouheb, D.; Drevet, J.R.; Roeckel-Drevet, P. Transgenic tomatoes showing higher glutathione peroxydase antioxidant activity are more resistant to an abiotic stress but more susceptible to biotic stresses. Plant Sci. 2011, 180, 548–553. [Google Scholar] [CrossRef]
- Alvarez-Viveros, M.F.; Inostroza-Blancheteau, C.; Timmermann, T.; González, M.; Arce-Johnson, P. Overexpression of GlyI and GlyII genes in transgenic tomato (Solanum lycopersicum Mill.) plants confers salt tolerance by decreasing oxidative stress. Mol. Biol. Rep. 2013, 4, 3281–3290. [Google Scholar] [CrossRef]
- Ram, C.; Danish, S.; Kesawat, M.S.; Panwar, B.S.; Verma, M.; Arya, A.; Yadav, S.; Sharma, V. Genome-wide comprehensive characterization and expression analysis of TLP gene family revealed its responses to hormonal and abiotic stresses in watermelon (Citrullus lanatus). Gene 2022, 844, 146818. [Google Scholar] [CrossRef] [PubMed]
- Goel, D.; Singh, A.K.; Yadav, V.; Babbar, S.B.; Bansal, K.C. Overexpression of osmotin gene confers tolerance to salt and drought stresses in transgenic tomato (Solanum lycopersicum L.). Protoplasma 2010, 245, 133–141. [Google Scholar] [CrossRef]
- Viktorova, J.; Klcova, B.; Rehorova, K.; Vlcko, T.; Stankova, L.; Jelenova, N.; Cejnar, P.; Kundu, J.K.; Ohnoutkova, L.; Macek, T. Recombinant expression of osmotin in barley improves stress resistance. PLoS ONE 2019, 14, e0212718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and abiotic stress in plants: A complex relationship. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef] [PubMed]
- Alćazar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Carrasco, P.; Tiburcio, A.F. Polyamines: Molecules with regulatory functions in plant abiotic stress tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef]
- Kumar, R.R.; Sharma, S.K.; Rai Gyanendra, K.; Sing, K.; Choudhury, M.; Dhawan, G.; Singh, G.P.; Goswami, S.; Pathak, H.; Rai, R.D. Exogenous application of putrescine at pre anthesis enchances the thermotolerance of wheat (Triticum aestivum L.). Indian J. Biochem. Biophys. 2014, 51, 396–406. [Google Scholar]
- Arguelles, J.C. Physiological roles of trehalose in bacteria and yeasts: A comparative analysis. Arch. Microbiol. 2000, 174, 217–224. [Google Scholar]
- Cortina, C.; Culianez-Macia, F.A. Tomato abiotic stress enhanced tolerance by trehalose biosynthesis. Plant Sci. 2005, 169, 75–82. [Google Scholar] [CrossRef]
- Gururani, M.A.; Tapan Mohanta, K.; Bae, H. Current understanding of the interplay between phytohormones and photosynthesis under environmental stress. Int. J. Mol. Sci. 2015, 16, 19055–19085. [Google Scholar] [CrossRef] [Green Version]
- Grichko, V.P.; Glick, B.R. Flooding tolerance of transgenic tomato plants expressing the bacterial enzyme ACC deaminase controlledby the 35S, rolD or PRB-1b promoter. Plant Physiol. Biochem. 2001, 39, 19–25. [Google Scholar] [CrossRef]
- Bienert, G.P.; Bienert, M.D.; Jahn, T.P.; Boutry, M.; Chaumont, F. Solanaceae XIPs are plasma membrane aquaporins that facilitate the transport of many uncharged substrates. Plant J. 2011, 66, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Raturi, G.; Kumawat, S.; Mandlik, R.; Duhan, D.; Thakral, V.; Sudhakaran, S.; Ram, C.; Sonah, H.; Deshmukh, R. Deciphering the role of aquaporins under different abiotic stress conditions in watermelon (Citrullus lanatus). J. Plant Growth Regul. 2022; preprint. [Google Scholar] [CrossRef]
- Reuscher, S.; Akiyama1, M.; Mori1, C.; Aoki, K.; Shibata, D.; Shiratake, K. Genome-wide identification and expression analysis of aquaporins in tomato. PLoS ONE 2013, 8, e79052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sade, N.; Vinocur, B.J.; Diber, A.; Shatil, A.; Ronen, G.; Nissan, H.; Wallach, R.; Karchi, H.; Moshelion, M. Improving plant stress tolerance and yield production: Is the tonoplast aquaporin SlTIP2;2 a key to isohydric to anisohydric conversion. New Phytol. 2009, 181, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Q.T.; Lei, Q.; Feng, C.; Zheng, X.; Zhou, F.; Li, L.; Liu, X.; Wang, Z.; Kong, J. Ectopically expressing MdPIP1;3, an aquaporin gene, increased fruit size and enhanced drought tolerance of transgenic tomatoes. BMC Plant Biol. 2017, 17, 246. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Wang, J.; Li, S.; Zhang, L.; Qi, C.; Weeda, S.; Zhao, B.; Ren, S.; Guo, Y.D. Plasma membrane intrinsic proteins SlPIP2; 1, SlPIP2; 7 and SlPIP2; 5 conferring enhanced drought stress tolerance in tomato. Sci. Rep. 2016, 6, 31814. [Google Scholar] [CrossRef]
- Ouziad, F.; Wilde, P.; Schmelzer, E.; Hildebrandt, U.; Bothe, H. Analysis of expression of aquaporins and Na+/H+ transporters in tomato colonized by arbuscular mycorrhizal fungi and affected by salt stress. Environ. Exp. Bot. 2006, 57, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Al-Whaibi, H.M. Plant heat-shock proteins: A mini review. J. King Saud. Univ.-Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Sabehat, A.; Weiss, D.; Lurie, S. The correlation between heatshock protein accumulation and persistence and chilling in tomato fruit. Plant Physiol. 1996, 110, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Kadyrzhanova, D.K.; Vlachonasios, K.E.; Ververidis, P.; Dilley, D.R. Molecular cloning of a novel heat induced/chilling tolerance related cDNA1 in tomato fruit by use of mRNA differential display. Plant Mol. Biol. 1998, 36, 885–895. [Google Scholar] [CrossRef]
- Zhao, C.; Mariko, M.; Sun, A.; Yi, S.; Li, M.; Liu, J. Constitutive expression of an endoplasmic reticulum small heat shock protein alleviates endoplasmic reticulum stress in transgenic tomato. J. Plant Physiol. 2007, 164, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Li, B.; Hu, Y.G.; Chen, L.; Min, D.H. The wheat E subunit of V-Type H+-ATPase is involved in the plant response to osmotic stress. Int. J. Mol. Sci. 2014, 15, 16196–16210. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Cai, G.; Xu, N.; Zhang, L.; Sun, X.; Guan, J.; Meng, Q. Novel DnaJ Protein Facilitates Thermotolerance of Transgenic Tomatoes. Int. J. Mol. Sci. 2019, 20, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, M.; Feng, H.L.; Wang, L.Y.; Li, D.; Meng, Q.W. Overexpression of thylakoidal ascorbate peroxidase shows enhanced resistance to chilling stress in tomato. J. Plant Physiol. 2012, 169, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Gamper, N.; Shapiro, M. Regulation of ion transport proteins by membrane phosphoinositides. Nat. Rev. Neurosci. 2007, 8, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Gisbert, C.; Rus, A.M.; Boların, C.M.; Lopez-Coronado, J.M.; Arrillaga, I.; Montesinos, C.; Caro, M.; Serrano, R.; Moreno, V. The yeast HAL1 gene improves salt tolerance of transgenic tomato. Plant Physiol. 2000, 123, 393–402. [Google Scholar] [CrossRef]
- Leidi, E.O.; Barragan, V.; Rubio, L.; El-Hamdaoui, A.; Ruiz, T.M.; Cubero, B.; Fernandez, J.A.; Bressan, R.A.; Hasegawa, P.H.; Quintero, F.J.; et al. The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J. 2010, 61, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Lenaerts, B.; Collard, B.C.Y.; Demont, M. Review: Improving global food security through accelerated plant breeding. Plant Sci. 2019, 287, 110207. [Google Scholar] [CrossRef]
- Ahmar, S.; Gill, R.A.; Jung, K.H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef] [Green Version]
- Kaur, N.; Sharma, S.; Hasanuzzaman, M.; Pati, P.K. Genome Editing: A Promising Approach for Achieving Abiotic Stress Tolerance in Plants. Int. J. Genom. 2022, 2022, 5547231. [Google Scholar] [CrossRef] [PubMed]
- Tran, M.T.; Doan, D.T.H.; Kim, J.; Song, Y.J.; Sung, Y.W.; Das, S.; Kim, E.J.; Son, G.H.; Kim, S.H.; Van Vu, T.; et al. CRISPR/Cas9-based precise excision of SlHyPRP1 domain (s) to obtain salt stress-tolerant tomato. Plant Cell Rep. 2021, 40, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, L.; Wang, L.; Chen, L.; Zhao, R.; Sheng, J.; Shen, L. Reduction of tomato-plant chilling tolerance by CRISPR–Cas9-mediated SlCBF1 mutagenesis. J. Agric. Food Chem. 2018, 66, 9042–9051. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, C.; Zhao, R.; Wang, L.; Chen, L.; Yu, W.; Zhang, S.; Shen, J.; Shen, L. CRISPR/Cas9-mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant Biol. 2019, 19, 38. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| S. lycopersicum Cultivar | Transformation Method | Type of Explant | Transformation Frequency (TF) | References |
|---|---|---|---|---|
| Micro-Tom | Indirect | Embryonic part of the seedling | 11% | [41] |
| NA | Indirect | Fruits | 54 to 68.0% | [44] |
| Micro-Tom | Indirect | Cotyledons (embryonic part) | 5.1% | [45] |
| Hezuo 908 | Indirect | Hypocotyls and embryonic part | 40% | [40] |
| Roma and Rio Grande | Indirect | Hypocotyls and leaf disks | 24% and 8%, respectively | [4] |
| Momotaro, UC-97, and Edkawi | Indirect | Hypocotyls | 54 to 67% | [46] |
| Castle Rock | Direct | Hypocotyls and part of cotyledons | 26.5% | [47] |
| Cambell-28 | Indirect | Cotyledons | 21.5% | [48] |
| Pusa Ruby, Sioux, and Arka Vikas | Indirect | Cotyledons | 41.4%, 22%, and 41%, respectively | [49] |
| Hezuo 908 | Indirect | Embryonic part and Hypocotyl | 40% | [40] |
| Shalimar | Indirect | Shoot and Leaf | NA | [50] |
| MicroTom | Indirect | Leaf | 19.1% | [51] |
| NA | Indirect | Hypocotyls | 33 to 59% | [41] |
| Pusa Ruby and DT-93 | Indirect | Cotyledons | higher than 37% | [52] |
| Summer | Indirect | Hypocotyls and cotyledons | 7% | [53] |
| Gene/Origin | Function | Expression | Results | References |
|---|---|---|---|---|
| Fe-SOD/A. thaliana | Lessens the oxidative stress | Upregulation | Increased ability to withstand oxidative stress and improve stability of photosynthetic equipment | [58] |
| SIERF3b & SIERF5 | Regulates transcription for stress conditions | Overexpression | Enhanced tolerance to abiotic stress and resistance to biotic stress | [59] |
| FAD3/rape FAD7/potato | Regulates the fatty acid unsaturation of membrane lipids | Upregulation | Boosted cold resistance; an increase in the 18:3/18:2 ratio in leaves and fruits | [60] |
| SlSAM1/tomato | Promotes the conversion of ATP plus methionine to S-adenosylmethionine, which is necessary for the production of ethylene and PAs | Upregulation | Improved resistance to saline-alkali stress | [61] |
| SIGGP (LIKE)/tomato | Transcription factor | Expression | Enhanced tolerance to abiotic and biotic stress | [62] |
| SIBZIP1/tomato | TF, Defense protein | Downregulation | Regulated ABA-mediated pathway to enhance drought tolerance | [63] |
| SLWRKY | TF, transcriptional regulation | Overexpression | Regulated biotic stress | [64] |
| RcGPX5/Salvimiltiorrhiza | Gluthatione biosynthesis | Overexpression | Tolerance to H2O2, drought and oxidative stress | [65] |
| CBF1/A. thaliana | TF and regulates transcription | Transcription/ regulation | Enhanced cold tolerance | [66] |
| MdVHA-B/apple | Maintains the homeostasis of ion | Upregulation | Improved drought tolerance | [67] |
| MdSOS2L1/apple | Signal-inducing proteins; influence on ion-driving transport mechanisms | Excessive expression | Increased salt tolerance | [68] |
| TERF1/sugarcane | Transcription factor for ethylene response; assimilates ethylene and osmotic stress pathways | Excessive expression | Increased tolerance to drought stress, osmotic stress caused by salt | [69] |
| LeNHX2/tomato | Transport of ions | Expression in excess | Salt tolerance increase | [70] |
| CWIN (Lin7)/tomato | Takes part in mechanisms related to temperature stress | Expresses at normal level | Improved heat resistance in tomato flowers | [71] |
| mt1D/E. coli | Biosynthesis of mannitol | Upregulation | Increased tolerance to drought, cold, and salinity | [72] |
| LeFAD7/tomato | Role in fatty acid transcription | Antisense regulation | Improved high-temperature tolerance; trienoic fatty acids reduced | [73] |
| TPSP (TPS/TPP fusion gene)/E. coli | Biosynthesis of Trehalose | Upregulation | Salt and drought resistance improved | [74] |
| SlICE1/tomato | Transcription regulation | Overexpression | Improved tolerance to cold | [75] |
| katE/tomato | Oxidative stress (catalase) | Overexpression | Upgraded resistance to photo-oxidative stress as a result of drought and Fungal stress | [76] |
| tas14/tomato | Accumulates chaperone-like proteins more effectively | upregulation | Enhanced tolerance to drought and salinity without any growth aberrations | [77] |
| Glycine betaine | Stress savior | Supplements | Chilling tolerance increased | [78] |
| ZAT12/B. carinata | Transcriptional regulation | Upregulation | Boosted resistance toward drought | [79] |
| CaKR1/pepper | Impact on defense machinery | Expression in excess | Improved salt tolerance as well as oxidative stress | [80] |
| ZAT12/B. carinata | Transcription of C2H2 zinc finger protein | Upregulation | Enhanced tolerance to heat | [81] |
| PtADC/P. trifoliata | Involved in PAs synthesis | Upregulation | Increased tolerance to water stress | [82] |
| LeFAD3/tomato | Transcription of fatty acid and lipids unsaturation | Transcribed in excess | Augmented tolerance to salt stress | [83] |
| TaNHX2/wheat | Transport of ions; equal-ion management | Upregulation | Boosted resistance to salt stress | [84] |
| LeHSP21.5/tomato | Heat shock protein | Overexpression | Combat with tunicamycin-induced stress | [85] |
| MdPIP1; 3/Apple | Aquaporin protein and used to increase fruit size | Expression | Increased drought tolerance | [86] |
| OSMOTIN gene/N. Tobacum | Has a higher level of proline | Constitutively expression | Tolerance to salt stress | [87] |
| CodA/tomato | Organic osmolyte | Expression | Enhanced chilling stress | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rai, G.K.; Kumar, P.; Choudhary, S.M.; Kosser, R.; Khanday, D.M.; Choudhary, S.; Kumar, B.; Magotra, I.; Kumar, R.R.; Ram, C.; et al. Biomimetic Strategies for Developing Abiotic Stress-Tolerant Tomato Cultivars: An Overview. Plants 2023, 12, 86. https://doi.org/10.3390/plants12010086
Rai GK, Kumar P, Choudhary SM, Kosser R, Khanday DM, Choudhary S, Kumar B, Magotra I, Kumar RR, Ram C, et al. Biomimetic Strategies for Developing Abiotic Stress-Tolerant Tomato Cultivars: An Overview. Plants. 2023; 12(1):86. https://doi.org/10.3390/plants12010086
Chicago/Turabian StyleRai, Gyanendra Kumar, Pradeep Kumar, Sadiya Maryam Choudhary, Rafia Kosser, Danish Mushtaq Khanday, Shallu Choudhary, Bupesh Kumar, Isha Magotra, Ranjit Ranjan Kumar, Chet Ram, and et al. 2023. "Biomimetic Strategies for Developing Abiotic Stress-Tolerant Tomato Cultivars: An Overview" Plants 12, no. 1: 86. https://doi.org/10.3390/plants12010086