
Biochemical Insights into the Ability of Lemna minor L. Extract to Counteract Copper Toxicity in Maize

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
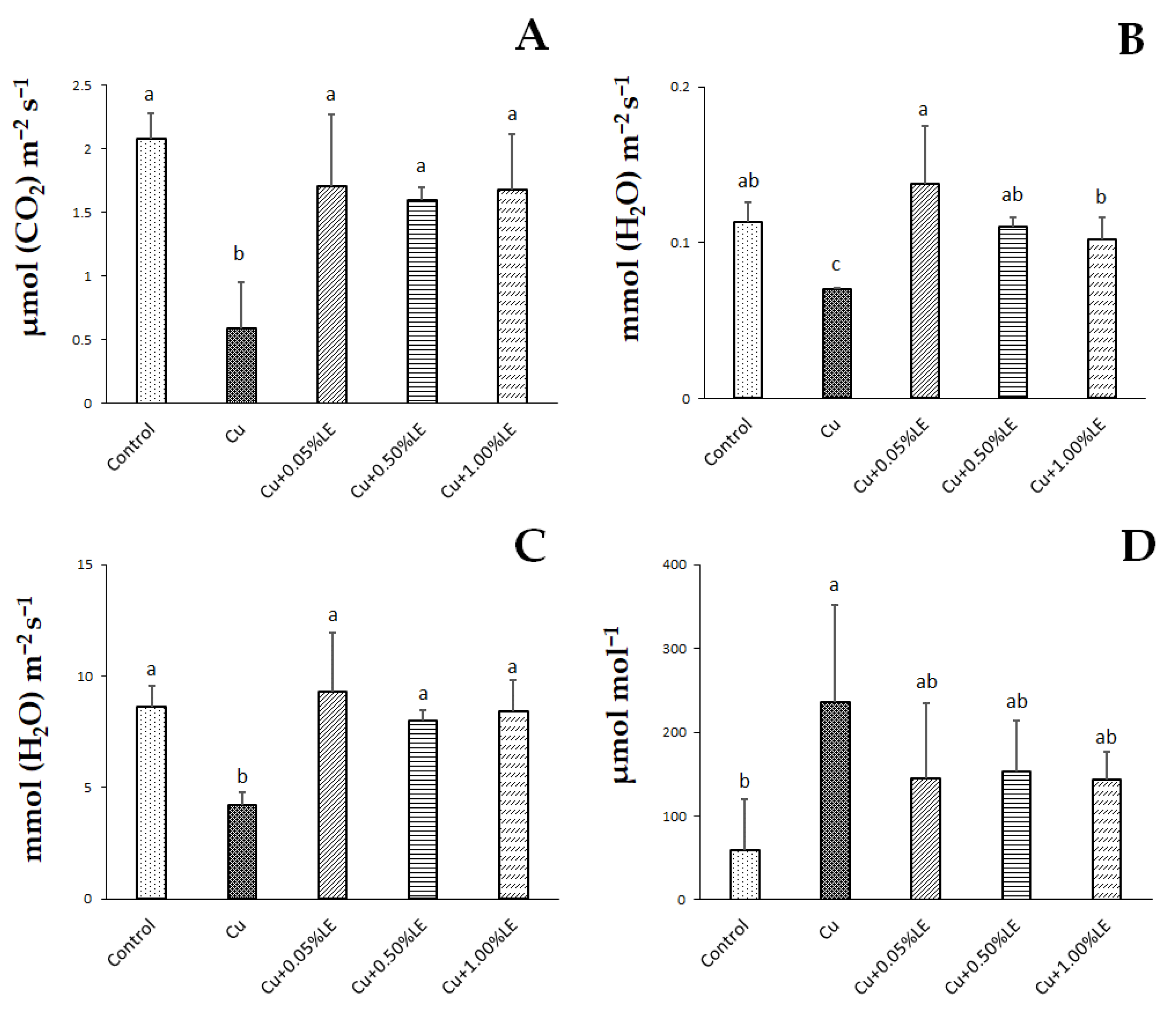
2.1. Leaves Gas Exchanges and Plant Biomass
2.2. Copper (Cu), Hydrogen Peroxide (H2O2) and Malondialdehyde (MDA) Contents in Maize Leaves
2.3. Activity of the Antioxidant Enzymes SOD, APX and CAT
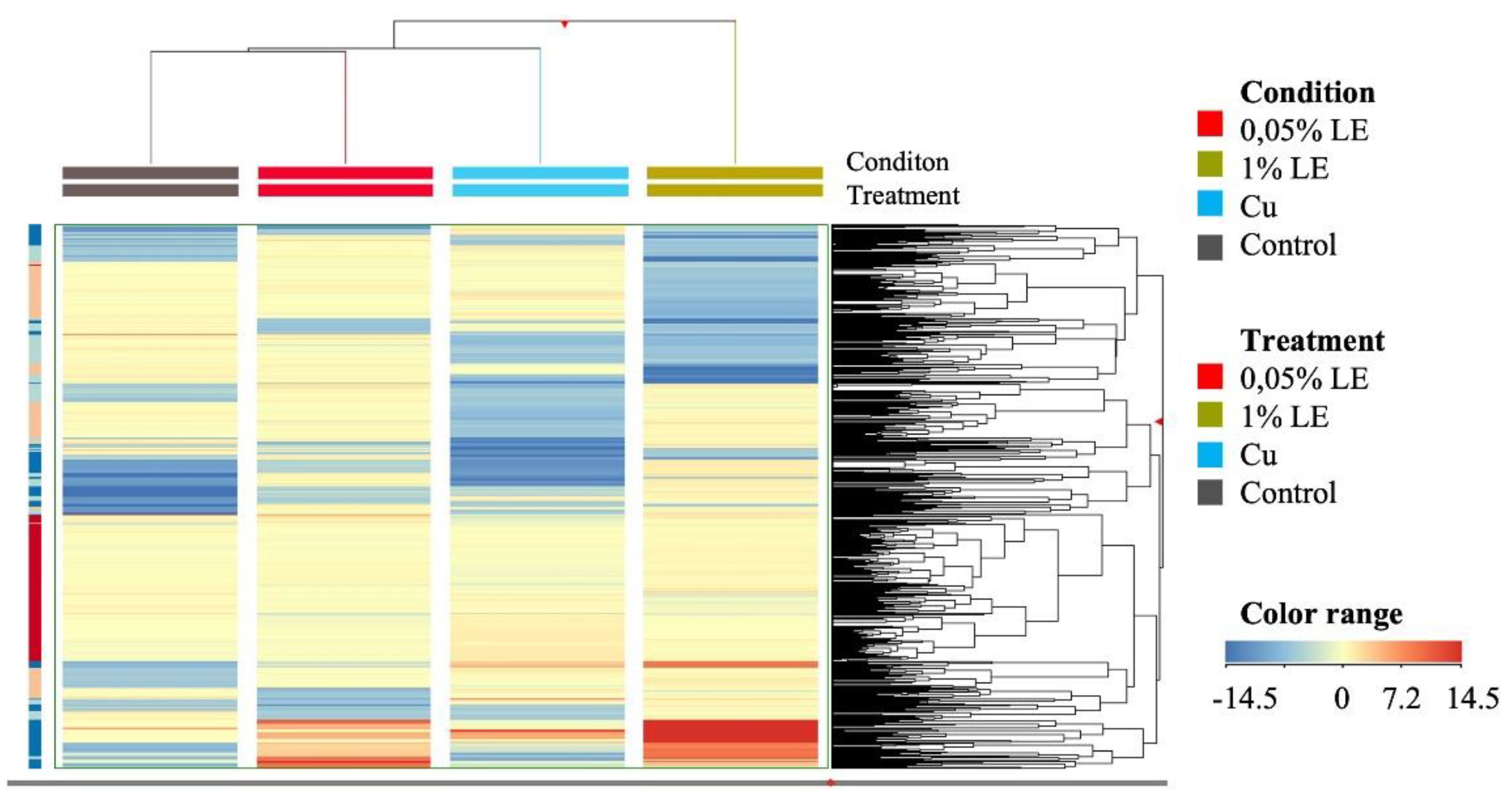
2.4. Untargeted Metabolomic Profile of Maize Samples
3. Discussion
4. Materials and Methods
4.1. Duckweed Growth Conditions and Extract Preparation
4.2. Maize Growth Conditions and Seeds Priming
4.3. Pn, gs, Evapotranspiration, Ci, and Plant Biomass
4.4. Determinations of Cu, H2O2 and MDA Contents
4.5. Activity of the Antioxidant Enzymes SOD, APX and CAT
4.6. Untargeted Metabolomics
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sall, M.L.; Diaw, A.K.D.; Gningue-Sall, D.; Efremova Aaron, S.; Aaron, J.-J. Toxic Heavy Metals: Impact on the Environment and Human Health, and Treatment with Conducting Organic Polymers, a Review. Environ. Sci. Pollut. Res. 2020, 27, 29927–29942. [Google Scholar] [CrossRef] [PubMed]
- Panfili, I.; Bartucca, M.L.; Del Buono, D. The Treatment of Duckweed with a Plant Biostimulant or a Safener Improves the Plant Capacity to Clean Water Polluted by Terbuthylazine. Sci. Total Environ. 2019, 646, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Adrees, M.; Ali, S.; Rizwan, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Zia-ur-Rehman, M.; Irshad, M.K.; Bharwana, S.A. The Effect of Excess Copper on Growth and Physiology of Important Food Crops: A Review. Environ. Sci. Pollut. Res. 2015, 22, 8148–8162. [Google Scholar] [CrossRef] [PubMed]
- Matayoshi, C.L.; Pena, L.B.; Arbona, V.; Gómez-Cadenas, A.; Gallego, S.M. Early Responses of Maize Seedlings to Cu Stress Include Sharp Decreases in Gibberellins and Jasmonates in the Root Apex. Protoplasma 2020, 257, 1243–1256. [Google Scholar] [CrossRef]
- Panfili, I.; Bartucca, M.L.; Ballerini, E.; Del Buono, D. Combination of Aquatic Species and Safeners Improves the Remediation of Copper Polluted Water. Sci. Total Environ. 2017, 601–602, 1263–1270. [Google Scholar] [CrossRef]
- Yruela, I.; Pueyo, J.J.; Alonso, P.J.; Picorel, R. Photoinhibition of Photosystem II from Higher Plants: Effect of Copper Inhibition. J. Biol. Chem. 1996, 271, 27408–27415. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Rea, E.; Colla, G. Grafting of Cucumber as a Means to Minimize Copper Toxicity. Environ. Exp. Bot. 2008, 63, 49–58. [Google Scholar] [CrossRef]
- Cambrollé, J.; García, J.L.; Ocete, R.; Figueroa, M.E.; Cantos, M. Growth and Photosynthetic Responses to Copper in Wild Grapevine. Chemosphere 2013, 93, 294–301. [Google Scholar] [CrossRef]
- Tanyolaç, D.; Ekmekçi, Y.; Ünalan, Ş. Changes in Photochemical and Antioxidant Enzyme Activities in Maize (Zea mays L.) Leaves Exposed to Excess Copper. Chemosphere 2007, 67, 89–98. [Google Scholar] [CrossRef]
- Tedeschini, E.; Proietti, P.; Timorato, V.; D’Amato, R.; Nasini, L.; Dei Buono, D.; Businelli, D.; Frenguelli, G. Selenium as Stressor and Antioxidant Affects Pollen Performance in Olea Europaea. Flora Morphol. Distrib. Funct. Ecol. Plants 2015, 215, 16–22. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Del Buono, D. Can Biostimulants Be Used to Mitigate the Effect of Anthropogenic Climate Change on Agriculture? It Is Time to Respond. Sci. Total Environ. 2021, 751, 141763. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Alharby, H.F.; Al-Zahrani, H.S.; Hakeem, K.R.; Alsamadany, H.; Desoky, E.-S.M.; Rady, M.M. Silymarin-Enriched Biostimulant Foliar Application Minimizes the Toxicity of Cadmium in Maize by Suppressing Oxidative Stress and Elevating Antioxidant Gene Expression. Biomolecules 2021, 11, 465. [Google Scholar] [CrossRef]
- Canellas, L.P.; Canellas, N.O.A.; da S. Irineu, L.E.S.; Olivares, F.L.; Piccolo, A. Plant Chemical Priming by Humic Acids. Chem. Biol. Technol. Agric. 2020, 7, 12. [Google Scholar] [CrossRef]
- Sherwood, J. The Significance of Biomass in a Circular Economy. Bioresour. Technol. 2020, 300, 122755. [Google Scholar] [CrossRef]
- Del Buono, D.; Bartucca, M.L.; Ballerini, E.; Senizza, B.; Lucini, L.; Trevisan, M. Physiological and Biochemical Effects of an Aqueous Extract of Lemna minor L. as a Potential Biostimulant for Maize. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Regni, L.; Del Buono, D.; Miras-Moreno, B.; Senizza, B.; Lucini, L.; Trevisan, M.; Morelli Venturi, D.; Costantino, F.; Proietti, P. Biostimulant Effects of an Aqueous Extract of Duckweed (Lemna minor L.) on Physiological and Biochemical Traits in the Olive Tree. Agriculture 2021, 11, 1299. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef] [PubMed]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Lopes, J.I.; Gonçalves, B.C.; Ferreira, T.C.; Correia, C.M. Physiological Behaviour, Oxidative Damage and Antioxidative Protection of Olive Trees Grown under Different Irrigation Regimes. Plant Soil 2007, 292, 1–12. [Google Scholar] [CrossRef]
- Proietti, P.; Nasini, L.; Del Buono, D.; D’Amato, R.; Tedeschini, E.; Businelli, D. Selenium Protects Olive (Olea europaea L.) from Drought Stress. Sci. Hortic. 2013, 164, 165–171. [Google Scholar] [CrossRef]
- Singh, S.K.; Raja Reddy, K. Regulation of Photosynthesis, Fluorescence, Stomatal Conductance and Water-Use Efficiency of Cowpea (Vigna unguiculata [L.] Walp.) under Drought. J. Photochem. Photobiol. B 2011, 105, 40–50. [Google Scholar] [CrossRef]
- Pätsikkä, E.; Kairavuo, M.; Šeršen, F.; Aro, E.-M.; Tyystjärvi, E. Excess Copper Predisposes Photosystem II to Photoinhibition in Vivo by Outcompeting Iron and Causing Decrease in Leaf Chlorophyll. Plant Physiol. 2002, 129, 1359–1367. [Google Scholar] [CrossRef] [Green Version]
- Vinit-Dunand, F.; Epron, D.; Alaoui-Sossé, B.; Badot, P.-M. Effects of Copper on Growth and on Photosynthesis of Mature and Expanding Leaves in Cucumber Plants. Plant Sci. 2002, 163, 53–58. [Google Scholar] [CrossRef]
- Wodala, B.; Eitel, G.; Gyula, T.N.; Ördög, A.; Horváth, F. Monitoring Moderate Cu and Cd Toxicity by Chlorophyll Fluorescence and P 700 Absorbance in Pea Leaves. Photosynthetica 2012, 50, 380–386. [Google Scholar] [CrossRef]
- Vassilev, A.; Lidon, F.C.; Matos, M.D.C.; Ramalho, J.C.; Yordanov, I. Photosynthetic Performance and Content of Some Nutrients in Cadmium- and Copper-Treated Barley Plants. J. Plant Nutr. 2002, 25, 2343–2360. [Google Scholar] [CrossRef]
- Del Buono, D.; Regni, L.; Del Pino, A.M.; Bartucca, M.L.; Palmerini, C.A.; Proietti, P. Effects of Megafol on the Olive Cultivar ‘Arbequina’ Grown Under Severe Saline Stress in Terms of Physiological Traits, Oxidative Stress, Antioxidant Defenses, and Cytosolic Ca2+. Front. Plant Sci. 2021, 11, 603576. [Google Scholar] [CrossRef]
- Almadi, L.; Paoletti, A.; Cinosi, N.; Daher, E.; Rosati, A.; Di Vaio, C.; Famiani, F. A Biostimulant Based on Protein Hydrolysates Promotes the Growth of Young Olive Trees. Agriculture 2020, 10, 618. [Google Scholar] [CrossRef]
- Kałuzewicz, A.; Krzesiński, W.; Spizewski, T.; Zaworska, A. Effect of Biostimulants on Several Physiological Characteristics and Chlorophyll Content in Broccoli under Drought Stress and Re-Watering. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Fascella, G.; Montoneri, E.; Ginepro, M.; Francavilla, M. Effect of Urban Biowaste Derived Soluble Substances on Growth, Photosynthesis and Ornamental Value of Euphorbia × Lomi. Sci. Hortic. 2015, 197, 90–98. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant Application with a Tropical Plant Extract Enhances Corchorus Olitorius Adaptation to Sub-Optimal Nutrient Regimens by Improving Physiological Parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Morgutti, S.; Cocetta, G.; Negrini, N.; Farris, S.; Calcante, A.; Spinardi, A.; Ferrari, E.; Mignani, I.; Oberti, R.; et al. Evaluation of Borage Extracts as Potential Biostimulant Using a Phenomic, Agronomic, Physiological, and Biochemical Approach. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A Novel Type of Seaweed Extract as a Natural Alternative to the Use of Iron Chelates in Strawberry Production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Sousa, F.; Martins, M.; Sousa, B.; Soares, C.; Azenha, M.; Pereira, R.; Fidalgo, F. Wrack Composed by Fucus Spp, Ascophyllum Nodosum and Pelvetia Canaliculata Limits Metal Uptake and Restores the Redox Homeostasis of Barley Plants Grown in Cu-Contaminated Soils. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Lee, M.H.; Cho, E.J.; Wi, S.G.; Bae, H.; Kim, J.E.; Cho, J.-Y.; Lee, S.; Kim, J.-H.; Chung, B.Y. Divergences in Morphological Changes and Antioxidant Responses in Salt-Tolerant and Salt-Sensitive Rice Seedlings after Salt Stress. Plant Physiol. Biochem. 2013, 70, 325–335. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Merwad, A.-R.M.; Rady, M.M. Natural Biostimulants Improve Saline Soil Characteristics and Salt Stressed-Sorghum Performance. Commun. Soil Sci. Plant Anal. 2018, 49, 967–983. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant Salt-Tolerance Mechanism: A Review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Willekens, H.; Langebartels, C.; Tiré, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Differential Expression of Catalase Genes in Nicotiana plumbaginifolia (L.). Proc. Natl. Acad. Sci. USA 1994, 91, 10450–10454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, J.; Lee, S.; Wen, R. Copper-Caused Oxidative Stress Triggers the Activation of Antioxidant Enzymes via ZmMPK3 in Maize Leaves. PLoS ONE 2018, 13, e0203612. [Google Scholar] [CrossRef] [Green Version]
- Luna, C.M.; González, C.A.; Trippi, V.S. Oxidative Damage Caused by an Excess of Copper in Oat Leaves. Plant Cell Physiol. 1994, 35, 11–15. [Google Scholar]
- Lombardi, L.; Sebastiani, L. Copper Toxicity in Prunus Cerasifera: Growth and Antioxidant Enzymes Responses of in Vitro Grown Plants. Plant Sci. 2005, 168, 797–802. [Google Scholar] [CrossRef]
- Rama Devi, S.; Prasad, M.N.V. Copper Toxicity in Ceratophyllum demersum L. (Coontail), a Floating Macrophyte: Response of Antioxidant Enzymes and Antioxidants. Plant Sci. 1998, 138, 157–165. [Google Scholar] [CrossRef]
- Elansary, H.O.; Mahmoud, E.A.; El-Ansary, D.O.; Mattar, M.A. Effects of Water Stress and Modern Biostimulants on Growth and Quality Characteristics of Mint. Agronomy 2020, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Borhannuddin Bhuyan, M.H.M.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Miras-Moreno, B.; Yildiztugay, E.; Ozfidan-Konakci, C.; Arikan, B.; Elbasan, F.; Ak, G.; Rouphael, Y.; Zengin, G.; Lucini, L. Metabolomics and Physiological Insights into the Ability of Exogenously Applied Chlorogenic Acid and Hesperidin to Modulate Salt Stress in Lettuce Distinctively. Molecules 2021, 26, 6291. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Yusuf, M.; Ahmad, I.; Ahmad, A. Brassinosteroids and Their Role in Response of Plants to Abiotic Stresses. Biol. Plant. 2014, 58, 9–17. [Google Scholar] [CrossRef]
- Carrió-Seguí, À.; Romero, P.; Sanz, A.; Peñarrubia, L. Interaction between ABA Signaling and Copper Homeostasis in Arabidopsis Thaliana. Plant Cell Physiol. 2016, 57, 1568–1582. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed Priming with Phytohormones: An Effective Approach for the Mitigation of Abiotic Stress. Plants 2021, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.H.; De Bang, T.C.; Laursen, K.H.; Pedas, P.; Husted, S.; Schjoerring, J.K. Multielement Plant Tissue Analysis Using ICP Spectrometry. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; Volume 953, p. 141. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative Stress and Some Antioxidant Systems in Acid Rain-Treated Bean Plants: Protective Role of Exogenous Polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for Superoxide Dismutase Activity: Some Large Consequences of Minor Changes in Conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Aebi, H. Catalase in Vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. ISBN 0076-6879. [Google Scholar]
- Borgognone, D.; Rouphael, Y.; Cardarelli, M.; Lucini, L.; Colla, G. Changes in Biomass, Mineral Composition, and Quality of Cardoon in Response to NO3−:Cl− Ratio and Nitrate Deprivation from the Nutrient Solution. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.; Ginzburg, D.; Zhao, K.; Dwyer, W.; Xue, B.; Xu, A.; Rice, S.; Cole, B.; Paley, S.; Karp, P.; et al. Plant Metabolic Network 15: A Resource of Genome-Wide Metabolism Databases for 126 Plants and Algae. J. Integr. Plant Biol. 2021, 63, 1888–1905. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The Role of Reporting Standards for Metabolite Annotation and Identification in Metabolomic Studies. GigaScience 2013, 2. [Google Scholar] [CrossRef]
- Caspi, R.; Dreher, K.; Karp, P.D. The Challenge of Constructing, Classifying, and Representing Metabolic Pathways. FEMS Microbiol. Lett. 2013, 345, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karp, P.D.; Paley, S.M.; Krummenacker, M.; Latendresse, M.; Dale, J.M.; Lee, T.J.; Kaipa, P.; Gilham, F.; Spaulding, A.; Popescu, L.; et al. Pathway Tools Version 13.0: Integrated Software for Pathway/Genome Informatics and Systems Biology. Brief Bioinform. 2009, 11, 40–79. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shoot Length (cm) | Roots Length (cm) | Shoot FW (g plant−1) | Roots FW (g plant−1) | |
|---|---|---|---|---|
| Control | 23.2 (0.8) a | 20.7 (1.7) ab | 0.98 (0.24) b | 0.200 (0.022) b |
| Cu | 14.5 (0.9) b | 8.3 (1.6) c | 0.58 (0.09) c | 0.129 (0.020) c |
| Cu + 0.05% LE | 22.7 (1.4) a | 18.8 (2.5) b | 1.21 (0.26) ab | 0.256 (0.045) ab |
| Cu + 0.50% LE | 23.9 (1.8) a | 23.2 (3.0) a | 1.35 (0.15) a | 0.289 (0.049) a |
| Cu +1.00% LE | 22.6 (1.52) a | 18.7 (1.0) b | 0.83 (0.10) bc | 0.264 (0.027) ab |
| Cu (µg g−1 DW) | H2O2 (nmol g−1 FW) | MDA (nmol g−1 FW) | |
|---|---|---|---|
| Control | 18.4 (1.2) b | 116.1 (5.0) d | 21.6 (2.8) c |
| Cu | 38.8 (7.1) a | 211.6 (15.0) a | 41.6 (4.3) a |
| Cu + 0.05% LE | 35.1 (3.7) a | 156.3 (19.4) bc | 19.6 (1.5) c |
| Cu + 0.50% LE | 36.3 (6.0) a | 135.4 (20.4) cd | 21.1 (3.7) c |
| Cu +1.00% LE | 32.2 (4.5) a | 179.6 (11.8) b | 30.6 (3.1) b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miras-Moreno, B.; Senizza, B.; Regni, L.; Tolisano, C.; Proietti, P.; Trevisan, M.; Lucini, L.; Rouphael, Y.; Del Buono, D. Biochemical Insights into the Ability of Lemna minor L. Extract to Counteract Copper Toxicity in Maize. Plants 2022, 11, 2613. https://doi.org/10.3390/plants11192613
Miras-Moreno B, Senizza B, Regni L, Tolisano C, Proietti P, Trevisan M, Lucini L, Rouphael Y, Del Buono D. Biochemical Insights into the Ability of Lemna minor L. Extract to Counteract Copper Toxicity in Maize. Plants. 2022; 11(19):2613. https://doi.org/10.3390/plants11192613
Chicago/Turabian StyleMiras-Moreno, Begoña, Biancamaria Senizza, Luca Regni, Ciro Tolisano, Primo Proietti, Marco Trevisan, Luigi Lucini, Youssef Rouphael, and Daniele Del Buono. 2022. "Biochemical Insights into the Ability of Lemna minor L. Extract to Counteract Copper Toxicity in Maize" Plants 11, no. 19: 2613. https://doi.org/10.3390/plants11192613