

Effects of Microplastic Contamination on the Aquatic Plant Lemna minuta (Least Duckweed)
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
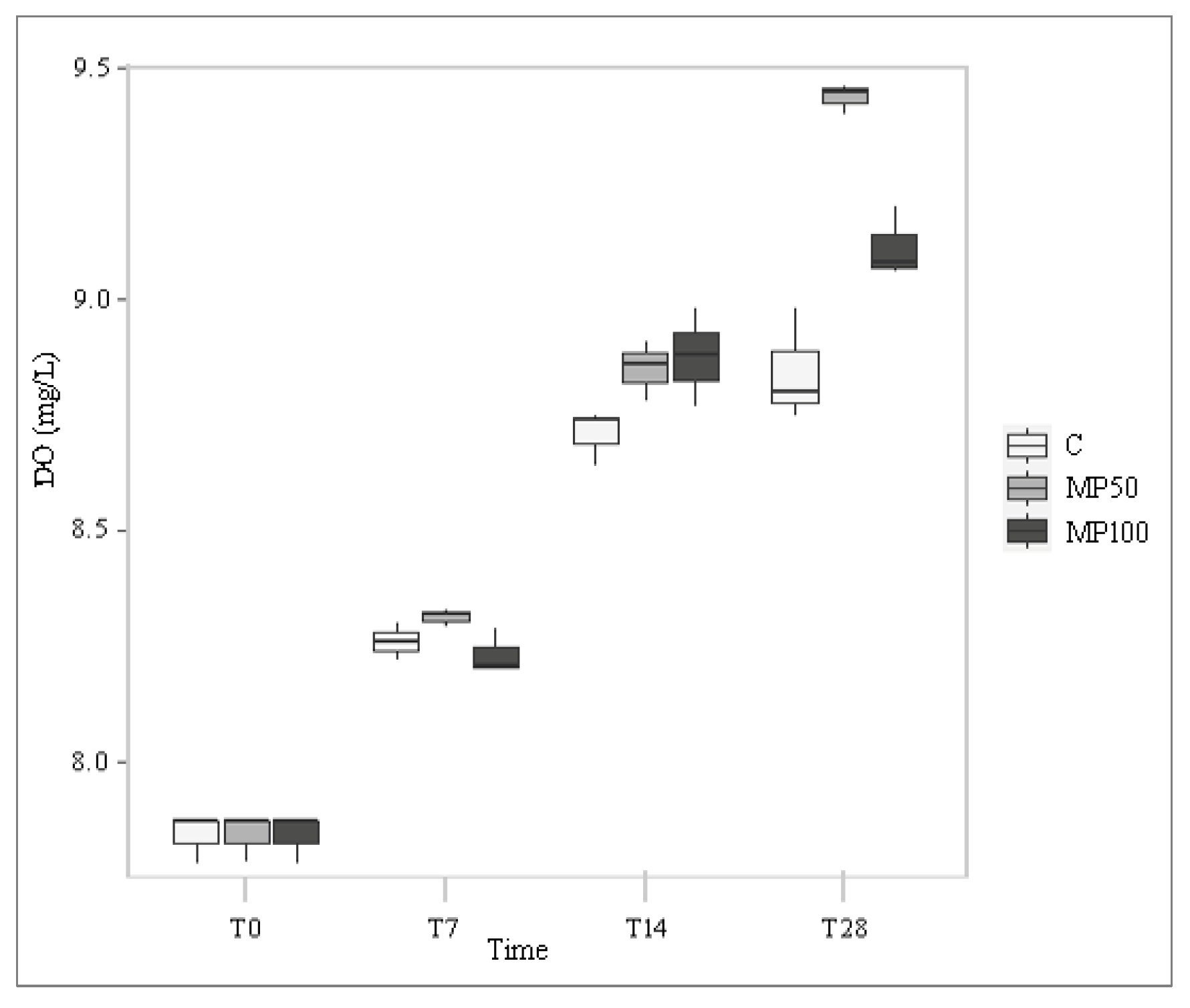
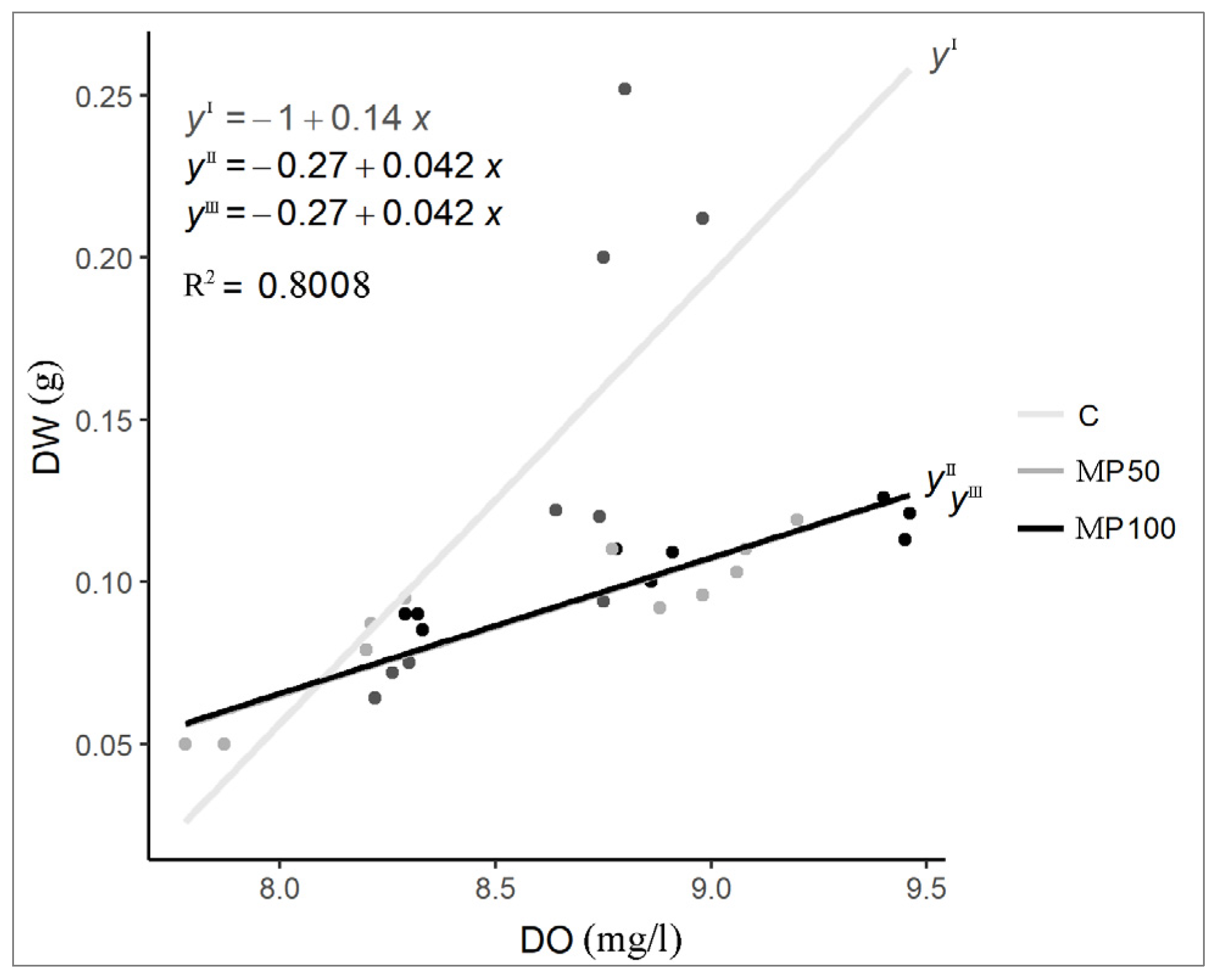
2.1. Water Chemical and Physical Parameters
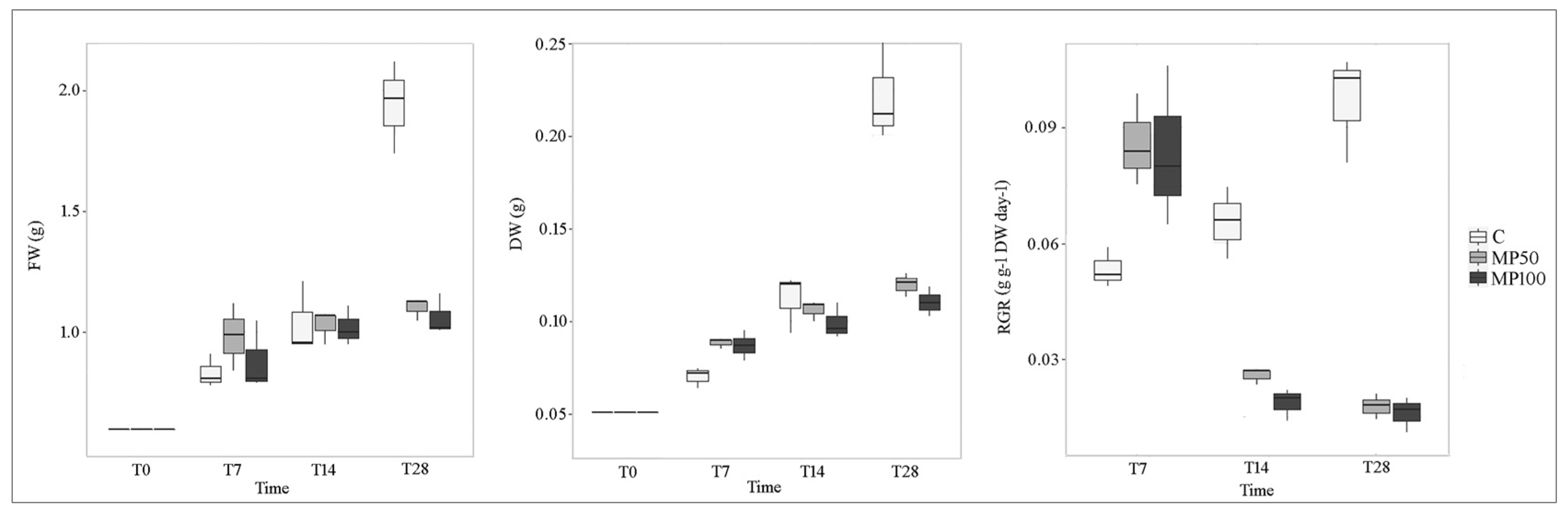
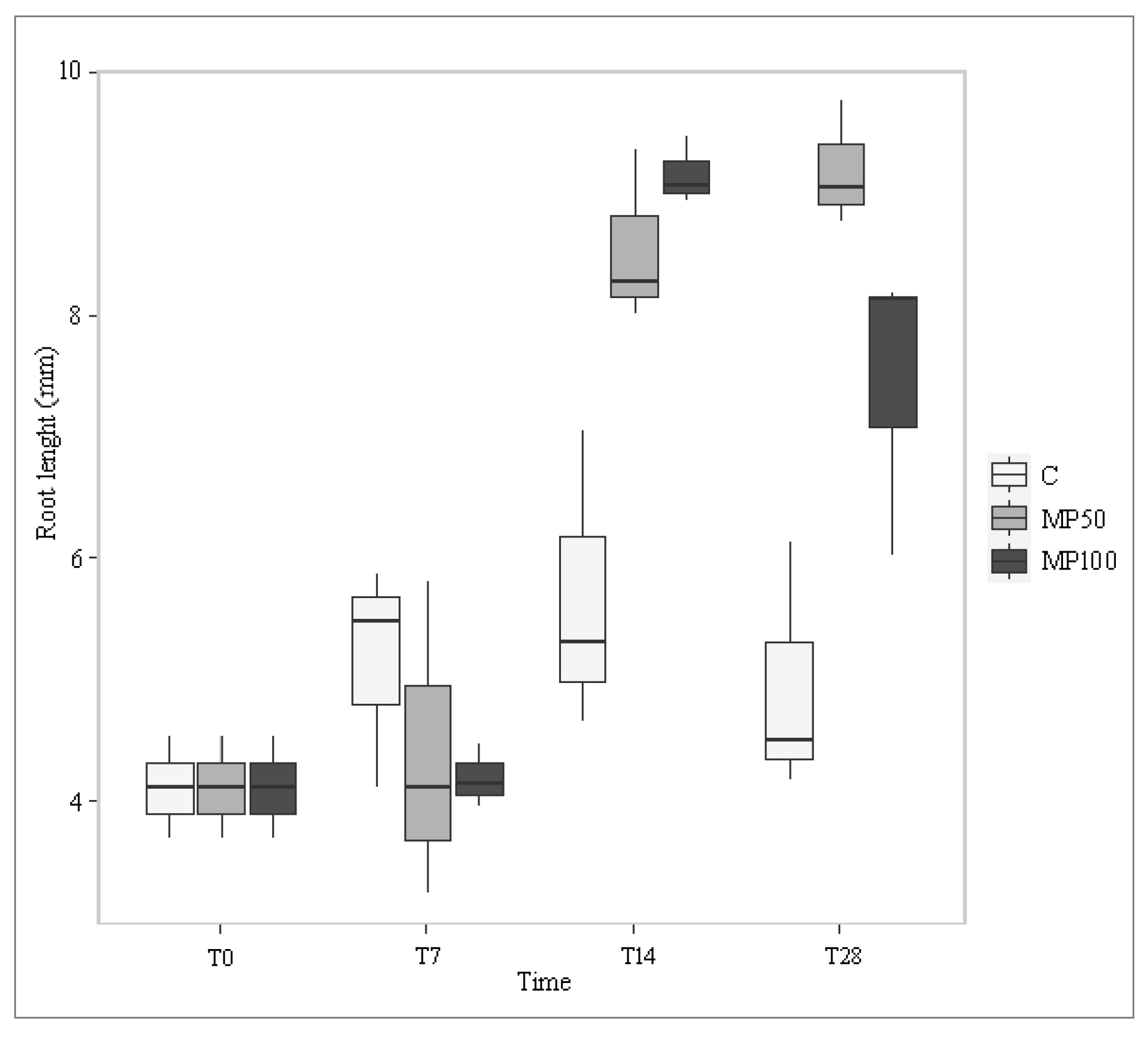
2.2. Effects of Microplastics on Plant Growth Parameters
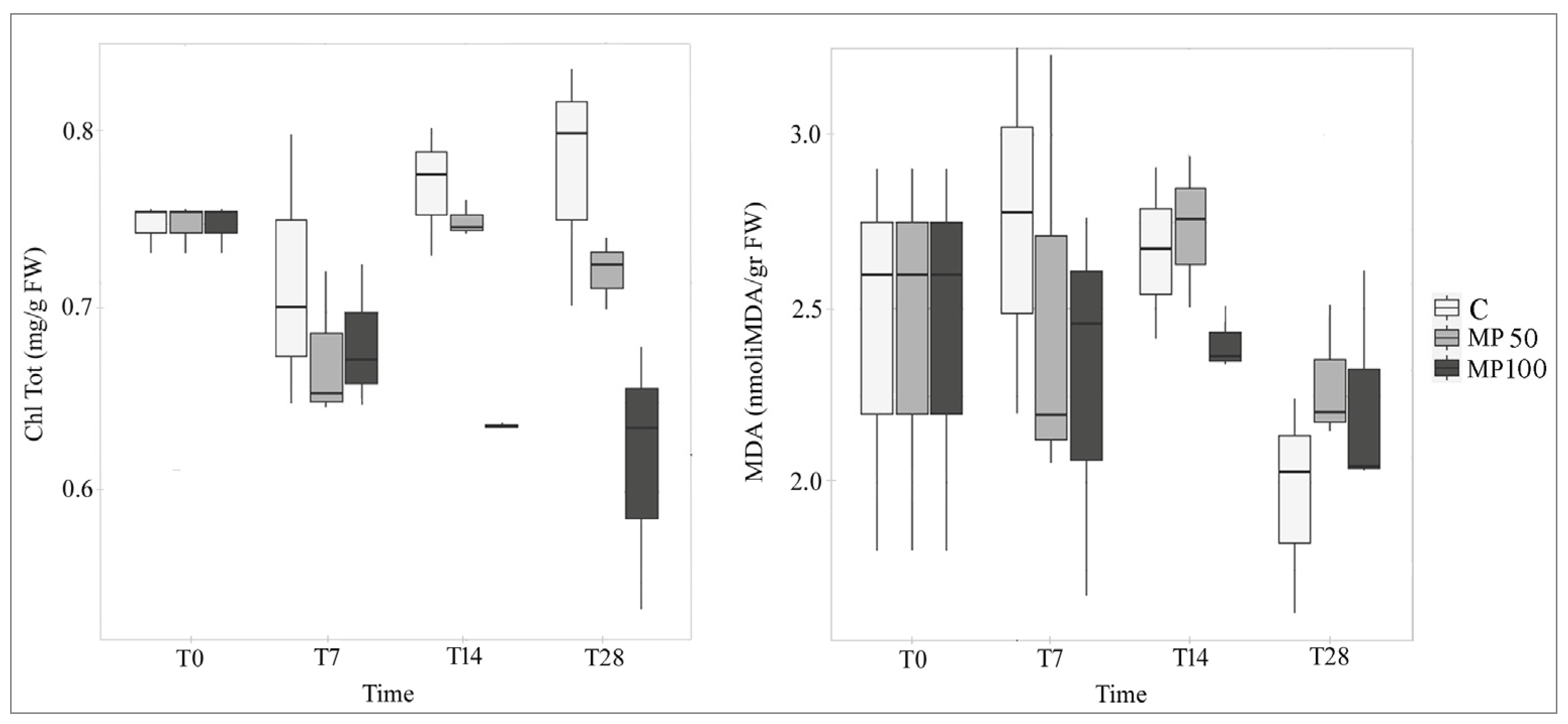
2.3. Effects of MP on Biochemical Parameters
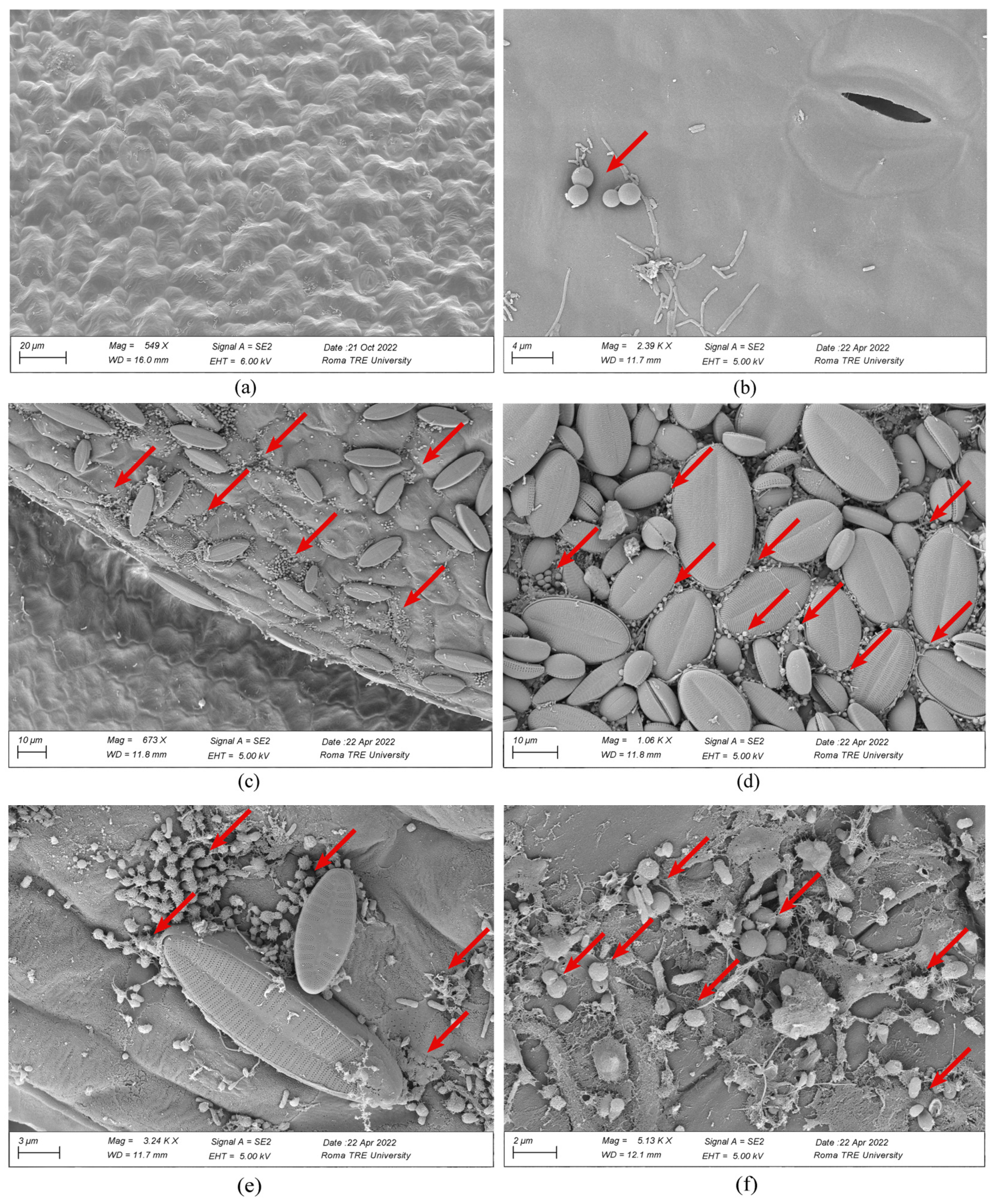
2.4. Adsorption of MP on Plants
3. Materials and Methods
3.1. Production of Microplastics
3.2. Plant Material and Experimental Set-Up
3.3. Determination of Water Chemical and Physical Parameters
3.4. Plant Growth and Morphological Measurements
3.4.1. Plant Growth Analysis
3.4.2. Morphological Analysis
3.5. Analysis of Biochemical Parameters
3.5.1. Determination of Chlorophyll Content
3.5.2. Determination of Malondialdehyde Content
3.6. SEM Observations of Microplastics
3.7. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cole, M.; Lindeque, P.; Halsband, C.; Galloway, T.S. Microplastics as contaminants in the marine environment: A review. Mar. Pollut. Bull. 2011, 62, 2588–2597. [Google Scholar] [CrossRef] [PubMed]
- Azeem, I.; Adeel, M.; Ahmad, M.A.; Shakoor, N.; Jiangcuo, G.D.; Azeem, K.; Ishfaq, M.; Shakoor, A.; Ayaz, M.; Xu, M.; et al. Uptake and Accumulation of Nano/Microplastics in Plants: A Critical Review. Nanomaterials 2021, 11, 2935. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.G.; McGonigle, D.; Russel, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.G.; Moore, C.J.; Van Franeker, J.A.; Moloney, C.L. Monitoring the abundance of plastic debris in the marine environment. Philosoph. Trans. R. Soc. B Biol. Sci. 2009, 364, 1999–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereao, O.; Opeolu, B.; Fatoki, O. Microplastics in aquatic environment: Characterization, ecotoxicological effect, implications for ecosystems and developments in South Africa. Environ. Sci. Pollut. Res. 2020, 27, 22271–22291. [Google Scholar] [CrossRef]
- Almroth, C.B.; Eggert, H. Marine plastic pollution: Sources, impacts, and policy issues. Rev. Env. Econ. Policy 2019, 13, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Ma, P.; Wei Wang, M.; Liu, H.; Feng Chen, Y.; Xia, J. Research on ecotoxicology of microplastics on freshwater aquatic organisms. Env. Poll. Bioavailab. 2019, 31, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Barnes, D.K.A.; Galgani, F.; Thompson, R.C.; Barlaz, M. Accumulation and fragmentation of plastic debris in global environments. Philosoph. Trans. R. Soc. B Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [Green Version]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R. Accumulation of microplastic on shorelines worldwide: Sources and sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef]
- Arossa, S.; Martin, C.; Rossbach, S.; Duarte, C.M. Microplastic removal by Red Sea giant clam (Tridacna maxima). Environ. Pollut. 2019, 252, 1257–1266. [Google Scholar] [CrossRef]
- Horton, A.A.; Walton, A.; Spurgeon, D.J.; Lahive, E.; Svendsen, C. Microplastics in freshwater and terrestrial environments: Evaluating the current understanding to identify the knowledge gaps and future research priorities. Sci. Total Environ. 2017, 586, 127–141. [Google Scholar] [CrossRef] [Green Version]
- Zbyszewski, M.; Corcoran, P.L. Distribution and degradation of freshwater plastic particles along the beaches of Lake Huron, Canada. Water Air Soil Pollut. 2011, 220, 365–372. [Google Scholar] [CrossRef]
- Zbyszewski, M.; Corcoran, P.L.; Hockin, A. Comparison of the distribution and degradation of plastic debris along shorelines of the Great Lakes, North America. J. Great Lakes Res. 2014, 40, 288–299. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Besseling, E.; Shim, W.J. Nanoplastics in the aquatic environment. Critical review. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer: Cham, Switzerland, 2015; pp. 325–340. [Google Scholar]
- Mattsson, K.; Hansson, L.A.; Cedervall, T. Nano-plastics in the aquatic environment. Environ. Sci. Process. Impacts 2015, 17, 1712–1721. [Google Scholar] [CrossRef]
- Van Sebille, E.; Wilcox, C.; Lebreton, L.; Maximenko, N.; Hardesty, B.D.; Van Franeker, J.A.; Eriksen, M.; Siegel, D.; Galgani, F.; Law, K.L. A global inventory of small floating plastic debris. Environ. Res. Lett. 2015, 10, 124006. [Google Scholar] [CrossRef]
- Rillig, M.C.; Lehmann, A.; de Souza Machado, A.A.; Yang, G. Microplastic effects on plants. New Phytol. 2019, 223, 1066–1070. [Google Scholar] [CrossRef] [Green Version]
- Kalčíková, G. Aquatic vascular plants—A forgotten piece of nature in microplastic research. Environ. Pollut. 2020, 262, 114354. [Google Scholar] [CrossRef]
- Ceschin, S.; Bellini, A.; Scalici, M. Aquatic plants and ecotoxicological assessment in freshwater ecosystems: A review. Environ. Sci. Pollut. Res. 2021, 28, 4975–4988. [Google Scholar] [CrossRef]
- Mateos-Cárdenas, A.; Scott, D.T.; Seitmaganbetova, G.; Van Pelt, F.N.A.M.; O’Halloran, J.; Jansen, M.A.K. Polyethylene microplastics adhere to Lemna minor (L.) yet have no effects on plant growth or feeding by Gammarus duebeni (Lillj.). Sci. Total Environ. 2019, 689, 413–421. [Google Scholar] [CrossRef]
- Rozman, U.; Kokalj, A.J.; Dolar, A.; Drobne, D.; Kalčíková, G. Long-term interactions between microplastics and floating macrophyte Lemna minor: The potential for phytoremediation of microplastics in the aquatic environment. Sci. Total Environ. 2022, 831, 154866. [Google Scholar] [CrossRef]
- Kalčíková, G.; Žgajnar Gotvajn, A.; Kladnik, A.; Jemec, A. Impact of polyethylene microbeads on the floating freshwater plant duckweed Lemna minor. Environ. Pollut. 2017, 230, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Bosker, T.; Bouwman, L.J.; Brun, N.R.; Behrens, P.; Vijver, M.G. Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant Lepidium sativum. Chemosphere 2019, 226, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Van Weert, S.; Redondo-Hasselerharm, P.E.; Diepens, N.J.; Koelmans, A.A. Effects of nanoplastics and microplastics on the growth of sediment-rooted macrophytes. Sci. Total Environ. 2019, 654, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Chi, T.; Li, Z.; Wang, J.; Yu, M.; Zhou, H. Combined effect of polystyrene plastics and triphenyltin chloride on the green algae Chlorella pyrenoidosa. Environ. Sci. Pollut. Res. 2019, 26, 15011–15018. [Google Scholar] [CrossRef] [PubMed]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Marchetto, D.; Latella, A.; Pojana, G. Le microplastiche nell’ambiente marino. Chim. Ambiente 2017, 1, 18–25. [Google Scholar]
- Ceschin, S.; Della Bella, V.; Piccari, F.; Abati, S. Colonization dynamics of the alien macrophyte Lemna minuta Kunth: A case study from a semi-natural pond in Appia Antica Regional Park (Rome, Italy). Fundam. Appl. Limnol. 2016, 188, 93–101. [Google Scholar] [CrossRef]
- Van Hoeck, A.; Horemans, N.; Monsieurs, P.; Cao, H.X.; Vandenhove, H.; Blust, R. The first draft genome of the aquatic model plant Lemna minor opens the route for future stress physiology research and biotechnological applications. Biotechnol. Biofuels 2015, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Rezania, S.; Taib, S.M.; Md Din, M.F.; Dahalan, F.A.; Kamyab, H. Comprehensive review on phytotechnology: Heavy metals removal by diverse aquatic plants species from wastewater. J. Hazard. Mater. 2016, 318, 587–599. [Google Scholar] [CrossRef]
- Ekperusi, A.O.; Sikoki, F.D.; Nwachukwu, E.O. Application of common duckweed (Lemna minor) in phytoremediation of chemicals in the environment: State and future perspective. Chemosphere 2019, 223, 285–309. [Google Scholar] [CrossRef]
- Dovidat, L.C.; Brinkmann, B.W.; Vijver, M.G.; Bosker, T. Plastic particles adsorb to the roots of freshwater vascular plant Spirodela polyrhiza but do not impair growth. Limnol. Oceanogr. Lett. 2020, 5, 37–45. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Lin, S.; Turner, J.P.; Ke, P.C. Physical adsorption of charged plastic nanoparticles affects algal photosynthesis. J. Phys. Chem. C 2010, 114, 16556–16561. [Google Scholar] [CrossRef]
- Schaffelke, B. Particulate organic matter as an alternative nutrient source for tropical sargassum species (Fucales, phaeophyceae). J. Phycol. 1999, 35, 1150–1157. [Google Scholar] [CrossRef]
- Gutow, L.; Eckerlebe, A.; Gim_enez, L.; Saborowski, R. Experimental evaluation of seaweeds as a vector for microplastics into marine food webs. Environ. Sci. Technol. 2016, 50, 915–923. [Google Scholar] [CrossRef]
- Savio, S.; Farrotti, S.; Di Giulio, A.; De Santis, S.; Ellwood, N.T.W.; Ceschin, S.; Congestri, R. Functionalization of frustules of the diatom Staurosirella pinnata for nickel (Ni) adsorption from contaminated aqueous solutions. Front. Mar. Sci. 2022, 9, 889832. [Google Scholar] [CrossRef]
- Goss, H.; Jaskiel, J.; Rotjan, R. Thalassia testudinum as a potential vector for incorporating microplastics into benthic marine food webs. Mar. Pollut. Bull. 2018, 135, 1085–1089. [Google Scholar] [CrossRef]
- Frazer, R.Q.; Byron, R.T.; Osborne, P.B.; West, K.P. PMMA: An essential material in medicine and dentistry. J. Long. Term. Eff. Med. Implants 2005, 15, 629–639. [Google Scholar] [CrossRef] [Green Version]
- Ceschin, S.; Abati, S.; Traversetti, L.; Spani, F.; Del Grosso, F.; Scalici, M. Effects of the invasive duckweed Lemna minuta on aquatic animals: Evidence from an indoor experiment. Plant Biosyst. 2019, 153, 749–755. [Google Scholar] [CrossRef]
- Ceschin, S.; Ellwood, N.T.W.; Ferrante, G.; Mariani, F.; Traversetti, L. Habitat change and alteration of plant and invertebrate communities in waterbodies dominated by the invasive alien macrophyte Lemna minuta Kunth. Biol. Invasions 2020, 22, 1325–1337. [Google Scholar] [CrossRef]
- Prata, J.C.; Da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Effects of microplastics on microalgae populations: A critical review. Sci. Total Environ. 2019, 665, 400–405. [Google Scholar] [CrossRef]
- Mateos-Cárdenas, A.; van Pelt, F.N.; O’Halloran, J.; Jansen, M.A. Adsorption, uptake and toxicity of micro-and nanoplastics: Effects on terrestrial plants and aquatic macrophytes. Environ. Pollut. 2021, 284, 117183. [Google Scholar] [CrossRef] [PubMed]
- Nava, V.; Leoni, B. A critical review of interactions between microplastics, microalgae and aquatic ecosystem function. Water Res. 2021, 188, 116476. [Google Scholar] [CrossRef] [PubMed]
- Chronopoulou, L.; Fratoddi, I.; Palocci, C.; Venditti, I.; Russo, M.V. Osmosis based method drives the self-assembly of polymeric chains into micro-and nanostructures. Langmuir 2009, 25, 11940–11946. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Chatterjee, S. Microplastic pollution, a threat to marine ecosystem and human health: A short review. Environ. Sci. Pollut. Res. 2017, 24, 21530–21547. [Google Scholar] [CrossRef]
- Radford, P.J. Growth analysis formulae—Their use and abuse. Crop. Sci. 1967, 7, 171–175. [Google Scholar] [CrossRef]
- Huang, F.; Guo, Z.; Xu, Z. Determined methods of chlorophyll from Lemma paucicostata. Exp. Technol. Manag. 2007, 24, 29–31. [Google Scholar]
- Kassambara, A. R Package, Version 0.4.0. ggpubr: ‘ggplot2’ Based Publication Ready Plots. R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Lüdecke, D. R Package, Version 2.8.11. sjPlot: Data Visualization for Statistics in Social Science. CERN, European Organization for Nuclear Research: Genève, Switzerland, 2022.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- ISO. ISO 20079; Water Quality—Determination of the Toxic Effect of Water Constituents and Wastewater on Duckweed (Lemna minor)—Duckweed Growth Inhibition Test. International Organization for Standardization: Geneve, Switzerland, 2005; pp. 1–23.
- OECD. OECD Test No. 221: Lemna sp. Growth Inhibition Test; OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2006; pp. 1–22. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceschin, S.; Mariani, F.; Di Lernia, D.; Venditti, I.; Pelella, E.; Iannelli, M.A. Effects of Microplastic Contamination on the Aquatic Plant Lemna minuta (Least Duckweed). Plants 2023, 12, 207. https://doi.org/10.3390/plants12010207
Ceschin S, Mariani F, Di Lernia D, Venditti I, Pelella E, Iannelli MA. Effects of Microplastic Contamination on the Aquatic Plant Lemna minuta (Least Duckweed). Plants. 2023; 12(1):207. https://doi.org/10.3390/plants12010207
Chicago/Turabian StyleCeschin, Simona, Flaminia Mariani, Dario Di Lernia, Iole Venditti, Emanuele Pelella, and Maria Adelaide Iannelli. 2023. "Effects of Microplastic Contamination on the Aquatic Plant Lemna minuta (Least Duckweed)" Plants 12, no. 1: 207. https://doi.org/10.3390/plants12010207