
High Input of Nitrogen Fertilization and Short Irrigation Frequencies Forcefully Promote the Development of Verticillium Wilt of Olive

Abstract
:1. Introduction
2. Results
2.1. Disease Symptoms and Pathogen Isolation from Affected Tissues
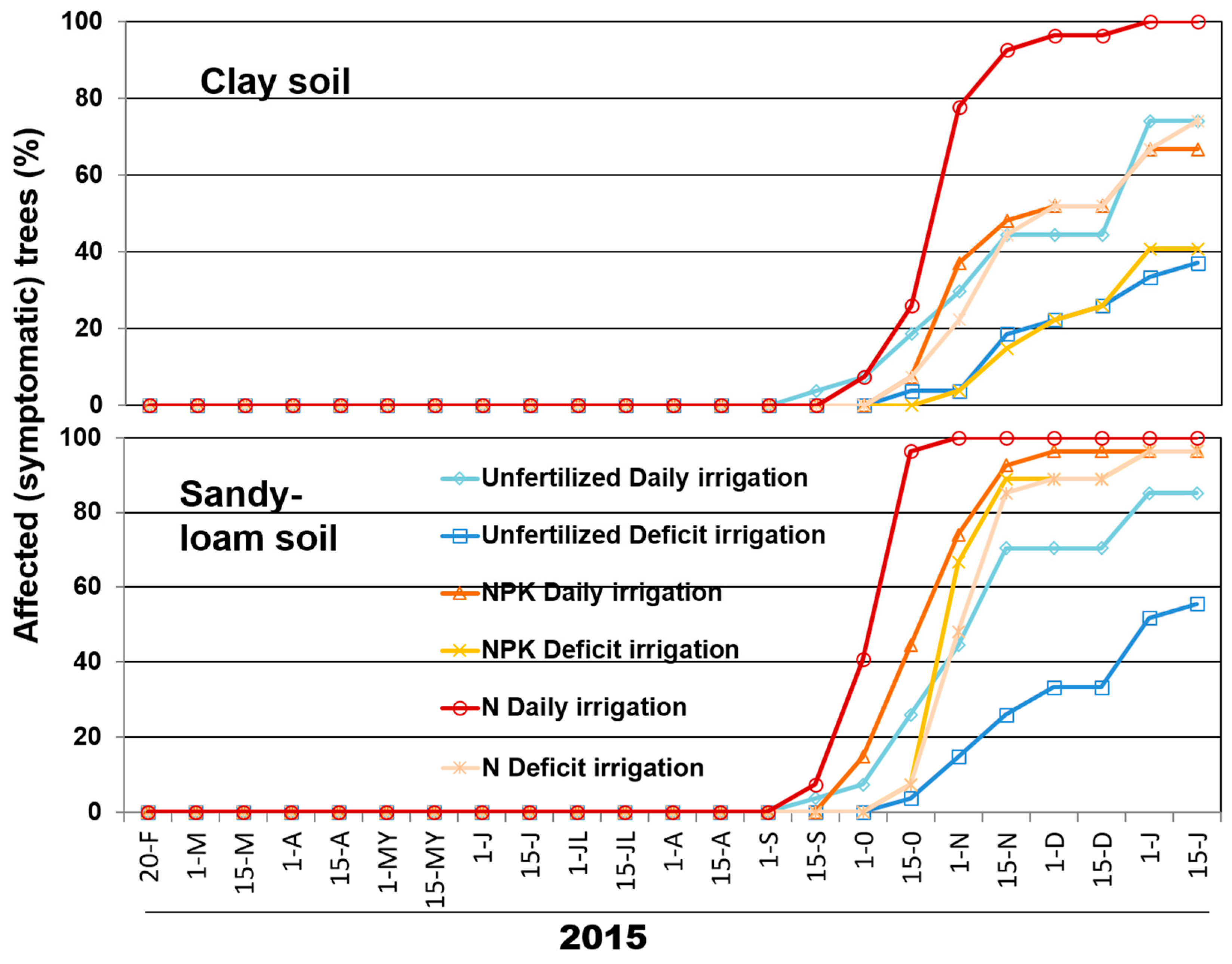
2.2. Inoculum Density of V. dahliae in the Soil and Disease Progression
3. Discussion
4. Materials and Methods
4.1. Experimental Plot
4.2. The Soils: General Characteristics, Inoculum Density of the Pathogen and Physical and Chemical Properties
4.2.1. General Characteristics
4.2.2. Inoculum Density of the Pathogen
4.2.3. Physical and Chemical Properties
4.2.4. Hydraulic Properties
4.3. The Plant Material
4.4. Microplot Establishment, Experimental Design, and Treatments
4.5. Soil Water Content Measurements in the Microplots
4.6. Disease Progression and Tree Development
4.7. Pathogen Isolation
4.8. Final Inoculum Density of the Pathogen in Microplots
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium Wilt of Olive: A Case Study to Implement an Integrated Strategy to Control a Soil-Borne Pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Tsror (Lahkim), L. Epidemiology and Control of Verticillium Wilt on Olive. Isr. J. Plant Sci. 2011, 59, 59–69. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; López-Escudero, F.J. Verticillium Wilt of Olive and Its Control: The Heat Is on. Plant Soil 2012, 355, 17–21. [Google Scholar] [CrossRef]
- Jiménez-Díaz, R.M.; Cirulli, M.; Bubici, G.; del Mar Jiménez-Gasco, M.; Antoniou, P.P.; Tjamos, E.C. Verticillium Wilt, a Major Threat to Olive Production: Current Status and Future Prospects for Its Management. Plant Dis. 2012, 96, 304–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montes-Osuna, N.; Mercado-Blanco, J. Verticillium Wilt of Olive and Its Control: What Did We Learn during the Last Decade? Plants 2020, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Barranco, D.; Fernández-Escobar, R.; Rallo, L. (Eds.) Olive Growing; Junta de Andalucía/Mundi-Prensa/RIRDC/AOA: Pendle Hill, Australia, 2010. [Google Scholar]
- Huber, D.M.; Tompson, I.A. Nitrogen and Plant Disease. In Mineral Nutrition and Plant Disease; Datnoff, L.E., Wade, H.E., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2007; p. 278. [Google Scholar]
- Datnoff, L.E.; Rodrigues, F.A.; Seebold, K.W. Silicon and Plant Disease. In Mineral Nutrition and Plant Disease; Datnoff, L.E., Wade, H.E., Huber, D.M., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2007; pp. 228–247. [Google Scholar]
- Pegg, G.F.; Brady, B.L. (Eds.) Verticillium Wilts; CABI Publishing: Wallingford, UK, 2002; ISBN 0851995292. [Google Scholar]
- McMahon, P. Effect of Nutrition and Soil Function on Pathogens of Tropical Tree Crops. In Plant Pathology; Cumagun, C.J., Ed.; InTech: London, UK, 2012. [Google Scholar]
- Isaac, I. Wilt of Lucerne Caused by Species of Verticillium. Ann. Appl. Biol. 1957, 45, 550–558. [Google Scholar] [CrossRef]
- Sewell, G.W.F.; Wilson, J.F. Verticillium Wilt of the Hop: Field Studies on Wilt in a Resistant Cultivar in Relation to Nitrogen Fertilizer Applications. Ann. Appl. Biol. 1967, 59, 265–273. [Google Scholar] [CrossRef]
- Talboys, P.W. Verticillium Wilt in English Hops: Retrospect and Prospect. Can. J. Plant Pathol. 1987, 9, 68–77. [Google Scholar] [CrossRef]
- Sivaprakasam, K.; Rajagopalan, C.K.S. Effect of Nitrogen on the Incidence of Verticillium Wilt Disease of Eggplant Caused by Verticillium Dahliae Kleb. Plant Soil 1974, 40, 217–220. [Google Scholar] [CrossRef]
- Presley, J.T.; Dick, J.B. Fertilizer and Weather Affect Verticillium Wilt. In Mississippi Farm Research; Mississippi Agricultural and Forestry Experiment Station, Ed.; Universidad de Wisconsin: Madison, WI, USA, 1951; Volume 14–15, p. 6. [Google Scholar]
- Young, V.H.; Fulton, N.D.; Waddle, B.A. Factors Affecting the Incidence and Severity of Verticillium Wilt Disease of Cotton. Bull. Ark. Agric. Exp. Stn. 1959, 612, 26. [Google Scholar]
- Shufelt, C.; Linderman, R.G. Abstracts of Papers Presented at the 4th International Verticillium Symposium, University of Guelph, Guelph, Ontario 17-21 August 1986. Can. J. Plant Pathol. 1987, 9, 78–87. [Google Scholar] [CrossRef]
- Ashworth, L.; Gaona, S.; Surber, E. Verticillium Wilt of Pistachio: The Influence of Potassium Nutrition on Susceptibility to Infection by Verticullium Dahliae. Phytopathology 1985, 75, 1091–1093. [Google Scholar] [CrossRef]
- Emechebe, M. The Effect of Soil Moisture and of N, P and K on Incidence of Infection of Cacao Seedlings Inoculated with Verticillium Dahliae. Plant Soil 1980, 54, 143–147. [Google Scholar] [CrossRef]
- López-Moral, A.; Agustí-Brisach, C.; Alcántara, E.; Trapero, A.; Ruiz-Blancas, C.; Antón-Domínguez, B.I. Elucidating the Effect of Nutritional Imbalances of N and K on the Infection of Verticillium Dahliae in Olive. J. Fungi 2022, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Cappaert, M.R.; Powelson, M.L.; Christensen, N.W.; Crowe, F. Influence of Irrigation on Severity of Potato Early Dying and Tuber Yield. Phytopahthology 1992, 82, 1448–1453. [Google Scholar] [CrossRef]
- Xiao, C.L.; Subbarao, K. V Effects of Irrigation and Verticillium Dahliae on Cauliflower Root and Shoot Growth Dynamics. Phytopathology 2000, 90, 995–1004. [Google Scholar] [CrossRef] [Green Version]
- Bletsos, F.A.F.; Thanassoulopoulos, C.C.; Roupakias, D.G. Water Stress and Verticillium Wilt Severity on Eggplant (Solanummelongena, L.). J. Phytopathol. 1999, 147, 243–248. [Google Scholar] [CrossRef]
- Wheeler, T.A.; Bordovsky, J.P.; Keeling, J.W.; Mullinix, B.G.; Woodward, J.E. Effects of Crop Rotation, Cultivar, and Irrigation and Nitrogen Rate on Verticillium Wilt in Cotton. Plant Dis. 2012, 96, 985–989. [Google Scholar] [CrossRef] [Green Version]
- Vigouroux, A. Verticilliose et Bact´eriose Deux Importants Facteurs de D´ep´erissment de L’abricotier. Arboric. Fruit. 1984, 31, 31–35. [Google Scholar]
- Pérez-Rodríguez, M.; Alcántara, E.; Amaro, M.; Serrano, N.; Lorite, I.J.; Orgaz, F.; López-escudero, F.J. The Influence of Irrigation Frequency on the Onset and Development of Verticillium Wilt of Olive. Plant Dis. 2015, 99, 488–495. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rodríguez, M.; Serrano, N.; Arquero, O.; Orgaz, F.; Moral, J.; López-escudero, F.J. The Effect of Short Irrigation Frequencies on the Development of Verticillium Wilt in the Susceptible Olive Cultivar “Picual” at Field Conditions. Plant Dis. 2016, 100, 1880–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Rodríguez, M.; Orgaz, F.; Lorite, I.J.; López-Escudero, F.J. Effect of the Irrigation Dose on Verticillium Wilt of Olive. Sci. Hortic. 2015, 197, 564–567. [Google Scholar] [CrossRef]
- Santos-Rufo, A.; Vega, V.; Hidalgo, J.C.; Hidalgo, J.J.; Rodríguez-Jurado, D. Assessment of the Effect of Surface Drip Irrigation on Verticillium Dahliae Propagules Differing in Persistence in Soil and on Verticillium Wilt of Olive. Plant Pathol. 2017, 66, 1117–1127. [Google Scholar] [CrossRef]
- Varo, A.; Raya-Ortega, M.C.; Trapero, A. Selection and Evaluation of Microorganisms for Biocontrol of Verticillium Dahliae in Olive. J. Appl. Microbiol. 2016, 121, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Barker, A.V. Mineral Nutrition and Plant Disease. HortScience 2009, 44, 1510. [Google Scholar] [CrossRef] [Green Version]
- Pennypacker, B. The Role of Mineral Nutrition in the Control of Verticillium Wilt. In Soilborne Plant Pathogens: Management of Diseases with Macro- and Microelements; Engelhard, A., Ed.; American Phytopathological Society: St. Paul, MN, USA, 1989; pp. 32–45. [Google Scholar]
- Santos-Rufo, A.; Hidalgo, J.J.; Hidalgo, J.C.; Vega, V.; Rodríguez-Jurado, D. Morphophysiological Response of Young Olive Trees to Verticillium Wilt under Different Surface Drip Irrigation Regimes. Plant Pathol. 2018, 67, 848–859. [Google Scholar] [CrossRef]
- Pérez-López, D.; Ribas, F.; Moriana, A.; Olmedilla, N.; de Juan, A. The Effect of Irrigation Schedules on the Water Relations and Growth of a Young Olive (Olea Europaea, L.) Orchard. Agric. Water Manag. 2007, 89, 297–304. [Google Scholar] [CrossRef]
- Carns, H.R.; Presley, J.T.; Schnathorst, W.C.; Harrison, G.K.; Taylor, E.E. Proceedings of the Beltwide Cotton Production Research Conference. In Proceedings of the Beltwide Cotton Production Research Conference, Memphis, TN, USA, 8–11 January 1964; pp. 83–89. [Google Scholar]
- Chernyayeva, I.I.; Hasanova, F.S.; Muromtsev, G.S. Effect of Various Nitrogen Forms upon Verticillium Dahliae and the Metabolism of Some Soil Bacteria. In Soil Biology and Conservation of the Biosphere; Szeji, J., Ed.; Academiai Kiado: Budapest, Hungary, 1984; pp. 703–710. [Google Scholar]
- Muromtsev, G.S.; Chernyaeva, I.I. The Effects of Various Form of Nitrogen Fertilizer on the Viability of Verticillium Dahliae, the Causative Organism of Cotton Wilt. Dokl. Vsesoyuznoi Akad. Sel’skokhozyaistvennykh Nauk Im. V. I. LeninaSel’skokhozyaistvennykh Nauk Im. V. I. Lenina 1979, 12, 7–9. [Google Scholar]
- Santos-Rufo, A.; Pérez-Rodriguez, M.; Heis Serrano, J.; Roca Castillo, L.F.; López-Escudero, F.J. Effect of Previous Crops and Soil Physicochemical Properties on the Population of Verticillium Dahliae in the Iberian Peninsula. J. Fungi 2022, 8, 988. [Google Scholar] [CrossRef]
- López-Escudero, F.J.; Mercado-Blanco, J.; Roca, J.M.; Valverde-Corredor, A.; Blanco-López, M.A. Verticillium Wilt of Olive in the Guadalquivir Valley (Southern Spain): Relations with Some Agronomical Factors and Spread of Verticillium Dahliae. Phytopathol. Mediterr. 2010, 49, 370–380. [Google Scholar]
- Staff, S.S. Keys to Soil Taxonomy; United States Department of Agriculture Natural Resources Conservation Service: Washington DC, USA, 2014; p. 360. [Google Scholar]
- Trapero, C.; Serrano, N.; Arquero, O.; Del Río, C.; Trapero, A.; López-Escudero, F.J. Field Resistance to Verticillium Wilt in Selected Olive Cultivars Grown in Two Naturally Infested Soils. Plant Dis. 2013, 97, 668–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valverde, P.; Trapero, C.; Arquero, O.; Serrano, N.; Barranco, D.; Muñoz Díez, C.; López-Escudero, F.J. Highly Infested Soils Undermine the Use of Resistant Olive Rootstocks as a Control Method of Verticillium Wilt. Plant Pathol. 2021, 70, 144–153. [Google Scholar] [CrossRef]
- Huisman, O.C.; Ashworth, L.J., Jr. Verticillium Albo-Atrum: Quantitative Isolation of Microsclerotia From Field Soils. Phytopathology 1974, 64, 1159. [Google Scholar] [CrossRef]
- Butterfield, E.J.; DeVay, J.E. Reassessment of Soil Assays for Verticillium Dahliae. Phytopathology 1977, 67, 1073–1078. [Google Scholar] [CrossRef]
- López-Escudero, F.J.; Blanco-Lápez, M.A. Isolation and Morphologic Characterization of Microsclerotia of Verticillium Dahliae Isolate from Soil. Biotechnology 2005, 4, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Mercado-Blanco, J.; Rodríguez-Jurado, D.; Parrilla-Araujo, S.; Jiménez-Díaz, R.M. Simultaneous Detection of the Defoliating and Nondefoliating Verticillium Dahliae Pathotypes in Infected Olive Plants by Duplex, Nested Polymerase Chain Reaction. Plant Dis. 2003, 87, 1487–1494. [Google Scholar] [CrossRef] [Green Version]
- López-Escudero, F.J.; del Río, C.; Caballero, J.M.; Blanco-López, M.A. Evaluation of Olive Cultivars for Resistance to Verticillium Dahliae. Eur. J. Plant Pathol. 2004, 110, 79–85. [Google Scholar] [CrossRef]
- Mulero-Aparicio, A.; Trapero, A.; Javier, F. A Non-Pathogenic Strain of Fusarium Oxysporum and Grape Marc Compost Control Verticillium Wilt of Olive. Phytopathol. Mediterr. 2020, 59, 159–167. [Google Scholar] [CrossRef]
- Hau, B.; Amorim, L.; Bergamin Filho, A. Mathematical Functions to Describe Disease Progress Curves of Double Sigmoid Pattern. Phytopathology 1993, 83, 928–932. [Google Scholar] [CrossRef]
- Navas-Cortés, J.A.; Landa, B.B.; Mercado-Blanco, J.; Trapero-Casas, J.L.; Rodríguez-Jurado, D.; Jiménez-Díaz, R.M. Spatiotemporal Analysis of Spread of Infections by Verticillium Dahliae Pathotypes Within a High Tree Density Olive Orchard in Southern Spain. Phytopathology 2008, 98, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; John Wiley & Sons: New York, NY, USA, 1990; ISBN 0471832367. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fertilization | Irrigation | FS (Range) b | Mortality (%) c | MS (%) d | Incidence (%) | MM (Weeks) e | RAUDPC f |
|---|---|---|---|---|---|---|---|
| Unfertilized | Daily | 40.6 a (31–48) | 18.5 a | 29.1 a | 74.1 a | 3.7 a | 51.3 a |
| Deficit | 42.9 a (35–50) | 7.4 a | 20.7 a | 37.0 a | 6.0 a | 22.9 a | |
| Mean | 41.7 A | 12.9 B | 24.9 B | 55.5 A | 4.8 A | 37.1 B | |
| NPK | Daily | 39.3 a (35–46) | 18.5 a | 29.3 a | 66.7 a | 6.0 a | 17.5 a |
| Deficit | 42.7 a (37–46) | 3.7 a | 24.9 a | 40.7 a | 6.0 a | 20.7 a | |
| Mean | 41.0 A | 11.1 B | 27.1 B | 53.7 A | 6.0 A | 34.1 B | |
| N | Daily | 41.5 a (35–50) | 37.0 a | 64.7 a | 100.0 a | 7.0 a | 89.2 a |
| Deficit | 37.3 a (33–46) | 25.9 a | 49.4 a | 74.1 a | 7.7 a | 50.8 a | |
| Mean | 39.4 A | 31.4 A | 57.1 A | 87.0 A | 7.3 A | 70.0 A |
| Fertilization | Irrigation | FS (Range) b | Mortality (%) c | MS (%) d | Incidence (%) | MM (Weeks) e | RAUDPC f |
|---|---|---|---|---|---|---|---|
| Unfertilized | Daily | 38.6 a (31–48) | 25.9 a | 47.6 a | 85.2 a | 5.8 a | 37.8 c |
| Deficit | 41.8 a (35–50) | 0.0 a | 23.0 a | 55.6 b | … | 9.6 d | |
| Mean | 40.2 B | 12.9 B | 35.3 B | 70.4 A | 23.8 B | ||
| NPK | Daily | 36.4 a (33–42) | 48.1 a | 74.4 a | 96.3 a | 5.7 a | 61.1 b |
| Deficit | 38.0 a (35–46) | 48.1 a | 71.9 a | 96.3 a | 9.1 a | 44.9 c | |
| Mean | 37.2 A | 48.1 A | 73.1 A | 96.3 A | 52.9 A | ||
| N | Daily | 34.1 a (31–46) | 85.2 a | 90.9 a | 100 a | 3.7 b | 94.3 a |
| Deficit | 38.8 a (35–46) | 22.2 a | 67.5 a | 96.3 a | 6.8 a | 42.6 c | |
| Mean | 36.4 A | 53.7 A | 79.2 A | 98.1 A | 68.4 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Rodríguez, M.; Santos-Rufo, A.; López-Escudero, F.J. High Input of Nitrogen Fertilization and Short Irrigation Frequencies Forcefully Promote the Development of Verticillium Wilt of Olive. Plants 2022, 11, 3551. https://doi.org/10.3390/plants11243551
Pérez-Rodríguez M, Santos-Rufo A, López-Escudero FJ. High Input of Nitrogen Fertilization and Short Irrigation Frequencies Forcefully Promote the Development of Verticillium Wilt of Olive. Plants. 2022; 11(24):3551. https://doi.org/10.3390/plants11243551
Chicago/Turabian StylePérez-Rodríguez, Mario, Antonio Santos-Rufo, and Francisco Javier López-Escudero. 2022. "High Input of Nitrogen Fertilization and Short Irrigation Frequencies Forcefully Promote the Development of Verticillium Wilt of Olive" Plants 11, no. 24: 3551. https://doi.org/10.3390/plants11243551