
Species- and Metal-Specific Responses of the Ionome of Three Duckweed Species under Chromate and Nickel Treatments
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culturing Conditions and Experimental Setup
2.2. Measurement of Growth
2.3. Determination of Metal Content in the Growth Medium and Biomass
2.4. mXRF Analyses
2.5. Data Processing and Statistics
3. Results and Discussion
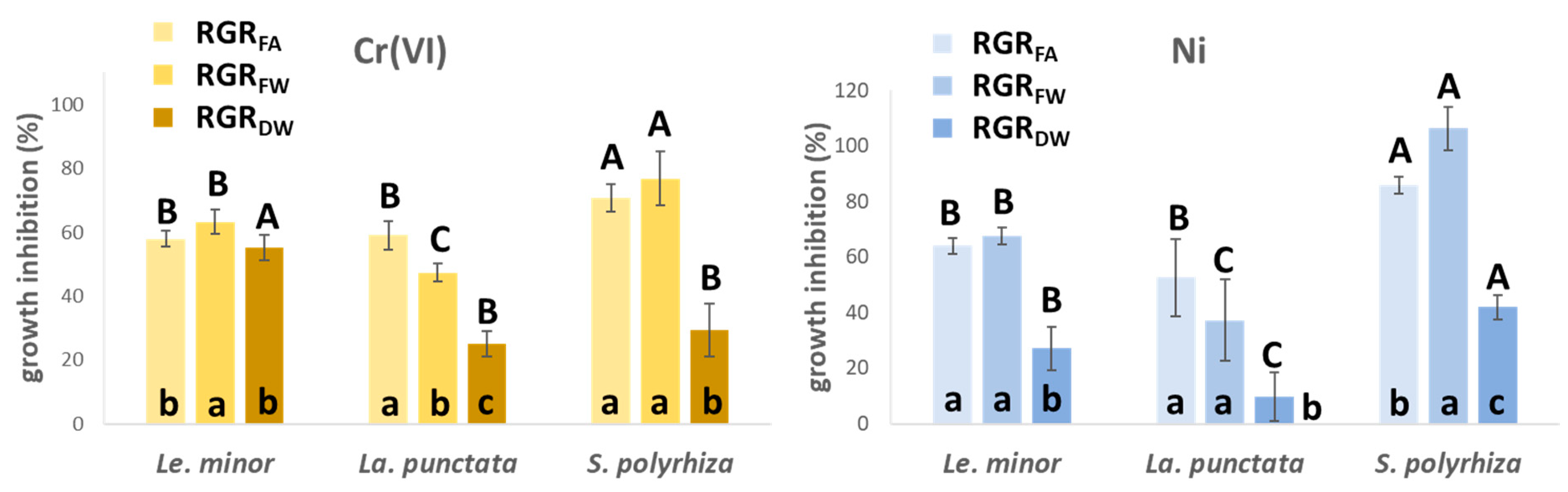
3.1. Metal-Induced Growth Inhibition
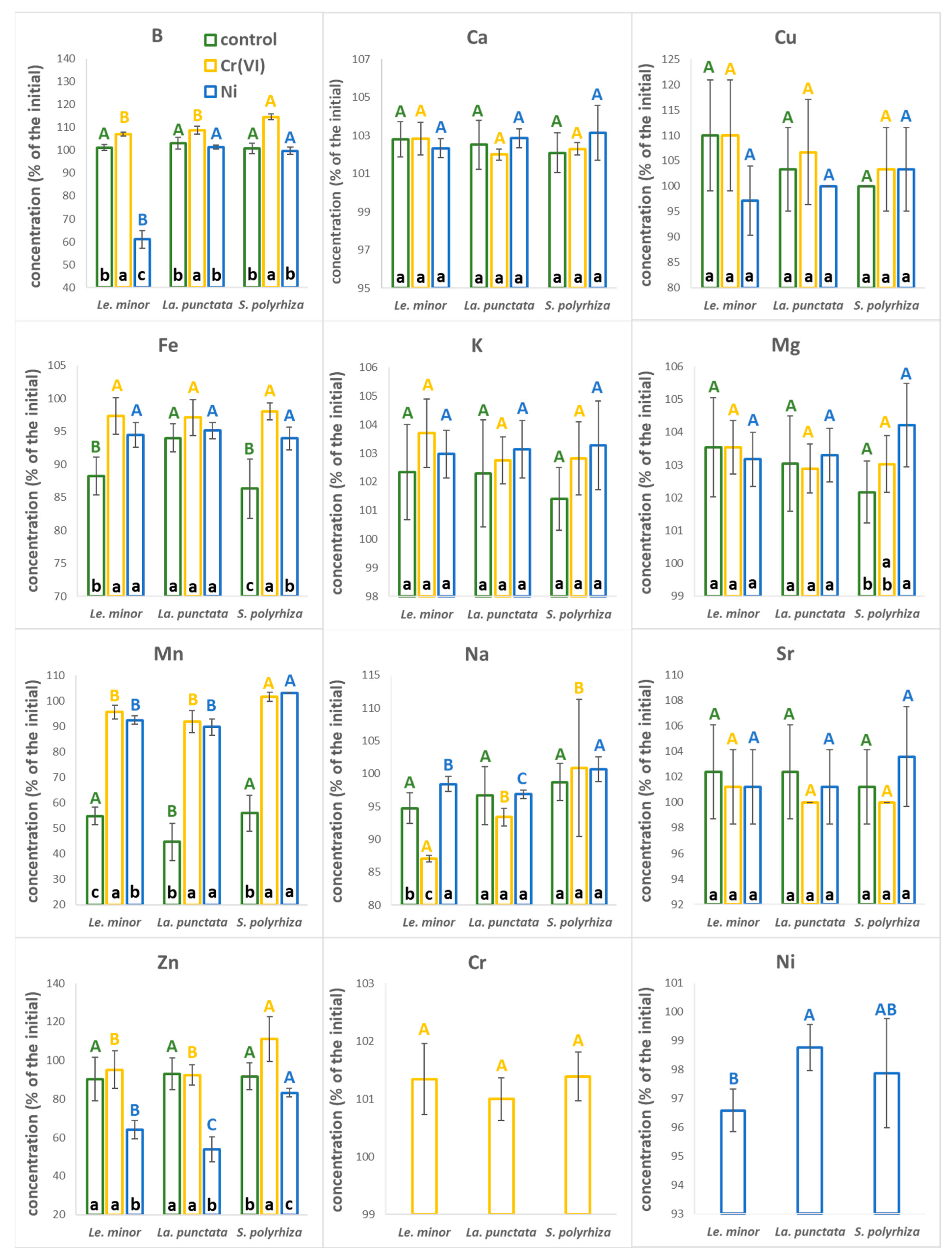
3.2. Changes in the Composition of the Nutrient Medium
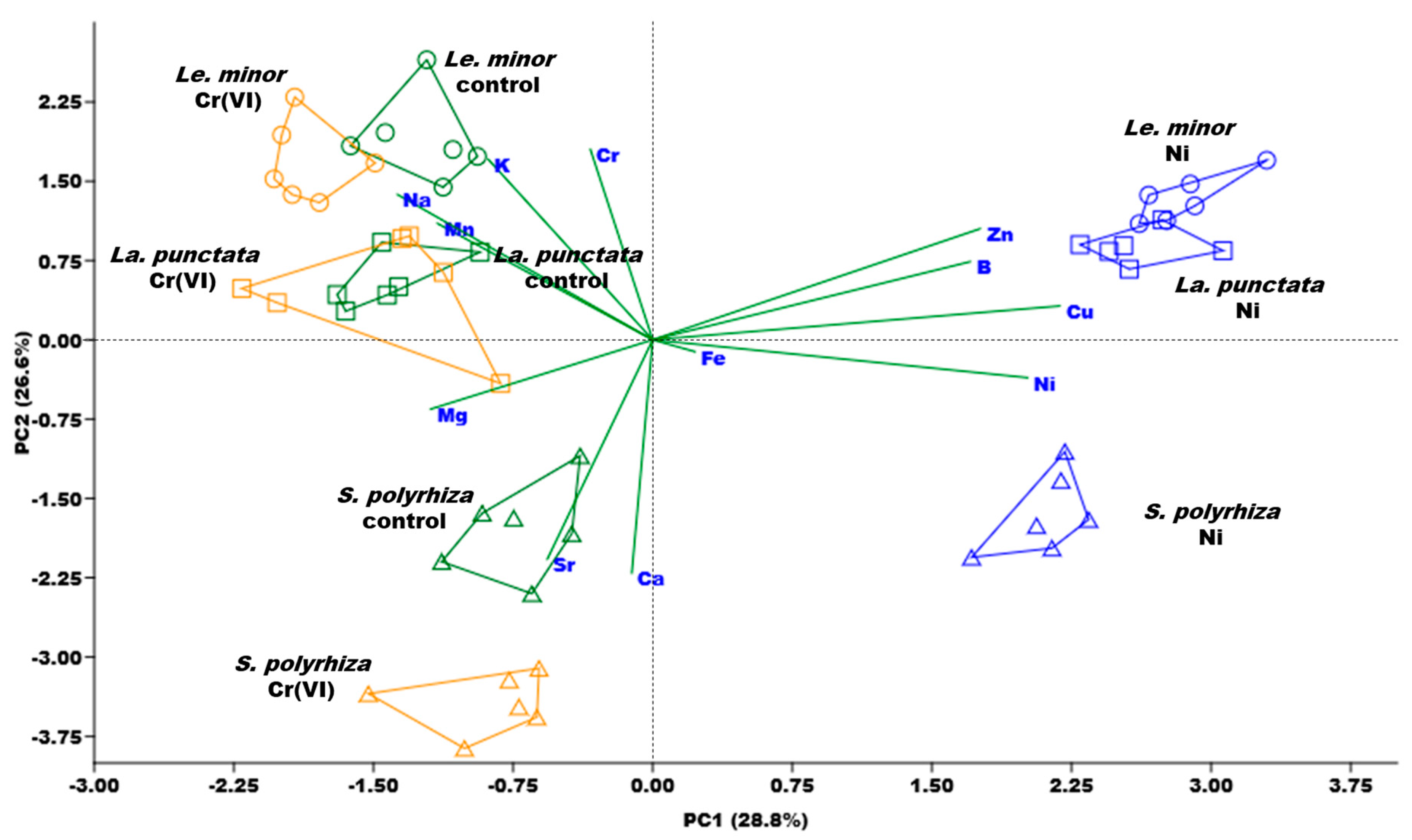
3.3. Biomass Ionomic Composition
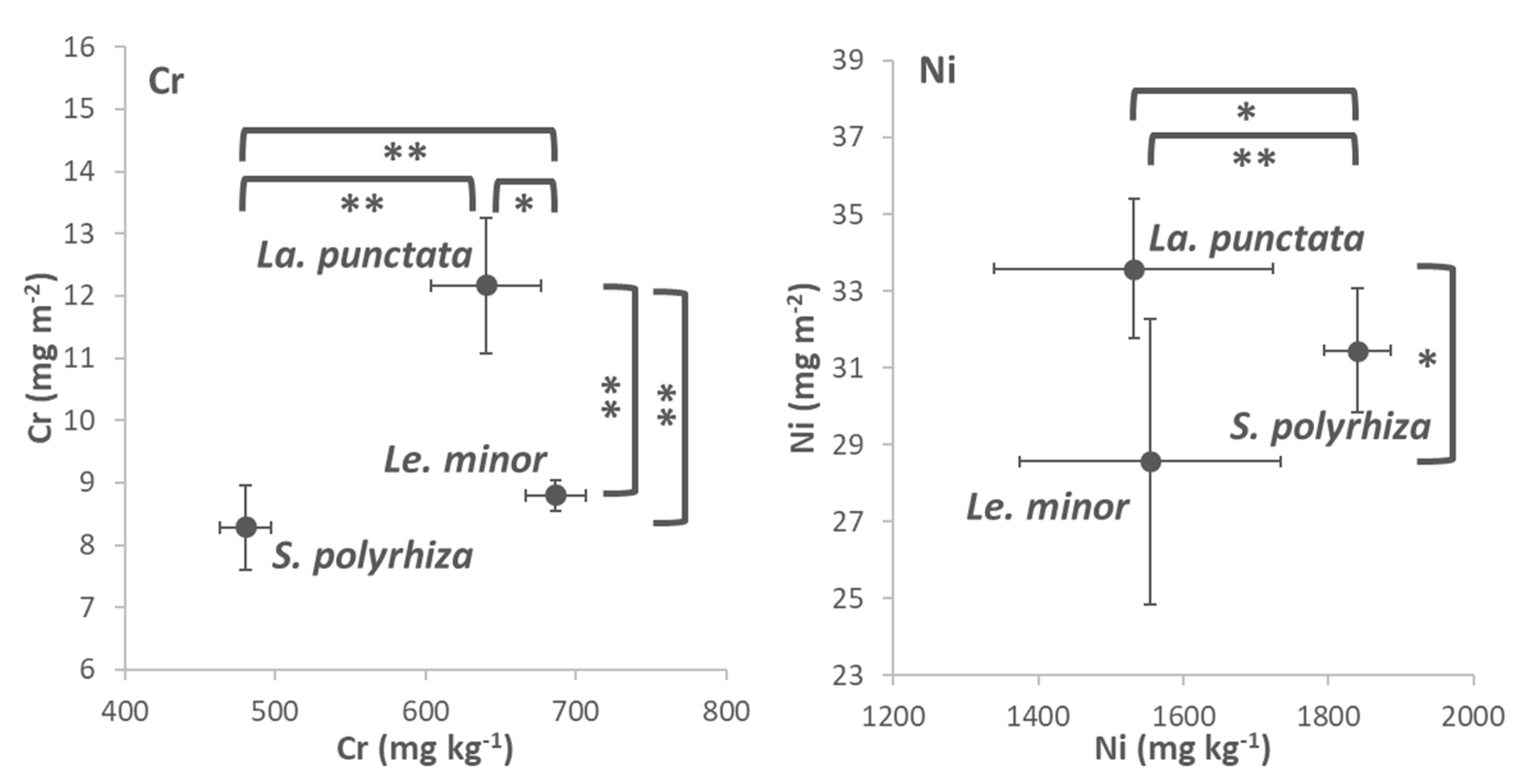
3.4. Cr and Ni Accumulation
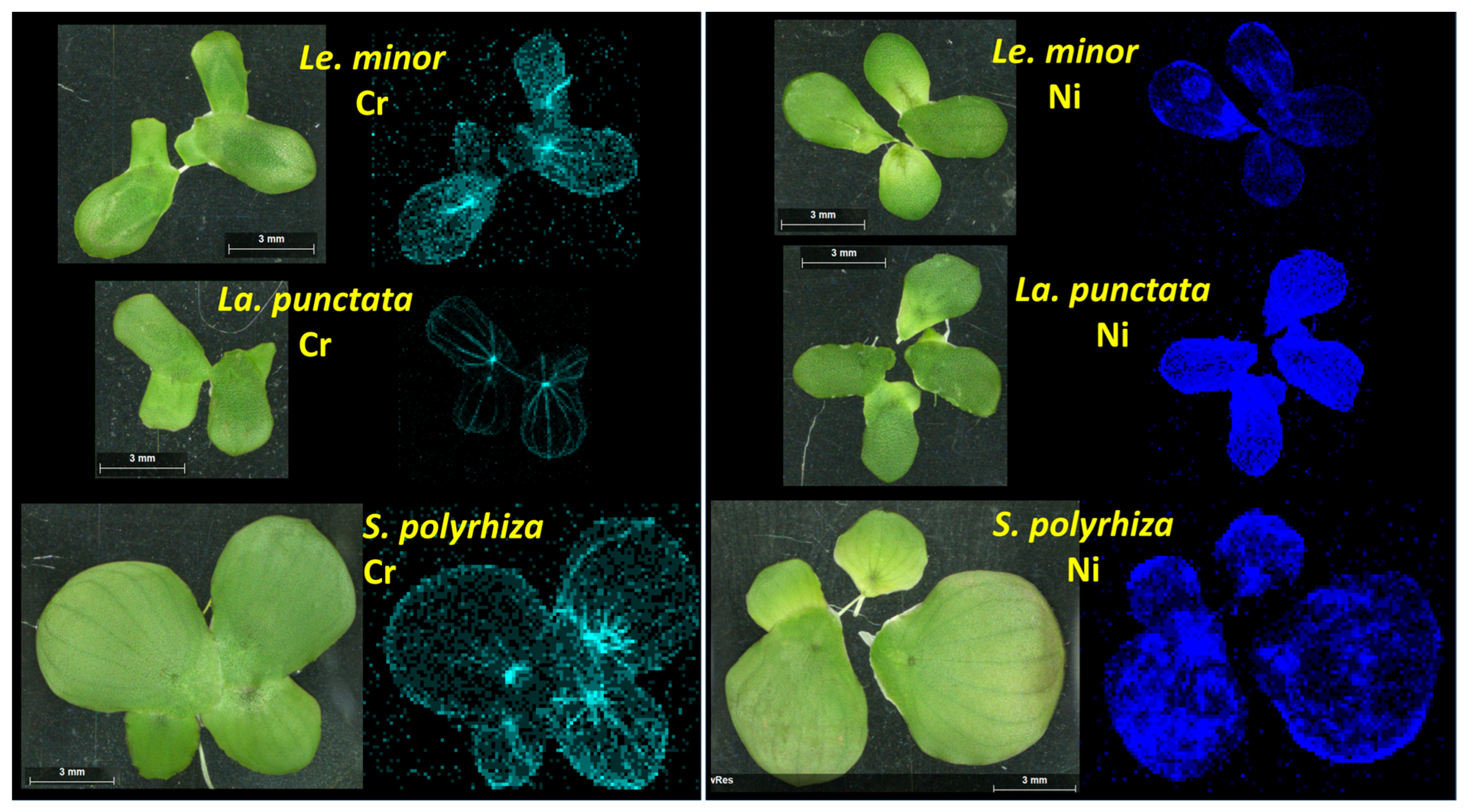
3.5. Within-Frond Distribution of Cr and Ni
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a Model Plant System in the Genomics and Postgenomics Era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef] [PubMed]
- Landolt, E.; Kandeler, R. Biosystematic Investigations in the Family of Duckweeds (Lemnaceae), Vol. 4: The Family of Lemnaceae-a Monographic Study, Vol. 2 (Phytochemistry, Physiology, Application, Bibliography); Geobotanisches Institut der Eidgenoessischen Technischen Hochschule, Stiftung Ruebel: Zürich, Switzerland, 1987. [Google Scholar]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.-J. Relative in Vitro Growth Rates of Duckweeds (Lemnaceae)-the Most Rapidly Growing Higher Plants. Plant Biol. J. 2015, 17, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Paolacci, S.; Stejskal, V.; Jansen, M.A.K. Estimation of the Potential of Lemna Minor for Effluent Remediation in Integrated Multi-Trophic Aquaculture Using Newly Developed Synthetic Aquaculture Wastewater. Aquac. Int. 2021, 29, 2101–2118. [Google Scholar] [CrossRef]
- Ziegler, P.; Sree, K.S.; Appenroth, K.-J. Duckweeds for Water Remediation and Toxicity Testing. Toxicol. Environ. Chem. 2016, 98, 1127–1154. [Google Scholar] [CrossRef]
- Pagliuso, D.; Grandis, A.; Fortirer, J.S.; Camargo, P.; Floh, E.I.S.; Buckeridge, M.S. Duckweeds as Promising Food Feedstocks Globally. Agronomy 2022, 12, 796. [Google Scholar] [CrossRef]
- Petersen, F.; Demann, J.; Restemeyer, D.; Olfs, H.-W.; Westendarp, H.; Appenroth, K.-J.; Ulbrich, A. Influence of Light Intensity and Spectrum on Duckweed Growth and Proteins in a Small-Scale, Re-Circulating Indoor Vertical Farm. Plants 2022, 11, 1010. [Google Scholar] [CrossRef]
- Abramson, B.W.; Novotny, M.; Hartwick, N.T.; Colt, K.; Aevermann, B.D.; Scheuermann, R.H.; Michael, T.P. The Genome and Preliminary Single-Nuclei Transcriptome of Lemna minuta Reveals Mechanisms of Invasiveness. Plant Physiol. 2022, 188, 879–897. [Google Scholar] [CrossRef]
- Ater, M.; Ali, N.; Kasmi, H. Tolerance and accumulation of copper and chromium in two duckweed species: Lemna minor L. and Lemna gibba L. Rev. Sci. L'eau/J. Water Sci. 2006, 19, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Bokhari, S.H.; Mahmood-Ul-Hassan, M.; Ahmad, M. Phytoextraction of Ni, Pb and, Cd by Duckweeds. Int. J. Phytoremediation 2019, 21, 799–806. [Google Scholar] [CrossRef]
- Khellaf, N.; Zerdaoui, M. Phytoaccumulation of Zinc Using the Duckweed Lemna gibba L.: Effect of Temperature, PH and Metal Source. Desalination Water Treat. 2013, 51, 5755–5760. [Google Scholar] [CrossRef]
- Mkandawire, M.; Taubert, B.; Dudel, E.G. Capacity of Lemna gibba L. (Duckweed) for Uranium and Arsenic Phytoremediation in Mine Tailing Waters. Int. J. Phytoremediation 2004, 6, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhang, H.; Wang, Q.; Shao, M.; Li, X.; Chen, D.; Zeng, R.; Song, Y. Intraspecific Variations in Cadmium Tolerance and Phytoaccumulation in Giant Duckweed (Spirodela polyrhiza). J. Hazard. Mater. 2020, 395, 122672. [Google Scholar] [CrossRef]
- Sree, K.S.; Adelmann, K.; Garcia, C.; Lam, E.; Appenroth, K.-J. Natural Variance in Salt Tolerance and Induction of Starch Accumulation in Duckweeds. Planta 2015, 241, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Bai, T.; Kishchenko, O. Potential of Lemnoideae species for phytoremediation of fresh water with elevated manganese concentration. Innov. Biosyst. Bioeng. 2019, 3, 232–238. [Google Scholar] [CrossRef] [Green Version]
- van der Spiegel, M.; Noordam, M.y.; van der Fels-Klerx, H.j. Safety of Novel Protein Sources (Insects, Microalgae, Seaweed, Duckweed, and Rapeseed) and Legislative Aspects for Their Application in Food and Feed Production. Compr. Rev. Food Sci. Food Saf. 2013, 12, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Salt, D.E.; Baxter, I.; Lahner, B. Ionomics and the Study of the Plant Ionome. Annu. Rev. Plant Biol. 2008, 59, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Demmig-Adams, B.; López-Pozo, M.; Polutchko, S.K.; Fourounjian, P.; Stewart, J.J.; Zenir, M.C.; Adams, W.W. Growth and Nutritional Quality of Lemnaceae Viewed Comparatively in an Ecological and Evolutionary Context. Plants 2022, 11, 145. [Google Scholar] [CrossRef]
- Kufel, L.; Strzałek, M.; Przetakiewicz, A. Plant Response to Overcrowding–Lemna minor Example. Acta Oecologica 2018, 91, 73–80. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Palme, K.J.; Jansen, M.A.K. Stress-Induced Morphogenic Responses: Growing out of Trouble? Trends Plant Sci. 2007, 12, 98–105. [Google Scholar] [CrossRef]
- Szabó, S.; Koleszár, G.; Braun, M.; Nagy, Z.; Vicei, T.T.; Peeters, E.T.H.M. Submerged Rootless Macrophytes Sustain a Stable State against Free-Floating Plants. Ecosystems 2022, 25, 17–29. [Google Scholar] [CrossRef]
- Szabó, S.; Koleszár, G.; Zavanyi, G.; Nagy, P.T.; Braun, M.; Hilt, S. Disentangling the Mechanisms Sustaining a Stable State of Submerged Macrophyte Dominance against Free-Floating Competitors. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Reale, L.; Ferranti, F.; Mantilacci, S.; Corboli, M.; Aversa, S.; Landucci, F.; Baldisserotto, C.; Ferroni, L.; Pancaldi, S.; Venanzoni, R. Cyto-Histological and Morpho-Physiological Responses of Common Duckweed (Lemna minor L.) to Chromium. Chemosphere 2016, 145, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Appenroth, K.-J.; Ziegler, P.; Sree, K.S. Accumulation of Starch in Duckweeds (Lemnaceae), Potential Energy Plants. Physiol. Mol. Biol. Plants 2021, 27, 2621–2633. [Google Scholar] [CrossRef] [PubMed]
- Kocjan, G.; Samardakiewicz, S.; Woźny, A. Regions of Lead Uptake in Lemna minor Plants and Localization of This Metal within Selected Parts of the Root. Biol. Plant. 1996, 38, 107–117. [Google Scholar] [CrossRef]
- Yue, L.; Zhao, J.; Yu, X.; Lv, K.; Wang, Z.; Xing, B. Interaction of CuO Nanoparticles with Duckweed (Lemna minor L.): Uptake, Distribution and ROS Production Sites. Environ. Pollut. 2018, 243, 543–552. [Google Scholar] [CrossRef]
- Xu, H.; Yu, C.; Xia, X.; Li, M.; Li, H.; Wang, Y.; Wang, S.; Wang, C.; Ma, Y.; Zhou, G. Comparative Transcriptome Analysis of Duckweed (Landoltia punctata) in Response to Cadmium Provides Insights into Molecular Mechanisms Underlying Hyperaccumulation. Chemosphere 2018, 190, 154–165. [Google Scholar] [CrossRef]
- Kanngießer, B.; Malzer, W.; Pagels, M.; Lühl, L.; Weseloh, G. Three-Dimensional Micro-XRF under Cryogenic Conditions: A Pilot Experiment for Spatially Resolved Trace Analysis in Biological Specimens. Anal. Bioanal. Chem. 2007, 389, 1171–1176. [Google Scholar] [CrossRef]
- Morales-Barrera, L.; Flores-Ortiz, C.M.; Cristiani-Urbina, E. Single and Binary Equilibrium Studies for Ni2+ and Zn2+ Biosorption onto Lemna gibba from Aqueous Solutions. Processes 2020, 8, 1089. [Google Scholar] [CrossRef]
- Environment Canada. Biological Test Method-Test for Measuring the Inhibition of Growth Using the Freshwater Macrophyte Lemna minor; Method Development and Applications Section, Environmental Technology Centre, Environment Canada: Ottawa, ON, Canada, 2007. [Google Scholar]
- Oláh, V.; Hepp, A.; Irfan, M.; Mészáros, I. Chlorophyll Fluorescence Imaging-Based Duckweed Phenotyping to Assess Acute Phytotoxic Effects. Plants 2021, 10, 2763. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 32–64. [Google Scholar]
- OECD. Guidelines for the Testing of Chemicals, Revised Proposal for a New Guideline 221, Lemna Sp. Growth Inhibition Test; OECD: Paris, France, 2006. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Lahive, E.; Halloran, J.O.; Jansen, M.A.K. Differential Sensitivity of Four Lemnaceae Species to Zinc Sulphate. Environ. Exp. Bot. 2011, 71, 25–33. [Google Scholar] [CrossRef]
- Oláh, V.; Lakatos, G.; Bertók, C.; Kanalas, P.; Szőllősi, E.; Kis, J.; Mészáros, I. Short-Term Chromium(VI) Stress Induces Different Photosynthetic Responses in Two Duckweed Species, Lemna gibba L. and Lemna minor L. Photosynthetica 2010, 48, 513–520. [Google Scholar] [CrossRef]
- Xyländer, M.; Augsten, H. Different Sensitivity of Some Lemnaceae to Nickel. Beiträge Zur Biol. Der Pflanz. 1992, 67, 89–99. [Google Scholar]
- Appenroth, K.-J.; Krech, K.; Keresztes, Á.; Fischer, W.; Koloczek, H. Effects of Nickel on the Chloroplasts of the Duckweeds Spirodela polyrhiza and Lemna minor and Their Possible Use in Biomonitoring and Phytoremediation. Chemosphere 2010, 78, 216–223. [Google Scholar] [CrossRef]
- Yang, J.; Li, G.; Bishopp, A.; Heenatigala, P.P.M.; Hu, S.; Chen, Y.; Wu, Z.; Kumar, S.; Duan, P.; Yao, L.; et al. A Comparison of Growth on Mercuric Chloride for Three Lemnaceae Species Reveals Differences in Growth Dynamics That Effect Their Suitability for Use in Either Monitoring or Remediating Ecosystems Contaminated with Mercury. Front. Chem. 2018, 6, 112. [Google Scholar] [CrossRef]
- Shao, J.; Liu, Z.; Ding, Y.; Wang, J.; Li, X.; Yang, Y. Biosynthesis of the Starch Is Improved by the Supplement of Nickel (Ni2+) in Duckweed (Landoltia punctata). J. Plant Res. 2020, 133, 587–596. [Google Scholar] [CrossRef]
- Liu, Y.; Sanguanphun, T.; Yuan, W.; Cheng, J.J.; Meetam, M. The Biological Responses and Metal Phytoaccumulation of Duckweed Spirodela polyrhiza to Manganese and Chromium. Environ. Sci. Pollut. Res. 2017, 24, 19104–19113. [Google Scholar] [CrossRef]
- Taylor, G.J.; Stadt, K.J. Interactive Effects of Cadmium, Copper, Manganese, Nickel, and Zinc on Root Growth of Wheat (Triticum aestivum) in Solution Culture. In Plant Nutrition—Physiology and Applications: Proceedings of the Eleventh International Plant Nutrition Colloquium, Wageningen, The Netherlands, 30 July–4 August 1989; van Beusichem, M.L., Ed.; Developments in Plant and Soil Sciences; Springer: Dordrecht, The Netherlands, 1990; pp. 317–322. ISBN 978-94-009-0585-6. [Google Scholar]
- van der Ent, A.; Pillon, Y.; Fogliani, B.; Gei, V.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; Spiers, K.M.; Paul, A.L.D.; Isnard, S. Contrasting Nickel and Manganese Accumulation and Localization in New Caledonian Cunoniaceae. Plant Soil 2022, 475, 515–534. [Google Scholar] [CrossRef]
- Park, J.H. Contrasting Effects of Cr(III) and Cr(VI) on Lettuce Grown in Hydroponics and Soil: Chromium and Manganese Speciation. Environ. Pollut. 2020, 266, 115073. [Google Scholar] [CrossRef]
- Dube, B.K.; Tewari, K.; Chatterjee, J.; Chatterjee, C. Excess Chromium Alters Uptake and Translocation of Certain Nutrients in Citrullus. Chemosphere 2003, 53, 1147–1153. [Google Scholar] [CrossRef]
- Turner, M.A.; Rust, R.H. Effects of Chromium on Growth and Mineral Nutrition of Soybeans. Soil Sci. Soc. Am. J. 1971, 35, 755–758. [Google Scholar] [CrossRef]
- Dalir, N.; Tandy, S.; Gramlich, A.; Khoshgoftarmanesh, A.; Schulin, R. Effects of Nickel on Zinc Uptake and Translocation in Two Wheat Cultivars Differing in Zinc Efficiency. Environ. Exp. Bot. 2017, 134, 96–101. [Google Scholar] [CrossRef]
- Pillon, Y.; Petit, D.; Gady, C.; Soubrand, M.; Joussein, E.; Saladin, G. Ionomics Suggests Niche Differences between Sympatric Heathers (Ericaceae). Plant Soil 2019, 434, 481–489. [Google Scholar] [CrossRef]
- Jovanović, P.; Rachmilevitch, S.; Roitman, N.; Erel, R. Strontium as a Tracer for Calcium: Uptake, Transport and Partitioning within Tomato Plants. Plant Soil 2021, 466, 303–316. [Google Scholar] [CrossRef]
- Ali, Z.; Waheed, H.; Kazi, A.G.; Hayat, A.; Ahmad, M. Chapter 16-Duckweed: An Efficient Hyperaccumulator of Heavy Metals in Water Bodies. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 411–429. ISBN 978-0-12-803158-2. [Google Scholar]
- Dirilgen, N.; İnel, Y. Effects of Zinc and Copper on Growth and Metal Accumulation in Duckweed, Lemna minor. Bull. Environ. Contam. Toxicol. 1994, 53, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Khellaf, N.; Zerdaoui, M. Growth Response of the Duckweed Lemna gibba L. to Copper and Nickel Phytoaccumulation. Ecotoxicology 2010, 19, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Devlamynck, R.; de Souza, M.F.; Leenknegt, J.; Jacxsens, L.; Eeckhout, M.; Meers, E. Lemna minor Cultivation for Treating Swine Manure and Providing Micronutrients for Animal Feed. Plants 2021, 10, 1124. [Google Scholar] [CrossRef] [PubMed]
- McGeer, J.C.; Brix, K.V.; Skeaff, J.M.; DeForest, D.K.; Brigham, S.I.; Adams, W.J.; Green, A. Inverse Relationship between Bioconcentration Factor and Exposure Concentration for Metals: Implications for Hazard Assessment of Metals in the Aquatic Environment. Environ. Toxicol. Chem. 2003, 22, 1017–1037. [Google Scholar] [CrossRef]
- Regoli, L.; Van Tilborg, W.; Heijerick, D.; Stubblefield, W.; Carey, S. The Bioconcentration and Bioaccumulation Factors for Molybdenum in the Aquatic Environment from Natural Environmental Concentrations up to the Toxicity Boundary. Sci. Total Environ. 2012, 435–436, 96–106. [Google Scholar] [CrossRef]
- Kalčíková, G.; Zupančič, M.; Jemec, A.; Žgajnar Gotvajn, A. The Impact of Humic Acid on Chromium Phytoextraction by Aquatic Macrophyte Lemn minor. Chemosphere 2016, 147, 311–317. [Google Scholar] [CrossRef]
- Uysal, Y. Removal of Chromium Ions from Wastewater by Duckweed, Lemna minor L. by Using a Pilot System with Continuous Flow. J. Hazard. Mater. 2013, 263, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, D.D.; Parlak, K.U. Nickel-Induced Changes in Lipid Peroxidation, Antioxidative Enzymes, and Metal Accumulation in Lemna gibba. Int. J. Phytoremediation 2011, 13, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Walsh, É.; Paolacci, S.; Burnell, G.; Jansen, M.A.K. The Importance of the Calcium-to-Magnesium Ratio for Phytoremediation of Dairy Industry Wastewater Using the Aquatic Plant Lemna minor L. Int. J. Phytoremediation 2020, 22, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Nagasawa, H.; Yamashiro, N.; Yasuno, N.; Watanabe, T.; Kitazawa, H.; Takano, S.; Tokuji, Y.; Tani, M.; Takamure, I.; et al. Rice RCN1/OsABCG5 Mutation Alters Accumulation of Essential and Nonessential Minerals and Causes a High Na/K Ratio, Resulting in a Salt-Sensitive Phenotype. Plant Sci. 2014, 224, 103–111. [Google Scholar] [CrossRef]
- Walsh, É.; Coughlan, N.E.; O’Brien, S.; Jansen, M.A.K.; Kuehnhold, H. Density Dependence Influences the Efficacy of Wastewater Remediation by Lemn minor. Plants 2021, 10, 1366. [Google Scholar] [CrossRef]
- Sharma, D.C.; Sharma, C.P.; Tripathi, R.D. Phytotoxic Lesions of Chromium in Maize. Chemosphere 2003, 51, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K. Effect of Trivalent and Hexavalent Chromium on Spinach (Spinacea oleracea L.). Environ. Ecol. 2001, 19, 807–810. [Google Scholar]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Aamer, M.; Nawaz, M.; Ali, A.; Khan, M.A.U.; Khan, T.A. Nickel Toxicity in Plants: Reasons, Toxic Effects, Tolerance Mechanisms, and Remediation Possibilities—A Review. Environ. Sci. Pollut. Res. 2019, 26, 12673–12688. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Cr(VI) | Ni | |
|---|---|---|---|
| Le. minor | 6.70 ± 0.53 Bc | 8.33 ± 0.19 Cb | 11.79 ± 0.24 Aa |
| La. punctata | 7.26 ± 0.24 Bb | 9.11 ± 0.34 Ba | 10.02 ± 0.35 Ba |
| S. polyrhiza | 8.36 ± 1.01 Ac | 11.78 ± 0.96 Ab | 13.20 ± 1.09 Aa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oláh, V.; Irfan, M.; Szabó, Z.B.; Sajtos, Z.; Ragyák, Á.Z.; Döncző, B.; Jansen, M.A.K.; Szabó, S.; Mészáros, I. Species- and Metal-Specific Responses of the Ionome of Three Duckweed Species under Chromate and Nickel Treatments. Plants 2023, 12, 180. https://doi.org/10.3390/plants12010180
Oláh V, Irfan M, Szabó ZB, Sajtos Z, Ragyák ÁZ, Döncző B, Jansen MAK, Szabó S, Mészáros I. Species- and Metal-Specific Responses of the Ionome of Three Duckweed Species under Chromate and Nickel Treatments. Plants. 2023; 12(1):180. https://doi.org/10.3390/plants12010180
Chicago/Turabian StyleOláh, Viktor, Muhammad Irfan, Zsuzsanna Barnáné Szabó, Zsófi Sajtos, Ágota Zsófia Ragyák, Boglárka Döncző, Marcel A. K. Jansen, Sándor Szabó, and Ilona Mészáros. 2023. "Species- and Metal-Specific Responses of the Ionome of Three Duckweed Species under Chromate and Nickel Treatments" Plants 12, no. 1: 180. https://doi.org/10.3390/plants12010180