
ITS Metabarcoding Reveals the Effects of Oregano Essential Oil on Fusarium oxysporum and Other Fungal Species in Soil Samples
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. GC Analysis
2.2. Sequencing
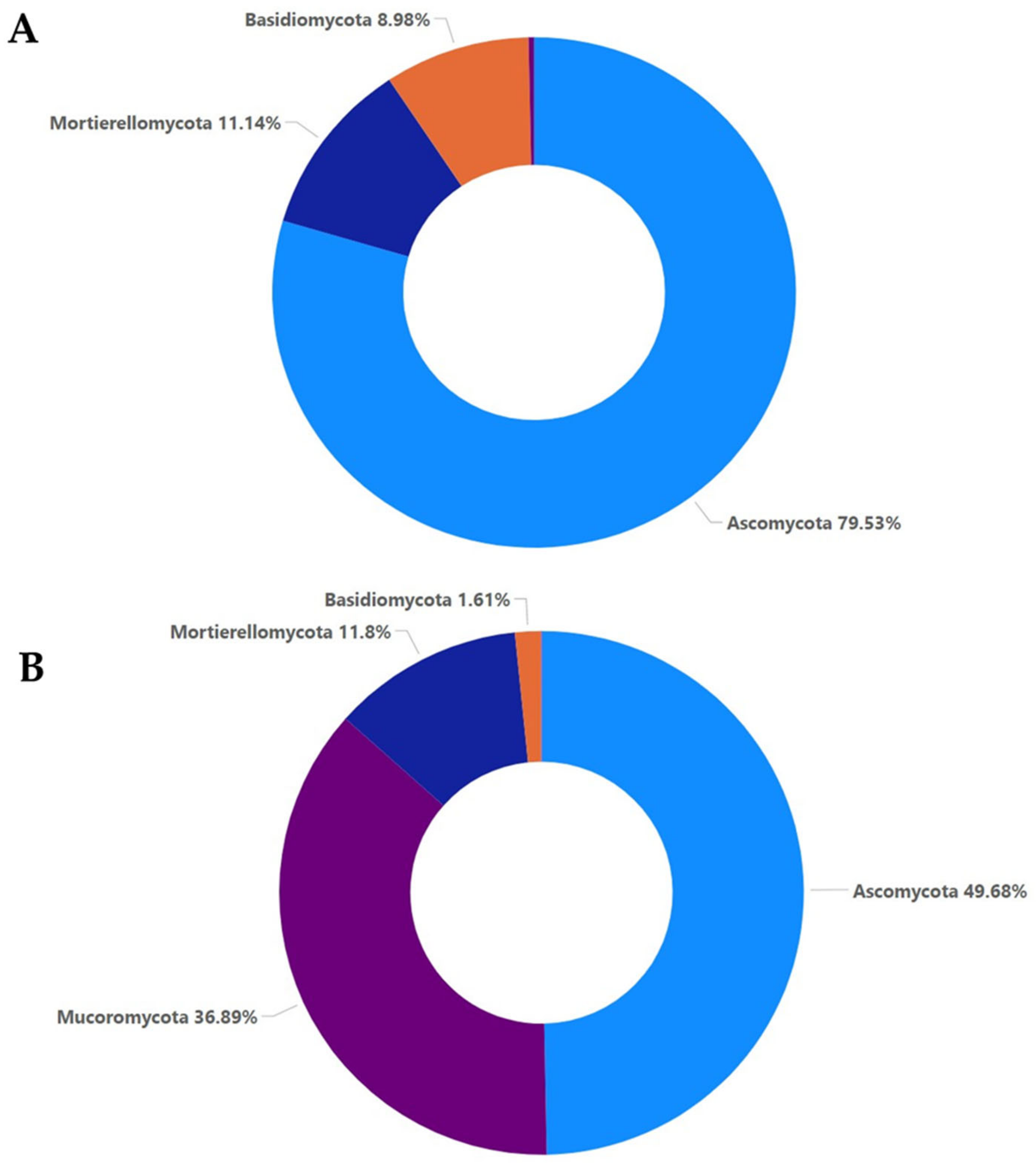
2.3. Relative Abundance of Soil Fungal Community per Treatment
2.4. Relative Abundance (Family and Genus) per Sample for Each Treatment
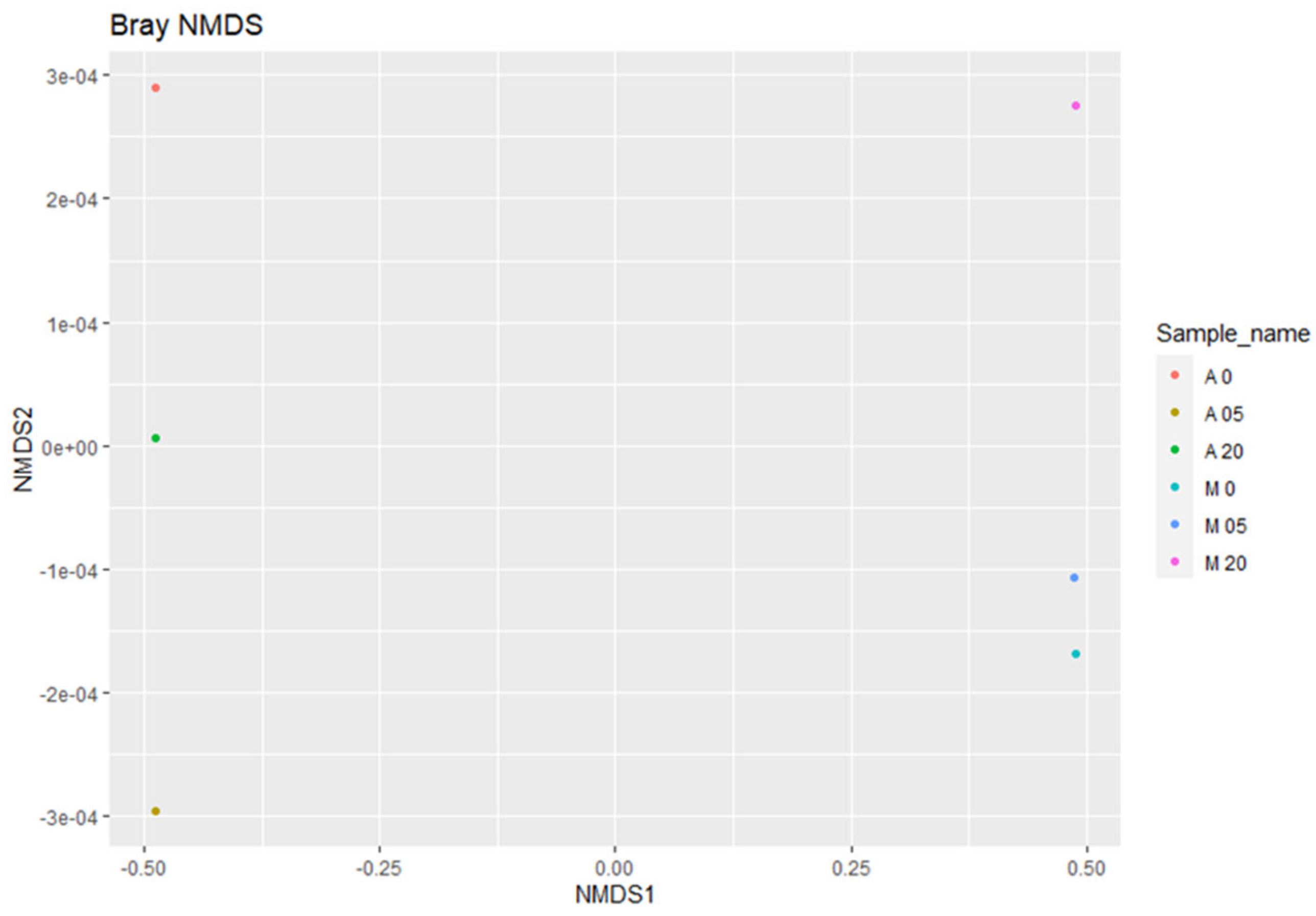
2.5. Species Diversity and Effect of Oregano Essential Oil
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aoki, T.; O’Donnell, K.; Geiser, D.M. Systematics of Key Phytopathogenic Fusarium Species: Current Status and Future Challenges. J. Gen. Plant Pathol. 2014, 80, 189–201. [Google Scholar] [CrossRef]
- Weller, D.M.; Raaijmakers, J.M.; Gardener, B.B.M.; Thomashow, L.S. Microbial Populations Responsible for Specific Soil Suppressiveness to Plant Pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isman, M.B. Plant Essential Oils for Pest and Disease Management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Burt, S. Essential Oils: Their Antibacterial Properties and Potential Applications in Foods—A Review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Grayer, R.J.; Harborne, J.B. A Survey of Antifungal Compounds from Higher Plants, 1982–1993. Phytochemistry 1994, 37, 19–42. [Google Scholar] [CrossRef]
- Singh, P.; Pandey, A.K.; Tripathi, N.N. Essential Oils: A Renewable Source for The Management of Stored Product Insects—A Review. Agri. Rev. 2012, 33, 226–236. [Google Scholar]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Oka, Y.; Nacar, S.; Putievsky, E.; Ravid, U.; Yaniv, Z.; Spiegel, Y. Nematicidal Activity of Essential Oils and Their Components Against the Root-Knot Nematode. Phytopathology 2000, 90, 710–715. [Google Scholar] [CrossRef] [Green Version]
- Soković, M.; Van Griensven, L.J.L.D. Antimicrobial Activity of Essential Oils and Their Components against the Three Major Pathogens of the Cultivated Button Mushroom, Agaricus Bisporus. Eur. J. Plant Pathol. 2006, 116, 211–224. [Google Scholar] [CrossRef]
- Nabigol, A.; Farzaneh, M. In Vitro Antifungal Activity of Some Plant Essential Oils on Postharvest Pathogens of Strawberry Fruit. Acta Hortic. 2010, 858, 305–311. [Google Scholar] [CrossRef]
- Daferera, D.J.; Ziogas, B.N.; Polissiou, M.G. The Effectiveness of Plant Essential Oils on the Growth of Botrytis Cinerea, Fusarium Sp. and Clavibacter Michiganensis Subsp. Michiganensis. Crop Prot. 2003, 22, 39–44. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and Antifungal Properties of Essential Oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.; Ademiluyi, A.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-González, G.F.; Phumthum, M. Fundamental Chemistry of Essential Oils and Volatile Organic Compounds, Methods of Analysis and Authentication. Plants 2022, 11, 789. [Google Scholar] [CrossRef]
- Tančinová, D.; Mašková, Z.; Denisa Foltinová, D.; Štefániková, J.; Árvay, J. Effect of Essential Oils of Lamiaceae Plants on the Rhizopus Spp. Potravin. Slovak J. Food Sci. 2018, 12, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Tsao, R.; Zhou, T. Antifungal Activity of Monoterpenoids against Postharvest Pathogens Botrytis Cinerea and Monilinia Fructicola. J. Essent. Oil Res. 2000, 12, 113–121. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, S.; Du, S.; Chen, S.; Sun, H. Antifungal Activity of Thymol and Carvacrol against Postharvest Pathogens Botrytis Cinerea. J. Food Sci. Technol. 2019, 56, 2611–2620. [Google Scholar] [CrossRef]
- Wogiatzi, E.; Gougoulias, N.; Papachatzis, A.; Vagelas, I.; Chouliaras, N. Greek Oregano Essential Oils Production, Phytotoxicity and Antifungal Activity. Biotechnol. Biotechnol. Equip. 2009, 23, 1150–1152. [Google Scholar] [CrossRef] [Green Version]
- Vokou, D.; Liotiri, S. Stimulation of Soil Microbial Activity by Essential Oils. Chemoecology 1999, 9, 41–45. [Google Scholar] [CrossRef]
- Buée, M.; De Boer, W.; Martin, F.; Van Overbeek, L.; Jurkevitch, E. The Rhizosphere Zoo: An Overview of Plant-Associated Communities of Microorganisms, Including Phages, Bacteria, Archaea, and Fungi, and of Some of Their Structuring Factors. Plant Soil 2009, 321, 189–212. [Google Scholar] [CrossRef]
- Yahr, R.; Schoch, C.L.; Dentinger, B.T.M. Scaling up Discovery of Hidden Diversity in Fungi: Impacts of Barcoding Approaches. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150336. [Google Scholar] [CrossRef] [PubMed]
- Piombo, E.; Abdelfattah, A.; Droby, S.; Wisniewski, M.; Spadaro, D.; Schena, L. Metagenomics Approaches for the Detection and Surveillance of Emerging and Recurrent Plant Pathogens. Microorganisms 2021, 9, 188. [Google Scholar] [CrossRef] [PubMed]
- Cristescu, M.E. From Barcoding Single Individuals to Metabarcoding Biological Communities: Towards an Integrative Approach to the Study of Global Biodiversity. Trends Ecol. Evol. 2014, 29, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal Community Analysis by High-throughput Sequencing of Amplified Markers—A User’s Guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarenkov, K.; Henrik Nilsson, R.; Larsson, K.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE Database for Molecular Identification of Fungi—Recent Updates and Future Perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Gao, M.; Xiong, C.; Gao, C.; Tsui, C.K.M.; Wang, M.-M.; Zhou, X.; Zhang, A.-M.; Cai, L. Disease-Induced Changes in Plant Microbiome Assembly and Functional Adaptation. Microbiome 2021, 9, 187. [Google Scholar] [CrossRef]
- Jiang, G.; Zhang, Y.; Gan, G.; Li, W.; Wan, W.; Jiang, Y.; Yang, T.; Zhang, Y.; Xu, Y.; Wang, Y.; et al. Exploring Rhizo-Microbiome Transplants as a Tool for Protective Plant-Microbiome Manipulation. ISME Commun. 2022, 2, 10. [Google Scholar] [CrossRef]
- Suman, J.; Rakshit, A.; Ogireddy, S.D.; Singh, S.; Gupta, C.; Chandrakala, J. Microbiome as a Key Player in Sustainable Agriculture and Human Health. Front. Soil Sci. 2022, 2, 821589. [Google Scholar] [CrossRef]
- Hedlund, J.; Longo, S.B.; York, R. Agriculture, Pesticide Use, and Economic Development: A Global Examination (1990–2014). Rural Sociol. 2020, 85, 519–544. [Google Scholar] [CrossRef]
- Araniti, F.; Graña, E.; Reigosa, M.J.; Sánchez-Moreiras, A.M.; Abenavoli, M.R. Individual and Joint Activity of Terpenoids, Isolated from Calamintha Nepeta Extract, on Arabidopsis Thaliana. Nat. Prod. Res. 2013, 27, 2297–2303. [Google Scholar] [CrossRef]
- Araniti, F.; Landi, M.; Laudicina, V.A.; Abenavoli, M.R. Secondary Metabolites and Eco-Friendly Techniques for Agricultural Weed/Pest Management. Plants 2021, 10, 1418. [Google Scholar] [CrossRef]
- López-González, D.; Costas-Gil, A.; Reigosa, M.J.; Araniti, F.; Sánchez-Moreiras, A.M. A Natural Indole Alkaloid, Norharmane, Affects PIN Expression Patterns and Compromises Root Growth in Arabidopsis Thaliana. Plant Physiol. Biochem. 2020, 151, 378–390. [Google Scholar] [CrossRef]
- Amini, J.; Farhang, V.; Javadi, T.; Nazemi, J. Antifungal Effect of Plant Essential Oils on Controlling Phytophthora Species. Plant Pathol. J. 2016, 32, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Bång, U. Screening of Natural Plant Volatiles to Control the Potato (Solanum Tuberosum) Pathogens Helminthosporium Solani, Fusarium Solani, Phoma Foveata and Rhizoctonia Solani. Potato Res. 2007, 50, 185–203. [Google Scholar] [CrossRef]
- Cavanagh, H.M.A. Antifungal Activity of the Volatile Phase of Essential Oils: A Brief Review. Nat. Prod. Commun. 2007, 2, 1934578X0700201. [Google Scholar] [CrossRef] [Green Version]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The Genomics of Opportunistic Success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Friedl, M.A.; Druzhinina, I.S. Taxon-Specific Metagenomics of Trichoderma Reveals a Narrow Community of Opportunistic Species That Regulate Each Other’s Development. Microbiology 2012, 158, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Akhtar, N.; Msimbira, L.A.; Antar, M.; Ashraf, S.; Khan, S.N.; Smith, D.L. Neocosmospora Rubicola, a Stem Rot Disease in Potato: Characterization, Distribution and Management. Front. Microbiol. 2022, 13, 953097. [Google Scholar] [CrossRef]
- Ziaee, A.; Zia, M.; Bayat, M.; Hashemi, J. Identification of Mucorales Isolates from Soil Using Morphological and Molecular Methods. Curr. Med. Mycol. 2016, 2, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Attia, M.S.; El-Wakil, D.A.; Hashem, A.H.; Abdelaziz, A.M. Antagonistic Effect of Plant Growth-Promoting Fungi Against Fusarium Wilt Disease in Tomato: In Vitro and In Vivo Study. Appl. Biochem. Biotechnol. 2022, 194, 5100–5118. [Google Scholar] [CrossRef] [PubMed]
- Durán, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 2018, 175, 973–983.e14. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, M.G.; De Bruin, S.; Luckerhoff, L.; Van Logtestijn, R.S.; Schlaeppi, K. A Widespread Plant-Fungal-Bacterial Symbiosis Promotes Plant Biodiversity, Plant Nutrition and Seedling Recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using Network Analysis to Explore Co-Occurrence Patterns in Soil Microbial Communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Wang, H.; Dsouza, M.; Lou, J.; He, Y.; Dai, Z.; Brookes, P.C.; Xu, J.; Gilbert, J.A. Geographic Patterns of Co-Occurrence Network Topological Features for Soil Microbiota at Continental Scale in Eastern China. ISME J. 2016, 10, 1891–1901. [Google Scholar] [CrossRef]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The Interconnected Rhizosphere: High Network Complexity Dominates Rhizosphere Assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [Green Version]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; De Hollander, M.; Soto, R.L.; Bouffaud, M.-L.; Buée, M.; Dimmers, W.; et al. Soil Networks Become More Connected and Take up More Carbon as Nature Restoration Progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef] [Green Version]
- De Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil Bacterial Networks Are Less Stable under Drought than Fungal Networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; Van der Heijden, M.G.A. Agricultural Intensification Reduces Microbial Network Complexity and the Abundance of Keystone Taxa in Roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [Green Version]
- Paster, N.; Juven, B.J.; Shaaya, E.; Menasherov, M.; Nitzan, R.; Weisslowicz, H.; Ravid, U. Inhibitory Effect of Oregano and Thyme Essential Oils on Moulds and Foodborne Bacteria. Lett. Appl. Microbiol. 1990, 11, 33–37. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Suo, M.; Qiu, Z.; Wu, H.; Zhao, M.; Yang, H. Regulating Root Fungal Community Using Mortierella Alpina for Fusarium Oxysporum Resistance in Panax Ginseng. Front. Microbiol. 2022, 13, 850917. [Google Scholar] [CrossRef]
- Kishore, G.K.; Pande, S.; Harish, S. Evaluation of Essential Oils and Their Components for Broad-Spectrum Antifungal Activity and Control of Late Leaf Spot and Crown Rot Diseases in Peanut. Plant Dis. 2007, 91, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Tanovic, B.; Milijasevic, S.; Obradovic, A. In Vitro Effect of Plant Essential Oils on Growth of Some Soil-Borne Pathogens. Acta Hortic. 2007, 729, 467–471. [Google Scholar] [CrossRef]
- Kumar, U. Antimicrobial Activity of Essential Oils Against Plant Pathogenic Fungi: A Review. Int. J. Incl. Dev. 2020, 6, 37–44. [Google Scholar] [CrossRef]
- Evergetis, E.; Michaelakis, A.; Papachristos, D.P.; Badieritakis, E.; Kapsaski-Kanelli, V.N.; Haroutounian, S.A. Seasonal Variation and Bioactivity of the Essential Oils of Two Juniperus Species against Aedes (Stegomyia) Albopictus (Skuse, 1894). Parasitol. Res. 2016, 115, 2175–2183. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; Van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Reads before Filtering | Reads after Filtering |
|---|---|---|
| A0 | 150,275 | 149,635 |
| A5 | 122,035 | 121,498 |
| A20 | 167,589 | 166,801 |
| M0 | 137,604 | 136,998 |
| M5 | 85,774 | 85,412 |
| M20 | 103,173 | 102,805 |
| Average | 127,742 | 127,192 |
| Total | 766,450 | 763,149 |
| Region | Primer | Sequence |
|---|---|---|
| ITS1 | Forward_1 | 5′- TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGAGAGTTCATGCCCGAAAGGG-3′ |
| Reverse_1 | 5′-GTCTCGTGGGCT CGGAGATGTGTATAAGAGACAGCTGCGTTCTTCATCGAT-3′ | |
| Forward_2 | 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGGAAGGTGAAGTCGTAACAAGG-3′ | |
| Reverse_2 | 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGAGCGTTCTTCATCGATGTGC-3′ | |
| ITS2 | Forward_1 | 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGATCGATG AAGAACGCAG-3′ |
| Reverse_1 | 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGGTTTTGGCAGAAGCACACC-3′ | |
| Forward_2 | 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCGTGAAGTGTCTTGCTGGTC-3′ | |
| Reverse_2 | 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGCACCATACTTCGCGCAACAC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karapetsi, L.; Pratsinakis, E.; Xirakias, F.; Osathanunkul, M.; Vagelas, I.; Madesis, P. ITS Metabarcoding Reveals the Effects of Oregano Essential Oil on Fusarium oxysporum and Other Fungal Species in Soil Samples. Plants 2023, 12, 62. https://doi.org/10.3390/plants12010062
Karapetsi L, Pratsinakis E, Xirakias F, Osathanunkul M, Vagelas I, Madesis P. ITS Metabarcoding Reveals the Effects of Oregano Essential Oil on Fusarium oxysporum and Other Fungal Species in Soil Samples. Plants. 2023; 12(1):62. https://doi.org/10.3390/plants12010062
Chicago/Turabian StyleKarapetsi, Lefkothea, Emmanouil Pratsinakis, Fotis Xirakias, Maslin Osathanunkul, Ioannis Vagelas, and Panagiotis Madesis. 2023. "ITS Metabarcoding Reveals the Effects of Oregano Essential Oil on Fusarium oxysporum and Other Fungal Species in Soil Samples" Plants 12, no. 1: 62. https://doi.org/10.3390/plants12010062