
Variations in Grain Traits among Local Rice Varieties Collected More Than Half-Century Ago in Indochinese Countries

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Verification of Collection Passport Data on Local Varieties
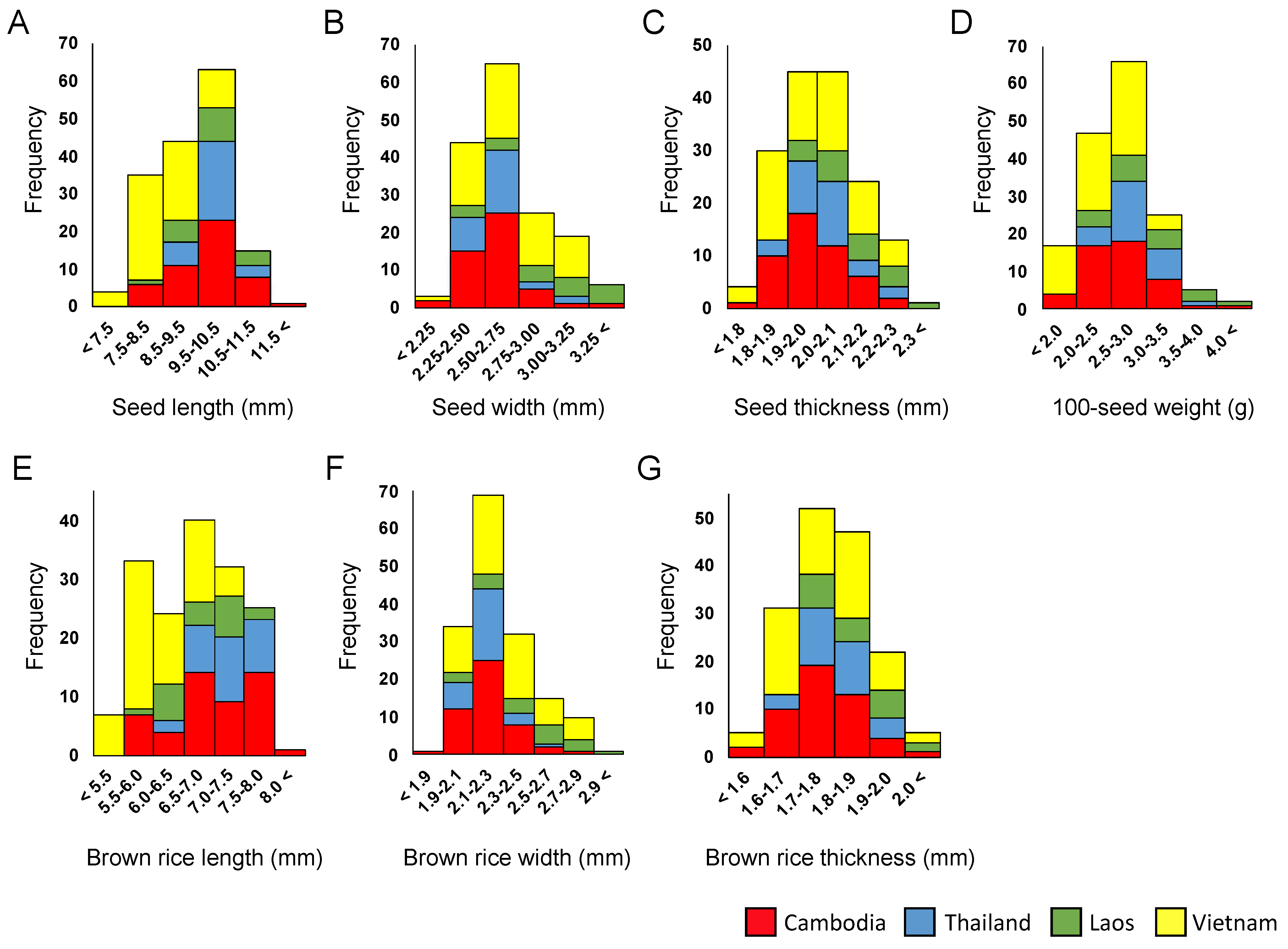
2.2. Grain-Size Variation among Local Varieties
2.3. Detection of Functional Mutations on Seed Size Using PCR-Based Markers
2.4. Evaluation of Other Seed Morphological Traits among Local Varieties
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Grain Morphological Traits
4.3. Detection of Functional Mutations at Grain-Related Loci Using PCR-Based Markers
4.4. Allele Genotyping at Major Loci for Hull Color, Pericarp Color, and Glutinosity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matsumoto, N. Mekong Kikou; Yomiuri Shinbun: Osaka, Japan, 1959; p. 222. (In Japanese) [Google Scholar]
- Hamada, H. Rice in the Mekong Valleys. In Indo-Chinese Studies: Synthetic Research of the Culture of Rice-Cultivating Races in Southeast Asian Counties (I); Matsumoto, N., Ed.; The Japanese Society of Ethnology: Yurindo, Yokohama, Japan, 1965; pp. 505–586. [Google Scholar]
- Song, X.J.; Huang, W.; Shi, M.; Zhu, M.Z.; Lin, H.X. A QTL for Rice Grain Width and Weight Encodes a Previously Unknown RING-Type E3 Ubiquitin Ligase. Nat. Genet. 2007, 39, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Y.X.; Fang, Y.X.; Zeng, L.J.; Xu, J.; Yu, H.P.; Shi, Z.Y.; Pan, J.J.; Zhang, D.; Kang, S.J.; et al. A Rare Allele of GS2 Enhances Grain Size and Grain Yield in Rice. Mol. Plant 2015, 8, 1455–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Han, R.X.; Wu, K.; Zhang, J.Q.; Ye, Y.F.; Wang, S.S.; Chen, J.F.; Pan, Y.J.; Li, Q.; Xu, X.P.; et al. G-protein Beta Gamma Subunits Determine Grain Size Through Interaction with MADS-Domain Transcription Factors in Rice. Nat. Commun. 2018, 9, 852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.H.; Xing, Y.Z.; Mao, H.L.; Lu, T.T.; Han, B.; Xu, C.G.; Li, X.H.; Zhang, Q.F. GS3, a Major QTL for Grain Length and Weight and Minor QTL for Grain Width and Thickness in Rice, Encodes a Putative Transmembrane Protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Lin, Y.S.; Song, X.J.; Shen, J.B.; Huang, W.; Shan, J.X.; Zhu, M.Z.; Jiang, L.W.; Gao, J.P.; Lin, H.X. The Novel Quantitative Trait Locus GL3.1 Controls Rice Grain Size and Yield by Regulating Cyclin-T1;3. Cell Res. 2012, 22, 1666–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, J.Z.; Ma, M.; Bai, C.; Huang, X.H.; Liu, J.L.; Fan, Y.Y.; Song, X.J. TGW3, a Major QTL that Negatively Modulates Grain Length and Weight in Rice. Mol. Plant 2018, 11, 750–753. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.B.; Fan, C.C.; Xing, Y.Z.; Jiang, Y.H.; Luo, L.J.; Sun, L.; Shao, D.; Xu, C.J.; Li, X.H.; Xiao, J.H.; et al. Natural Variation in GS5 Plays an Important Role in Regulating Grain Size and Yield in Rice. Nat. Genet. 2011, 43, 1266–1269. [Google Scholar] [CrossRef]
- Shomura, A.; Izawa, T.; Ebana, K.; Ebitani, T.; Kanegae, H.; Konishi, S.; Yano, M. Deletion in a Gene Associated with Grain Size Increased Yields during Rice Domestication. Nat. Genet. 2008, 40, 1023–1028. [Google Scholar] [CrossRef]
- Duan, P.; Xu, J.; Zeng, D.; Zhang, B.; Geng, M.; Zhang, G.; Huang, K.; Huang, L.; Xu, R.; Ge, S.; et al. Natural Variation in the Promoter of GSE5 Contributes to Grain Size Diversity in Rice. Mol. Plant 2017, 10, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.J.; Li, X.J.; Fu, Y.C.; Zhu, Z.F.; Tan, L.B.; Liu, F.X.; Sun, X.Y.; Sun, X.W.; Sun, C.Q. GS6, a Member of the GRAS Gene Family, Negatively Regulates Grain Size in Rice. J. Integr. Plant Biol. 2013, 55, 938–949. [Google Scholar] [CrossRef]
- Ishimaru, K.; Hirotsu, N.; Madoka, Y.; Murakami, N.; Hara, N.; Onodera, H.; Kashiwagi, T.; Ujiie, K.; Shimizu, B.; Onishi, A.; et al. Loss of Function of the IAA-Glucose Hydrolase Gene TGW6 Enhances Rice Grain Weight and Increases Yield. Nat. Genet. 2013, 45, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Song, X.J.; Kuroha, T.; Ayano, M.; Furuta, T.; Nagai, K.; Komeda, N.; Segami, S.; Miura, K.; Ogawa, D.; Kamura, T.; et al. Rare Allele of a Previously Unidentified Histone H4 Acetyltransferase Enhances Grain Weight, Yield, and Plant Biomass in Rice. Proc. Natl. Acad. Sci. USA 2015, 112, 76–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, L.Z.; Chen, J.Y.; Huang, X.H.; Gong, H.; Luo, J.H.; Hou, Q.Q.; Zhou, T.Y.; Lu, T.T.; Zhu, J.J.; Shangguan, Y.Y.; et al. OsSPL13 Controls Grain Size in Cultivated Rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef]
- Wang, Y.X.; Xiong, G.S.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.X.; Zeng, L.J.; Xu, E.B.; Xu, J.; et al. Copy Number Variation at the GL7 Locus Contributes to Grain Size Diversity in Rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Wu, K.; Yuan, Q.B.; Liu, X.Y.; Liu, Z.B.; Lin, X.Y.; Zeng, R.Z.; Zhu, H.T.; Dong, G.J.; Qian, Q.; et al. Control of Grain Size, Shape and Quality by OsSPL16 in Rice. Nat. Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, B.; Bian, Z.; Li, X.; Zhang, C.; Liu, J.; Li, Q.; Liu, Q.; He, Z. Grain Size Selection Using Novel Functional Markers Targeting 14 Genes in Rice. Rice 2020, 13, 63. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.F.; Si, L.; Wang, Z.; Zhou, Y.; Zhu, J.; Shangguan, Y.; Lu, D.; Fan, D.; Li, C.; Lin, H.; et al. Genetic Control of a Transition from Black to Straw-White Seed Hull in Rice Domestication. Plant Physiol. 2011, 155, 1301–1311. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, M.T.; Thomson, M.J.; Pfeil, B.E.; McCouch, S. Caught Red-Handed: Rc Encodes a Basic Helix-Loop-Helix Protein Conditioning Red Pericarp in Rice. Plant Cell 2006, 18, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Wanchana, S.; Toojinda, T.; Tragoonrung, S.; Vanavichit, A. Duplicated Coding Sequence in the Waxy Allele of Tropical Glutinous Rice (Oryza sativa L.). Plant Sci. 2003, 165, 1193–1199. [Google Scholar] [CrossRef]
- Mao, H.; Sun, S.; Yao, J.; Wang, C.; Yu, S.; Xu, C.; Li, X.; Zhang, Q. Linking Differential Domain Functions of the GS3 Protein to Natural Variation of Grain Size in Rice. Proc. Natl. Acad. Sci. USA 2010, 107, 19579–19584. [Google Scholar] [CrossRef]
- Maeda, H.; Yamaguchi, T.; Omoteno, M.; Takarada, T.; Fujita, K.; Murata, K.; Iyama, Y.; Kojima, Y.; Morikawa, M.; Ozaki, H.; et al. Genetic Dissection of Black Grain Rice by the Development of a Near Isogenic Line. Breed. Sci. 2014, 64, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panaud, O.; Chen, X.; McCouch, S.R. Development of Microsatellite Markers and Characterization of Simple Sequence Length Polymorphism (SSLP) in Rice (Oryza sativa L.). Mol. Gen. Genet. 1996, 252, 597–607. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Trait a | Min. | Max. | Ave. | Ave. | |||
|---|---|---|---|---|---|---|---|
| Cambodia | Thailand | Laos | Vietnam | ||||
| SL (mm) | 7.15 | 11.52 | 9.32 | 9.69 | 9.93 | 9.72 | 8.62 |
| SW (mm) | 2.20 | 3.46 | 2.68 | 2.58 | 2.58 | 2.97 | 2.71 |
| ST (mm) | 1.75 | 2.30 | 2.01 | 1.99 | 2.03 | 2.11 | 1.99 |
| BL (mm) | 5.14 | 8.27 | 6.69 | 6.99 | 7.23 | 6.83 | 6.15 |
| BW (mm) | 1.77 | 2.90 | 2.27 | 2.19 | 2.18 | 2.46 | 2.32 |
| BT (mm) | 1.54 | 2.04 | 1.79 | 1.77 | 1.81 | 1.87 | 1.78 |
| 100W (g) | 1.71 | 4.40 | 2.61 | 2.62 | 2.82 | 2.98 | 2.39 |
| Trait a | SL | SW | ST | BL | BW | BT |
|---|---|---|---|---|---|---|
| SW | −0.158 | |||||
| ST | 0.310 | 0.736 | ||||
| BL | 0.967 | −0.218 | 0.274 | |||
| BW | −0.248 | 0.971 | 0.714 | −0.281 | ||
| BT | 0.282 | 0.705 | 0.965 | 0.258 | 0.698 | |
| 100W | 0.650 | 0.531 | 0.830 | 0.631 | 0.491 | 0.807 |
| Marker | Allele | No. | SL | SW | ST | BL | BW | BT | 100W |
|---|---|---|---|---|---|---|---|---|---|
| acc. | (mm) | (mm) | (mm) | (mm) | (mm) | (mm) | (g) | ||
| GW2_SNP | Allele 1 | 46 | 9.72 | 2.72 | 2.05 | 6.95 | 2.28 | 1.82 | 2.82 |
| Allele 2 | 116 | 9.16 | 2.66 | 2.00 | 6.58 | 2.26 | 1.78 | 2.53 | |
| GS3_SNP1 | Allele 1 | 69 | 8.57 | 2.70 | 1.97 | 6.10 | 2.30 | 1.75 | 2.31 |
| Allele 2 | 93 | 9.88 | 2.66 | 2.04 | 7.13 | 2.25 | 1.82 | 2.84 | |
| GS3_SNP2 | Allele 1 | 95 | 9.86 | 2.64 | 2.03 | 7.11 | 2.23 | 1.81 | 2.80 |
| Allele 2 | 67 | 8.56 | 2.74 | 1.98 | 6.09 | 2.33 | 1.77 | 2.35 | |
| GS5_InDel | Allele 1 | 112 | 9.51 | 2.65 | 2.02 | 6.84 | 2.25 | 1.80 | 2.68 |
| Allele 2 | 46 | 8.84 | 2.74 | 1.98 | 6.31 | 2.32 | 1.77 | 2.43 | |
| GW5_InDel | Allele 1 | 42 | 8.94 | 2.96 | 2.09 | 6.33 | 2.48 | 1.87 | 2.75 |
| Allele 2 | 103 | 9.47 | 2.53 | 1.96 | 6.83 | 2.15 | 1.75 | 2.51 | |
| GW6a_InDel2 | Allele 1 | 55 | 9.64 | 2.76 | 2.05 | 6.92 | 2.32 | 1.83 | 2.83 |
| Allele 2 | 107 | 9.16 | 2.64 | 1.99 | 6.57 | 2.25 | 1.78 | 2.50 |
| Marker | SL | SW | ST | BL | BW | BT | 100W |
|---|---|---|---|---|---|---|---|
| GW2_SNP | <0.001 *** | 0.215 | 0.010 * | 0.004 ** | 0.652 | 0.033 * | <0.001 *** |
| GS3_SNP1 | <0.001 *** | 0.472 | <0.001 *** | <0.001 *** | 0.185 | <0.001 *** | <0.001 *** |
| GS3_SNP2 | <0.001 *** | 0.031 * | 0.012 * | <0.001 *** | 0.006 ** | 0.011 * | <0.001 *** |
| GS5_InDel | <0.001 *** | 0.097 | 0.035 * | <0.001 *** | 0.056 | 0.040 * | 0.003 ** |
| GW5_InDel | 0.002 ** | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | 0.003 ** |
| GW6a_InDel2 | 0.001 ** | 0.011 * | 0.001 ** | 0.003 ** | 0.059 | 0.008 ** | <0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S.; Onoda, A.; Orn, C.; Iwamoto, H.; Ishikawa, R.; Saito, H.; Sato, Y.; Ishii, T. Variations in Grain Traits among Local Rice Varieties Collected More Than Half-Century Ago in Indochinese Countries. Plants 2023, 12, 133. https://doi.org/10.3390/plants12010133
Lim S, Onoda A, Orn C, Iwamoto H, Ishikawa R, Saito H, Sato Y, Ishii T. Variations in Grain Traits among Local Rice Varieties Collected More Than Half-Century Ago in Indochinese Countries. Plants. 2023; 12(1):133. https://doi.org/10.3390/plants12010133
Chicago/Turabian StyleLim, Sathya, Anna Onoda, Chhourn Orn, Hiromu Iwamoto, Ryo Ishikawa, Hiroki Saito, Yutaka Sato, and Takashige Ishii. 2023. "Variations in Grain Traits among Local Rice Varieties Collected More Than Half-Century Ago in Indochinese Countries" Plants 12, no. 1: 133. https://doi.org/10.3390/plants12010133