
Floral Scent Chemistry and Pollinators of a Sexually Dimorphic Neotropical Orchid

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
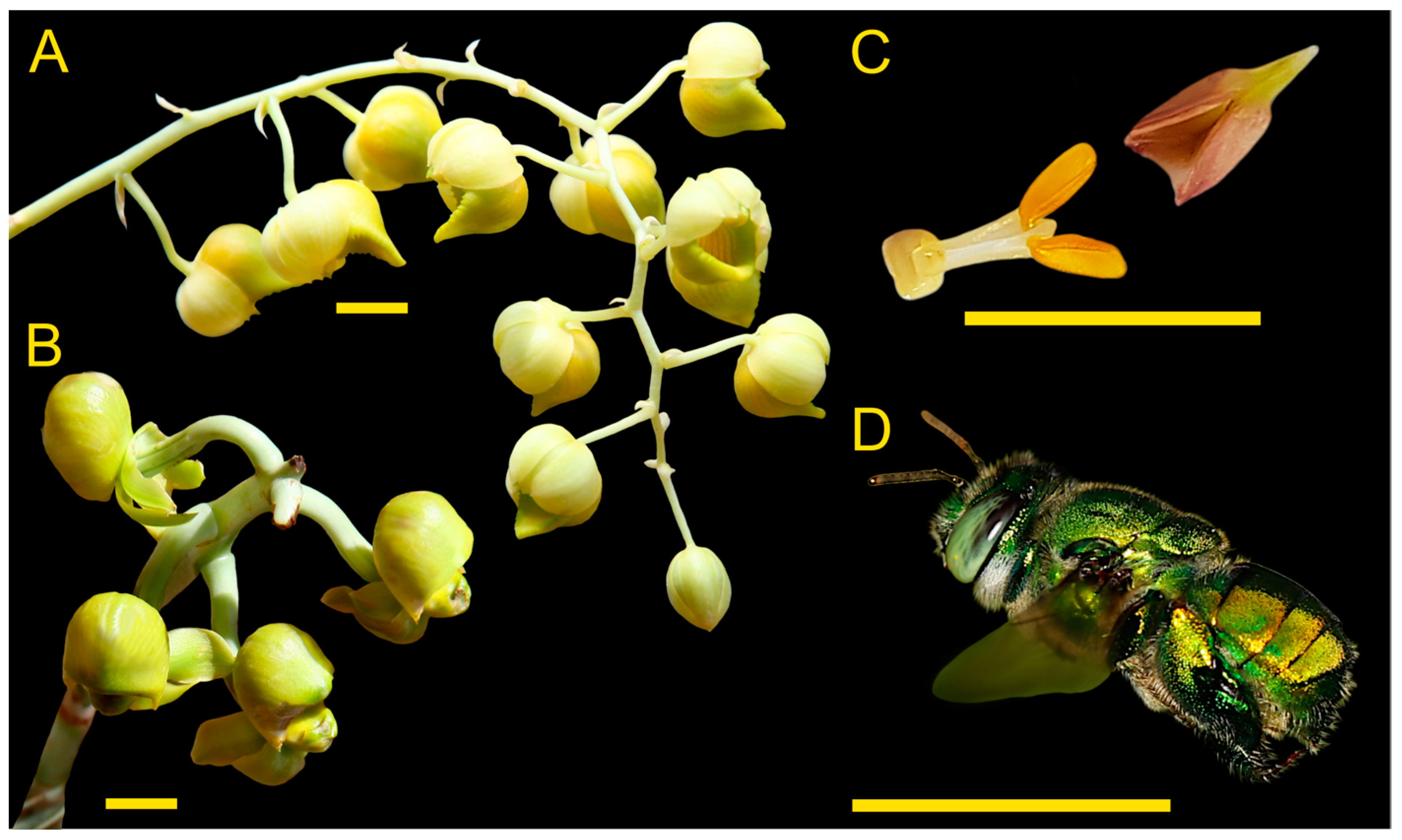
2.1. Observation of Floral Visitors
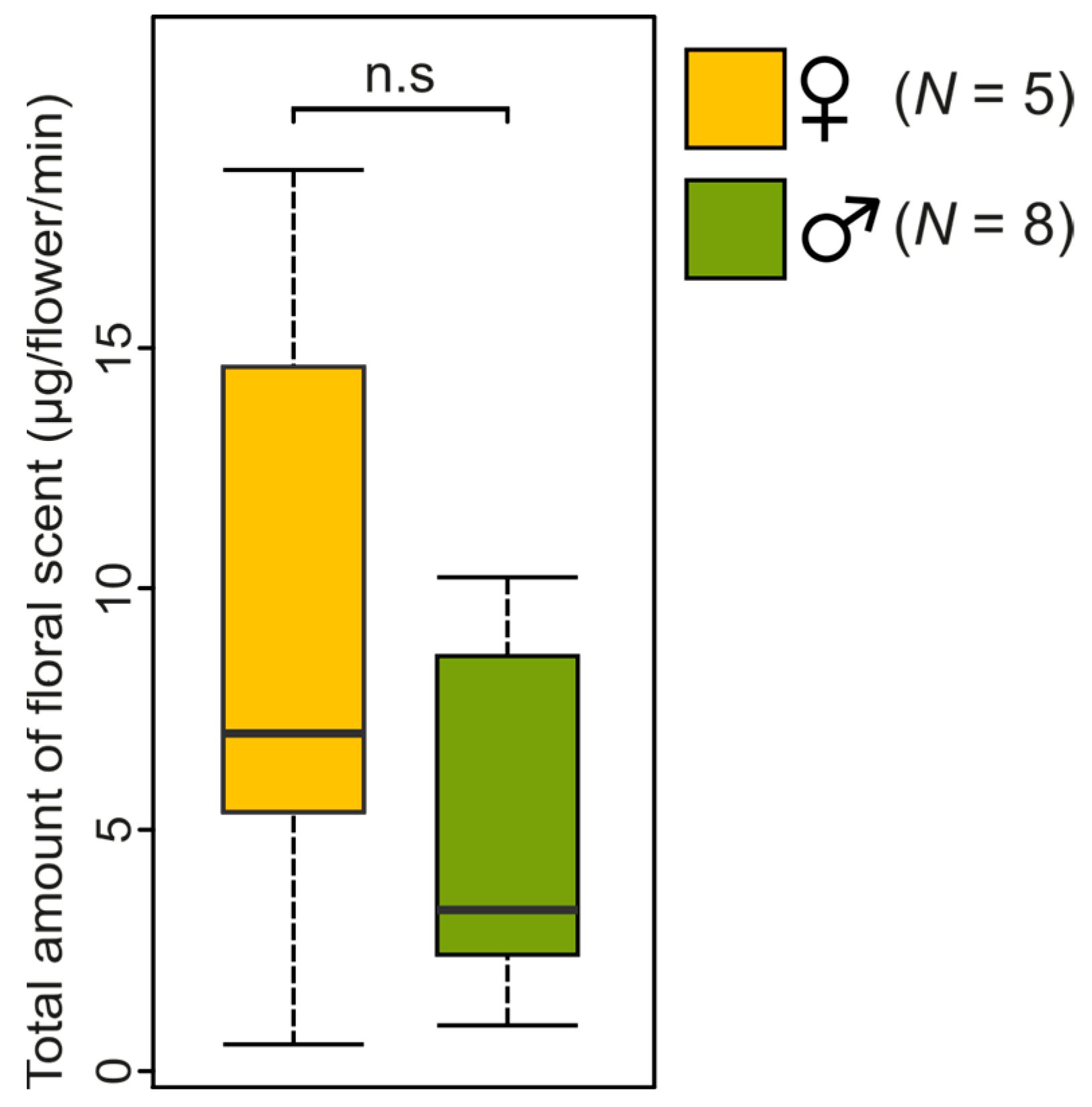
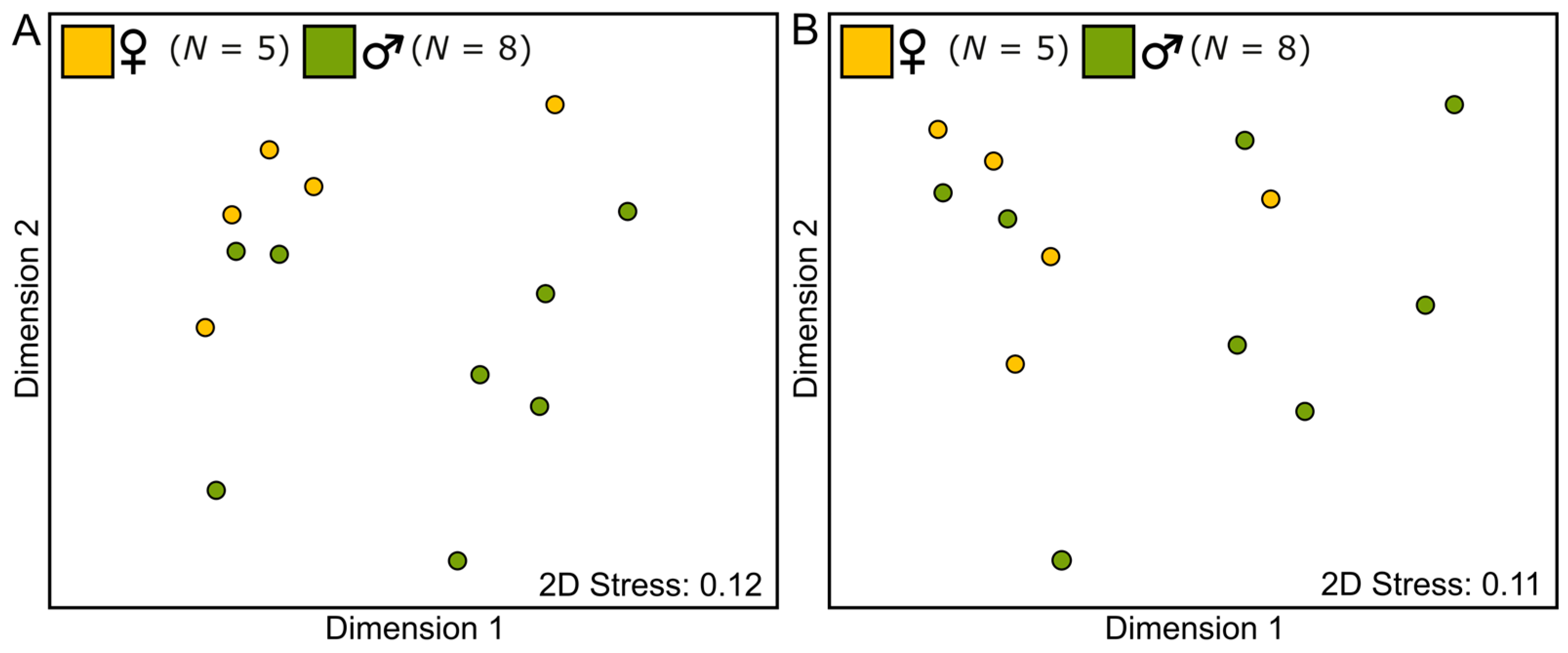
2.2. Characterisation of the Floral Scent Traits of Catasetum maranhense
3. Materials and Methods
3.1. Observation of Floral Visitors
3.2. Sampling of Floral Scents
3.3. Chemical Analysis
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roubik, D.; Hanson, P. Orchid Bees of Tropical America: Biology and Field Guide; INBio Pres.: Heredia, Costa Rica, 2004. [Google Scholar]
- Vogel, S. Parfümsammelnde Bienen Als Bestäuber von Orchidaceen Und Gloxinia. Österr. Bot. Z 1966, 113, 302–361. [Google Scholar] [CrossRef]
- Ramírez, S.R. Orchid Bees. Curr. Biol. 2009, 19, R1061–R1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemésio, A. Orchid Bees (Hymenoptera: Apidae) of the Brazilian Atlantic Forest. Zootaxa 2009, 2041, 1–242. [Google Scholar] [CrossRef] [Green Version]
- Dressler, R.L. Biology of the Orchid Bees (Euglossini). Annu. Rev. Ecol. Syst. 1982, 13, 373–394. [Google Scholar] [CrossRef]
- Eltz, T.; Whitten, W.M.; Roubik, D.W.; Linsenmair, K.E. Fragrance Collection, Storage, and Accumulation by Individual Male Orchid Bees. J. Chem. Ecol. 1999, 25, 157–176. [Google Scholar] [CrossRef]
- Eltz, T.; Roubik, D.W.; Whitten, M.W. Fragrances, Male Display and Mating Behaviour of Euglossa hemichlora: A Flight Cage Experiment. Physiol. Entomol. 2003, 28, 251–260. [Google Scholar] [CrossRef]
- Petini-Benelli, A. Orquídeas de Mato Grosso: Genus Catasetum, L.C. Rich Ex Kunth; PoD Editora: Rio de Janeiro, Brazil, 2012; ISBN 9788582250068. [Google Scholar]
- Romero, G.A.; Jenny, R. Contributions towards a Monograph of Catasetum (Catasetinae, Orchidaceae) I: A Checklist of Species, Varieties, and Natural Hybrids. Harvard Pap. Bot. 1993, 9, 59–84. [Google Scholar]
- Milet-Pinheiro, P.; Gerlach, G. Biology of the Neotropical Orchid Genus Catasetum: A Historical Review on Floral Scent Chemistry and Pollinators. Perspect. Plant Ecol. Evol. Syst. 2017, 27, 23–34. [Google Scholar] [CrossRef]
- Pijl, L.; Dodson, H. Orchid Flowers. Their Pollination and Evolution; Fairchild Tropical Garden and the University of Miami Press: Coral Gables, FL, USA, 1966; ISBN 9780870240690. [Google Scholar]
- Dodson, C.H. Pollination and Variation in the Subtribe Catasetinae (Orchidaceae). Ann. Missouri Bot. Gard. 1962, 49, 35. [Google Scholar] [CrossRef]
- Romero, G.A.; Nelson, C.E. Sexual Dimorphism in Catasetum Orchids: Forcible Pollen Emplacement and Male Flower Competition. Science 1986, 232, 1538–1540. [Google Scholar] [CrossRef]
- Eltz, T.; Roubik, D.W.; Lunau, K. Experience-Dependent Choices Ensure Species-Specific Fragrance Accumulation in Male Orchid Bees. Behav. Ecol. Sociobiol. 2005, 59, 149–156. [Google Scholar] [CrossRef]
- Ashman, T.L. Sniffing out Patterns of Sexual Dimorphism in Floral Scent. Funct. Ecol. 2009, 23, 852–862. [Google Scholar] [CrossRef]
- Delle-Vedove, R.; Schatz, B.; Dufay, M. Understanding Intraspecific Variation of Floral Scent in Light of Evolutionary Ecology. Ann. Bot. 2017, 120, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, K.; Machado, I.C.; do Amaral Ferraz Navarro, D.M.; Dötterl, S.; Ayasse, M.; Milet-Pinheiro, P. Sexual Dimorphism in Floral Scents of the Neotropical Orchid Catasetum arietinum and Its Possible Ecological and Evolutionary Significance. AoB Plants 2020, 12, 30. [Google Scholar] [CrossRef]
- Milet-Pinheiro, P.; Do Amaral Ferraz Navarro, D.M.; Dötterl, S.; Carvalho, A.T.; Pinto, C.E.; Ayasse, M.; Schlindwein, C. Pollination Biology in the Dioecious Orchid Catasetum uncatum: How Does Floral Scent Influence the Behaviour of Pollinators? Phytochemistry 2015, 116, 149–161. [Google Scholar] [CrossRef]
- Petini-Benelli, A. Catasetum. Available online: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB20005 (accessed on 8 February 2022).
- Gerlach, G.; Schill, R. Composition of Orchid Scents Attracting Euglossine Bees. Bot. Acta 1991, 104, 379–384. [Google Scholar] [CrossRef]
- Williams, N.H.; Whitten, W.M. Orchid Floral Fragrances and Male Euglossine Bees: Methods and Advances in the Last Sesquidecade. Biol. Bull. 1983, 164, 355–395. [Google Scholar] [CrossRef]
- Hetherington-Rauth, M.C.; Ramírez, S.R. Evolutionary Trends and Specialization in the Euglossine Bee–Pollinated Orchid Genus Gongora 1, 2. Ann. Missouri Bot. Gard. 2015, 100, 271–299. [Google Scholar] [CrossRef]
- Milet-Pinheiro, P.; Silva, J.B.F.; Navarro, D.M.A.F.; Machado, I.C.S.; Gerlach, G. Notes on Pollination Ecology and Floral Scent Chemistry of the Rare Neotropical Orchid Catasetum galeritum R Chb.F. Plant Species Biol. 2018, 33, 158–163. [Google Scholar] [CrossRef]
- Ramirez, S.R.; Dressler, R.L.; Ospina, M. Abejas Euglosinas (Hymenoptera: Apidae) de La Región Neotropical: Listado de Especies Con Notas Sobre Su Biología. Biota Colomb. 2002, 3, 7–118. [Google Scholar]
- Rebêlo, J.M.M.; Silva, F.S. Distribuição Das Abelhas Euglossini (Hymenoptera: Apidae) No Estado Do Maranhão, Brasil. An. Soc. Entomológica Bras. 1999, 28, 389–401. [Google Scholar] [CrossRef]
- Dodson, C.H.; Dressler, R.L.; Hills, H.G.; Adams, R.M.; Williams, N.H. Biologically Active Compounds in Orchid Fragrances. Science 1969, 164, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Hills, H.G.; Williams, N.H.; Dodson, C.H. Floral Fragrances and Isolating Mechanisms in the Genus Catasetum (Orchidaceae). Biotropica 1972, 4, 61. [Google Scholar] [CrossRef]
- Barbosa, C.M.; Maia, A.C.D.; Martel, C.; Regueira, J.C.S.; Navarro, D.M.d.A.F.; Raguso, R.A.; Milet-Pinheiro, P.; Machado, I.C. Reproductive Biology of Syagrus coronata (Arecaceae): Sex-Biased Insect Visitation and the Unusual Case of Scent Emission by Peduncular Bracts. Plant Biol. 2021, 23, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Dötterl, S.; Ramírez, S.R.; Etl, F.; Machado, I.C.; do Navarro, D.M.A.F.; Dobler, D.; Reiser, O.; Ayasse, M.; Milet-Pinheiro, P. Unraveling the Olfactory Biases of Male Euglossine Bees: Species-Specific Antennal Responses and Their Evolutionary Significance for Perfume Flowers. Front. Ecol. Evol. 2021, 9, 690. [Google Scholar] [CrossRef]
- Nemésio, A.; Ferrari, R.R. The Species of Eulaema (Eulaema) Lepeletier, 1841 (Hymenoptera: Apidae: Euglossina) from Eastern Brazil, with Description of Eulaema quadragintanovem Sp. N. From the State of Ceará. Zootaxa 2012, 123–132. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2021. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Compounds | RI | Male (% Mean ± SD) | Female (Mean ± SD) |
|---|---|---|---|
| Aromatics | |||
| 1,4-Dimethoxybenzene * | 1168 | 0 ± 0 | Tr |
| Methyl p-anisate * | 1376 | 2.28 ± 2.46 | 0.69 ± 0.56 |
| (E)-Methyl cinnamate * | 1386 | 0.80 ± 0.97 | 1.23 ± 0.59 |
| Ar-curcumene | 1486 | 0.17 ± 0.20 | 0.27 ± 0.36 |
| (Z)-Methyl p-methoxycinnamate | 1592 | 6.07 ± 4.63 | 14.46 ± 9.54 |
| (E)-Methyl p-methoxycinnamate | 1675 | 10.38 ± 11.10 | 18.66 ± 23.97 |
| Monoterpenes | |||
| α-Thujene * | 927 | Tr | 0.07 ± 0.06 |
| α-Pinene * | 932 | 1.46 ± 0.96 | 1.56 ± 1.38 |
| Camphene * | 947 | Tr | Tr |
| Sabinene * | 972 | 0.51 ± 0.73 | 0.90 ± 0.68 |
| β-Pinene * | 974 | 0.03 ± 0.10 | 0.25 ± 0.17 |
| β-Myrcene * | 992 | 3.92 ± 2.13 | 5.52 ± 3.34 |
| Limonene * | 1028 | 1.96 ± 2.88 | 0.97 ± 0.69 |
| Eucalyptol * | 1030 | 53.27 ± 20.71 | 36.64 ± 21.70 |
| (Z)-β-Ocimene * | 1040 | Tr | 0.02 ± 0.04 |
| (E)-β-Ocimene * | 1050 | 0.96 ± 0.94 | 1.11 ± 1.03 |
| γ-Terpinene * | 1058 | Tr | 0.08 ± 0.05 |
| α-Terpinolene * | 1088 | 0.02 ± 0.04 | 0.01 ± 0.02 |
| α-Terpineol * | 1191 | 0.15 ± 0.24 | 0.07 ± 0.17 |
| N-bearing compounds | |||
| Indole * | 1293 | 13.75 ± 11.10 | 13.57 ± 7.08 |
| Sesquiterpenes | |||
| Dihydro-β-ionone * | 1439 | 0.10 ± 0.03 | 0.05 ± 0.09 |
| (Z,Z)-α-Farnesene | 1494 | 0.04 ± 0.07 | 0.04 ± 0.02 |
| (E,E)-α-Farnesene | 1507 | 3.29 ± 2.42 | 3.40 ± 2.57 |
| Sesquicineole | 1516 | 0 ± 0 | 0.06 ± 0.10 |
| 5 unknown compounds | 0.81 ±1.51 | 0.24 ± 0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milet-Pinheiro, P.; Pinto, C.E.; Navarro, D.M.A.F.; Silva, J.B.F.; Brandt, K.; Ayasse, M. Floral Scent Chemistry and Pollinators of a Sexually Dimorphic Neotropical Orchid. Plants 2023, 12, 17. https://doi.org/10.3390/plants12010017
Milet-Pinheiro P, Pinto CE, Navarro DMAF, Silva JBF, Brandt K, Ayasse M. Floral Scent Chemistry and Pollinators of a Sexually Dimorphic Neotropical Orchid. Plants. 2023; 12(1):17. https://doi.org/10.3390/plants12010017
Chicago/Turabian StyleMilet-Pinheiro, Paulo, Carlos E. Pinto, Daniela M. A. F. Navarro, João B. F. Silva, Katharina Brandt, and Manfred Ayasse. 2023. "Floral Scent Chemistry and Pollinators of a Sexually Dimorphic Neotropical Orchid" Plants 12, no. 1: 17. https://doi.org/10.3390/plants12010017